 2021, Vol. 42
2021, Vol. 42

2. 西北大学秦岭研究院, 西安 710127;
3. 武汉大学水资源与水电工程科学国家重点实验室, 武汉 430072
 , XIA Jun1,2,3
, XIA Jun1,2,3 , SONG Jin-xi1,2 , CHANG Jian-bo3 , WU Qiong1,2 , CHENG Dan-dong1,2 , ZHANG Yi-xuan1,2 , KONG Fei-he1,2 , REN Yuan-xin1,2
, SONG Jin-xi1,2 , CHANG Jian-bo3 , WU Qiong1,2 , CHENG Dan-dong1,2 , ZHANG Yi-xuan1,2 , KONG Fei-he1,2 , REN Yuan-xin1,2
2. Institute of Qinling Mountains, Northwest University, Xi'an 710127, China;
3. State Key Laboratory of Water Resources and Hydropower Engineering Science, School of Water Resources and Hydropower Engineering, Wuhan University, Wuhan 430072, China
浮游动物是水生态系统的重要组成部分, 在整个营养系统中转移能量和营养物质, 对水生态系统的物质循环、能量流动、信息传递和稳定性等方面起着重要作用[1].浮游动物对环境变化以及有毒物质极为敏感, 也是衡量水生态健康状况的重要指标[2].物种间的相互作用是维持水生态系统稳定的重要因素, 其中关键物种会对其他物种的丰富度和系统的动态变化产生强烈影响, 并对维持生物多样性及其结构、功能和稳定性起关键作用[3].生态位的概念最初是由动物学家约瑟夫·格林内尔提出的, 他强调了栖息地和食物在定义动物生态位中的作用[4].生态位能够反映种群在时间和空间上所占据的位置、种群分布的交错程度、潜在竞争关系以及对环境资源的利用情况, 是分析和评价群落结构和功能、种间关系、群落动态演替和种群进化等方面的重要手段[5].现阶段, 国内外对于生态位的研究侧重于分析生物优势种生态位分化、群落演替过程、生境适应性以及进化水平上物种的生态位动态研究[6].Dorey等[7]的研究分析了热带小岛蜂的生态位保守性对生物多样性的重要驱动作用.Fu等[8]通过研究植物群落演替过程中物种的生态位动态变化, 揭示了竞争/互利共生与物种生态位之间的关系, 以及演替过程中位移/共存与物种生态位之间的关系.但是, 缺乏通过分子方法对生物多样性及生态位变化特征的研究.
目前, 对于物种的识别主要以观察生物的形态特征为主, 但传统方法在调查过程中会对生态系统产生干扰, 部分物种存在样本采集困难、耗时费力和分辨度低等局限[9].如何快速、准确和有效地揭示生物的多样性仍面临着巨大的挑战[10].近年来, 随着高通量测序技术的快速发展, 生物信息学处理方法和软件的不断更新, 推动了分子技术在生物群落监测方面的研究, 从而使得物种识别方法开始从形态分类向分子生物方法转变[11].环境DNA(environmental DNA, eDNA)是指从环境(如水、土壤或空气)中提取的所有DNA集合, 包括环境微生物和生物体脱落的细胞, 以及在生物死亡后因细胞破裂而释放的细胞外DNA[12]. eDNA技术因通量高、易发现新物种, 在探究生物群落组成和检测物种多样性方面会更加准确、全面和快捷, 适合大规模生物多样性分析[13].该方法能够克服传统方法在调查生物多样性时存在的局限, 表现出更好的检测能力和成本效益[14, 15].
渭河作为黄河流域生态保护战略的重要组成部分, 其水生态安全问题直接关系到黄河流域的高质量发展.但现有研究难以深入而全面地反映该流域的水生态系统的现状[16].因此, 本研究利用eDNA方法分析渭河浮游动物的组成和多样性特征, 探究其空间分布的差异性和群落内部物种间的相互关系.此外, 基于OTU数据从生态位宽度和生态位重叠两个角度揭示关键种(类)的生态位变化特征, 解析环境变化对生物群落的影响机制, 以期为河流生态系统修复及监测管理提供理论指导和科学支撑.
1 材料与方法 1.1 研究区概况渭河(北纬33°42′~37°20′, 东经106°18′~110°37′)是黄河的最大支流, 发源于甘肃省渭源县鸟鼠山(图 1).渭河全长共818 km, 流域内地貌类型复杂多样, 主要流经黄土高原、天水盆地、宝鸡峡谷以及关中平原, 地势西高东低[17].气候类型为大陆性季风气候, 属于暖温带半湿润半干旱的气候带, 年平均气温在7.8~13.5℃之间, 降雨量在350~700 mm之间[18].

|
图 1 渭河采样点示意 Fig. 1 Location of the monitoring stations in the Weihe River |
本研究通过野外实地考察, 并参照国家水文水质监测站的布置, 设置了5个采样断面(图 1).采样工作于2018年4月进行, 采样地点覆盖渭河的上, 中, 下游, 分别为渭源(WY)、陈仓(CC)、眉县(MX)、高陵(GL)和华阴(HY).在每个采样点的表层, 中层和底层采集2 L混合水样, 共收集15个样品, 用于DNA提取.每个DNA样品使用0.45 μm滤膜过滤水样, 然后将富集DNA的滤膜装入无菌试管中, 于-80℃冰箱中保存[19].
1.3 环境因子监测总氮(TN)和总磷(TP)指标根据文献[20]测定.流速、水深、河宽、pH值、溶解氧和水温等指标经现场测量获得.
1.4 eDNA数据分析利用线粒体细胞色素C氧化酶I(COI)基因, 构建测试样品的测序文库.根据QIIME软件中≥97%的相似度, 对获得的高质量序列进行OTU聚类, 并通过BLAST方法将获得的代表序列与数据库进行比较, 获得OTU的分类信息.将COI基因序列与NCBI GenBank参考数据库比对, 进行物种的注释.后续分析是基于数据集中获得的简化OTU列表进行的.在生物多样性分析中, PCA分析(principal component analysis)、冗余分析(RDA)、聚类分析和非度量多维尺度(NMDS)分析均由R软件完成.使用Mothur软件对门水平的各个分类单元在样本之间的序列量差异进行比较检验, 并计算所有物种之间的Spearman等级相关系数, 对其构建关联网络.采用物种的丰度、Chao1丰富度估计指数(SChao1)[21]、ACE丰富度估计指数(SACE)[22]、Shannon多样性指数(H)[23]、Simpson多样性指数(D)[24]、Levins生态位宽度指数(Bi)[25]和Pianka生态位重叠指数(Qik)[26]对浮游动物多样性进行分析.
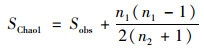
|
(1) |
式中, SChao1表示估计的OTU数, Sobs表示实际观测到的OTU数, n1表示只含有一条序列的OTU数目, n2表示只含有两条序列的OTU数目.
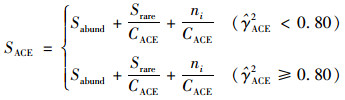
|
(2) |
式中: 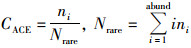
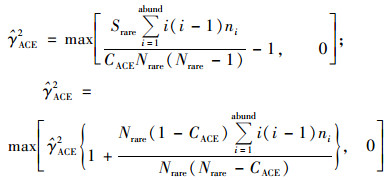
|
式中, ni表示含有i条序列的OTU数目, Srare表示含有“abund”条序列或者少于“abund”的OTU数目, Sabund表示多于“abund”条序列的OTU数目.
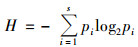
|
(3) |
式中, s为OTU的数量, pi是OTU i在群落中的占比.
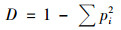
|
(4) |
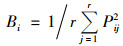
|
(5) |
式中, Bi为生态位宽度, Pij为种i在第j个资源状态下的OTU数占该种所有OTU数比例, r为资源总数, Bi值越大, 生态位宽度就越大.
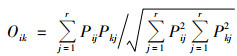
|
(6) |
式中, Oik为生态位重叠指数, Pij和Pkj为种i和种k在第j个资源状态下的OTU数占其所有OTU数比例, r为资源总数.
2 结果与分析 2.1 浮游动物的多样性特征通过对获得的OTU序列进行物种注释发现, 浮游动物主要鉴定为轮虫、枝角类和桡足类, 其中轮虫的OTUs占比最高(图 2).臂尾轮虫属(Brachionus)在下游地区的丰度较高, 而疣毛轮虫属(Synchaeta)则在上游地区占有较大优势.丰度排名前3位的优势物种分别为萼花臂尾轮虫(Brachionus calyciflorus)、异尾轮虫(Trichocerca sp.)和颤动疣毛轮虫(Synchaeta tremula).生物多样性分析表明, Chao1和ACE指数从上游到下游逐渐增大, 其中Chao1指数的变化范围为6.33~39.67, ACE指数的变化范围为6.59~39.67, 两种指数均表明群落的丰富度在逐渐增高.Shannon和Simpson指数也显示下游明显高于上游, 其中Shannon指数在1.73~2.80之间, Simpson指数在0.61~0.82之间, 下游的群落多样性更高(图 3).

|
1.长剑水蚤属(Diacyclops); 2剑水蚤属(Cyclops); 3.龟纹轮属(Anuraeopsis); 4.蒲氏轮虫属(Beauchampia); 5.腔轮属(Lecane); 6.轮虫属(Rotaria); 7.华哲水蚤属(Sinocalanus); 8.扁甲轮属(Plationus); 9须足轮属(Euchlanis); 10.无柄轮虫属(Ascomorpha); 11.纺锤水蚤属(Acartia); 12.哲水蚤属(Bestiolina); 13.晶囊轮虫属(Asplanchna); 14.拟剑水蚤属(Paracyclops); 15.真胖水蚤属(Euchirella); 16.旋轮属(philodina); 17.龟甲轮属(Keratella); 18.多肢轮属(Polyarthra); 19.异尾轮属(Trichocerca); 20.疣毛轮属(Synchaeta); 21.臂尾轮属(Brachionus) 图 2 浮游动物群落组成和丰度分布 Fig. 2 Community composition and abundance distribution of zooplankton |

|
图 3 浮游动物群落的多样性指数 Fig. 3 Diversity index of the zooplankton community |
浮游动物的群落结构呈现出明显的区域差异.轮虫的OTUs丰度表现出从上游到下游逐渐增加的趋势, 而节肢动物(枝角类和桡足类)则呈现出从上游到下游逐渐递减的特征(图 4).NMDS分析结果显示[图 5(a)], 上下游群落间的空间分布存在明显的差异性(P < 0.05).层次聚类结果显示, 下游的HY和GL聚为一类, 而上中游的WY和MX、CC各自聚为一类[图 5(b)].说明上下游不同河段的浮游动物群落在物种组成、相对丰度及多样性方面存在空间异质性.此外, 也反映出浮游动物存在距离衰减效应, 群落结构随着地理距离、海拔距离和环境异质性的增大, 相似性逐渐降低.

|
图 4 浮游动物丰度的空间差异 Fig. 4 Spatial differences in the abundance of zooplankton |

|
图 5 基于浮游动物OTUs的NMDS分析和样本层次聚类分析 Fig. 5 Cluster analysis of the zooplankton community structure |
分子生态网络分析显示, 群落内物种间的网络节点数和相互连接度较高, 网络结构表现出复杂化特点(图 6).群落中丰度高的优势种, 如臂尾轮虫(Brachionus)、多肢轮虫(Polyarthra sp.)以及异尾轮虫(Trichocereca sp.)之间连接度较高, 具有高节点度、高紧密中心性, 且呈现模块化特征.同时优势种与群落内其它物种间存在着直接或间接的联系, 在群落中起着至关重要的作用.此外, 剑水蚤(Cyclops sp.)、扁甲轮虫(Plationus patulus)、龟甲轮虫(Keratella sp.)和细巧华哲水蚤(Sinocalanus_tenellus)等稀有物种(相对丰度低于1%)在群落中也有较高的连接度.这些高连接度的优势种和稀有物种共同组成群落的关键种.

|
图 6 浮游动物群落的网络结构 Fig. 6 Network structure of the zooplankton community |
生态位宽度一般分为广生态位(Bi>0.60)、中生态位(Bi为0.30~0.60)和窄生态位(Bi < 0.30)这3种类型.综合浮游动物OTUs的丰度和连接度, 对关键种进行生态位分析.生态位宽度值(Bi)计算结果显示(表 1), 渭河浮游动物关键种的生态位宽度值(Bi)变化范围为0.38~0.80, 主要为广生态位和中生态位种(类)群.广生态位的物种有疣毛轮虫(0.80)、颤动疣毛轮虫(0.80)和萼花臂尾轮虫(0.61).中生态位物种包括龟甲轮虫(0.56)、异尾轮虫(0.40)、剑水蚤(0.40)、扁甲轮虫(0.40)和多肢轮虫(0.38), 中生态位种类占全部关键种(类)的63%.渭河浮游动物关键种之间的生态位重叠程度总体较高, 有23对关键种之间的Qik大于0.60, 平均值为0.72, 高度重叠者占比高达72%, 且部分物种间生态位重叠指数达到了0.90以上.
|
|
表 1 浮游动物关键种(类)生态位宽度值和生态位重叠指数1) Table 1 Niche breadth and the niche overlap index of the keystone species |
2.5 浮游动物群落与环境因子的关系
浮游动物群落关键种OTUs数据与环境因子间的冗余分析结果显示(图 7), 浮游动物群落结构与环境因素有明显的关系, 且存在空间异质性特征.RDA排序图前两轴累计解释率为58.81%(P=0.001), 总氮、水温、总磷、河宽、pH值和水深等环境因素之间呈正相关, 且对下游的关键物种的影响较大, 其中总氮(R2=0.896 7, P=0.001)和水温(R2=0.888 8, P=0.001)是其主要限制因子.溶解氧和流速对上游河段浮游动物群落影响更大.以上结果表明, 环境因子与浮游动物关键种存在显著的相关性(P < 0.01), 冗余分析能够反映浮游动物种群及生态位变化与环境因子间的基本关系.

|
图 7 浮游动物群落关键种与环境因子的RDA排序 Fig. 7 Redundancy analysis of the zooplankton community and significant environmental variables |
渭河浮游动物群落的种类组成以轮虫类为主, 个体较大的枝角类和桡足类丰度相对较低, 与该地区已有的传统方法调查结果一致[27].在物种数量鉴定方面, eDNA技术通过较少的断面监测即获得了较多的物种信息, 显示出分子方法在物种鉴定方面具有较高的物种分辨率和检测效率[28].不同区域间物种组成的差异可能是受到浮游动物的繁殖、发育特征以及环境适用性等因素的影响[29].水体理化环境和水文条件的变化, 对生物群落的多样性和空间分布起着重要作用.上游河段流速快、温度低, 不利于浮游生物的生长发育, 导致上游生物多样性指数偏低[30].下游河段有泾河和北洛河两大支流汇入, 营养物质丰富, 且该区域海拔较低, 水体流速较缓, 水温较高, 适宜浮游动物的生长繁殖[31].此外, 浮游动物扩散能力弱, 存在一定的距离衰减模式.中性理论认为, 物种的迁移、扩散等行为在塑造生物群落方面发挥着重要作用[32].
3.2 浮游动物群落的分子生态网络分析网络分析揭示了群落内轮虫与枝角类和桡足类之间存在的复杂关联.有研究表明, 节肢动物(枝角类和桡足类)和轮虫类之间可能存在资源利用性竞争, 机械干涉性竞争和化学干涉性竞争[33].群落中优势类群间联系密切且存在模块化现象, 说明优势种在群落中占据相似的生态位, 并具有类似的生态功能[34].群落中稀有类群与优势类群共存, 共同在群落功能和稳定性方面发挥着重要驱动作用.此外, 关键种在维持食物网稳定性方面也发挥着不可替代的作用, 例如, 通过密切联系群落的其他物种, 维持捕食者的密度来限制其他被捕食者的密度, 控制群落的结构和能量流动[35].臂尾轮虫等关键种(类)会通过影响其他物种的丰度或使生态位发生改变的方式对生态系统的动态产生强烈影响[36].此外, 食物密度、环境耐受性、移动速度以及温度等水环境变化也会对种群关系产生影响[37].
3.3 浮游动物关键种的生态位分化及环境适应性生态位是生物种(类)对环境适应的结果, 能够反映其应对环境变化的策略, 进而决定物种的分布格局[38].广生态位物种对于环境的适应能力和资源的利用能力最强, 其次为中生态位的生物类群, 而窄生态位的物种适应性最低, 对生境的变化最为敏感[39].渭河浮游动物关键种(类)主要为广生态位类型, 且生态位重叠指数较高.生态位重叠指数能够充分反映浮游动物关键物种(类)间对资源利用的相互关系[40].已有研究表明, 当生态位重叠指数>0.6时, 群落内物种在同一生境内对空间和食物资源利用的相似性也较高, 并对水环境的变化表现出较好的生态适应性[41].生态位重叠度较高的关键种群, 在资源足够丰富的情况下不会引发竞争, 但当资源匮乏时, 会导致竞争加剧.白洋淀浮游动物的调查结果也显示, 优势种间的生态位重叠平均值为0.679, 高度重叠者的占比高达61.90%[42].此外, 重叠度较高的关键种在竞争中会通过生态位分化降低物种间的竞争, 从而促进共存和维持整个系统的生物多样性[43].但是, 部分生态位宽度较大的物种, 其生态学特征和对资源的利用也并非完全一致, 生态位宽度较大的物种间也可能会出现较低幅度的生态位重叠[44].
物种的群落结构和生态位均与环境变化密切相关, 物种对资源的利用及对环境的适应也会受到环境变化的影响[45].渭河水体中的总氮含量高, 已超过Ⅴ类水标准, 说明受到有机物的严重污染.冗余分析结果显示, 导致渭河生物多样性产生变动的最主要水质因素是氮的含量.相关研究表明, 浮游动物群落与水体营养物质的变化密切相关, 营养物质不仅能通过影响浮游植物生长来影响浮游动物, 还可通过渗透和离子调节机制影响浮游动物生长繁殖[46].渭河的河道形态复杂多样, 海拔落差大, 加上支流汇入带来的营养物质, 导致生活在该区域的浮游动物组成和分布呈现多样化[47].渭河宝鸡峡大坝改变了原有河流的水文、水动力学条件, 也改变了上下游间的栖息地环境, 继而对浮游动物的基因、物种和分布模式产生影响[48].下游河段出现的浮游动物以轮虫类为主, 其中臂尾轮虫、异尾轮虫、多肢轮虫和龟甲轮虫等富营养化指示种与关中城市群区域所产生的污水密切相关[49].此外, 浮游动物群落的构建也会受到随机生态漂变和生态位分化共同的作用[50].总体而言, 浮游动物多样性和生态位的异质性反映了群落和环境间的响应关系.
4 结论(1) 浮游动物多样性呈现出从上游到下游逐渐增高的特征.群落结构在各监测断面间具有显著的区域差异(P < 0.05), 并存在距离衰减效应.
(2) 浮游动物分子生态网络具有高节点度、高紧密中心性, 且呈现模块化特征.优势种和稀有物种共同在群落中发挥重要作用.
(3) 渭河浮游动物关键种的生态位宽度值变化范围在0.38~0.80之间, 主要为广生态位和中生态位种(类)群.生态位重叠程度总体较高, 其中高度重叠者占比高达72%, 且部分物种间生态位重叠指数达到了0.90以上.
(4) 浮游动物群落结构主要与水体物理特征(水温、河宽、流速和水深)以及氮磷元素密切相关(P < 0.01).总氮(R2=0.8967, P=0.001)和水温(R2=0.8888, P=0.001)是其主要限制因子, 对浮游动物的群落结构的变化具有重要影响.
| [1] | Ismail A H, Lim C C, Omar W M W. Evaluation of spatial and temporal variations in zooplankton community structure with reference to water quality in Teluk Bahang Reservoir, Malaysia[J]. Tropical Ecology, 2019, 60(2): 186-198. DOI:10.1007/s42965-019-00023-2 |
| [2] | Isibor P O, Imoobe T O T, Dedeke G A, et al. Health risk indices and zooplankton-based assessment of a tropical rainforest river contaminated with iron, lead, cadmium, and chromium[J]. Scientific Reports, 2020, 10(1). DOI:10.1038/s41598-020-72526-1 |
| [3] | Krztoń W, Kosiba J, Pociecha A, et al. The effect of cyanobacterial blooms on bio-and functional diversity of zooplankton communities[J]. Biodiversity and Conservation, 2019, 28(7): 1815-1835. DOI:10.1007/s10531-019-01758-z |
| [4] | Grinnell J. Field tests of theories concerning distributional control[J]. The American Naturalist, 1917, 51(602): 115-128. DOI:10.1086/279591 |
| [5] | Barton P S, Westgate M J, Foster C N, et al. Using ecological niche theory to avoid uninformative biodiversity surrogates[J]. Ecological Indicators, 2020, 108. DOI:10.1016/j.ecolind.2019.105692 |
| [6] | Feng K, Wang S, Wei Z Y, et al. Niche width of above-and below-ground organisms varied in predicting biodiversity profiling along a latitudinal gradient[J]. Molecular Ecology, 2020, 29(10): 1890-1902. DOI:10.1111/mec.15441 |
| [7] | Dorey J B, Groom S V C, Freedman E H, et al. Radiation of tropical island bees and the role of phylogenetic niche conservatism as an important driver of biodiversity[J]. Proceedings of the Royal Society B: Biological Sciences, 2020, 287(1925). DOI:10.1098/rspb.2020.0045 |
| [8] | Fu W G, Wang F K, Yin Q L, et al. Niche dynamics of species in succession process in the wetland of Yangtze Rivers Lower Reach, China[J]. Plant Ecology and Evolution, 2015, 148(1): 43-51. DOI:10.5091/plecevo.2015.956 |
| [9] | Yang J H, Zhang X W. eDNA metabarcoding in zooplankton improves the ecological status assessment of aquatic ecosystems[J]. Environment International, 2020, 134. DOI:10.1016/j.envint.2019.105230 |
| [10] | Furlan E M, Gleeson D, Wisniewski C, et al. eDNA surveys to detect species at very low densities: a case study of European carp eradication in Tasmania, Australia[J]. Journal of Applied Ecology, 2019, 56(11): 2505-2517. DOI:10.1111/1365-2664.13485 |
| [11] |
张莉, 张远, 林佳宁, 等. 基于多个扩增子的DNA metabarcoding技术探究黄海微型真核浮游植物多样性[J]. 环境科学, 2019, 40(9): 4052-4060. Zhang L, Zhang Y, Lin J N, et al. Community diversity of eukaryotic nano-phytoplankton in Yellow Sea using DNA metabarcoding technology based on multiple amplicons[J]. Environmental Science, 2019, 40(9): 4052-4060. |
| [12] | Taberlet P, Coissac E, Hajibabaei M, et al. Environmental DNA[J]. Molecular Ecology, 2012, 21(8): 1789-1793. DOI:10.1111/j.1365-294X.2012.05542.x |
| [13] | Valentini A, Taberlet P, Miaud C, et al. Next-generation monitoring of aquatic biodiversity using environmental DNA metabarcoding[J]. Molecular Ecology, 2016, 25(4): 929-942. DOI:10.1111/mec.13428 |
| [14] |
徐念, 常剑波. 长江中下游干流环境DNA样本鱼类物种检测的初步研究[J]. 水生态学杂志, 2016, 37(5): 49-55. Xu N, Chang J B. Preliminary study on fish species detection in the middle and lower Yangtze River using environmental DNA[J]. Journal of Hydroecology, 2016, 37(5): 49-55. |
| [15] | Bakker J, Wangensteen O S, Baillie C, et al. Biodiversity assessment of tropical shelf eukaryotic communities via pelagic eDNA metabarcoding[J]. Ecology and Evolution, 2019, 9(24): 14341-14355. DOI:10.1002/ece3.5871 |
| [16] |
贺玉晓, 李珂, 任玉芬, 等. 春季北京市河流大型底栖动物群落结构特征及影响因子分析[J]. 环境科学, 2020, 41(6): 2951-2962. He Y X, Li K, Ren Y F, et al. Characteristics of macrobenthos community structure and their relationships with environmental factors in rivers of Beijing in spring[J]. Environmental Science, 2020, 41(6): 2951-2962. |
| [17] |
张妍, 毕直磊, 张鑫, 等. 土地利用类型对渭河流域关中段地表水硝酸盐污染的影响[J]. 生态学报, 2019, 39(12): 4319-4327. Zhang Y, Bi Z L, Zhang X, et al. Effects of land-use types on nitrate pollution of surface water in Guanzhong area in the Weihe River basin[J]. Acta Ecologica Sinica, 2019, 39(12): 4319-4327. |
| [18] | Song J X, Yang X G, Zhang J L, et al. Assessing the variability of heavy metal concentrations in liquid-solid two-phase and related environmental risks in the Weihe River of Shaanxi Province, China[J]. International Journal of Environmental Research and Public Health, 2015, 12(7): 8243-8262. DOI:10.3390/ijerph120708243 |
| [19] |
陈治, 宋娜, 源利文, 等. 舟山近海水样环境DNA获取方法的建立[J]. 水生生物学报, 2020, 44(1): 50-58. Chen Z, Song N, Minamoto T, et al. The eDNA collection method of Zhoushan coastal waters[J]. Acta Hydrobiologica Sinica, 2020, 44(1): 50-58. |
| [20] | 国家环境保护总局. 水和废水监测分析方法[M]. (第四版). 北京: 中国环境科学出版社, 2002. |
| [21] | Chao A. Nonparametric estimation of the number of classes in a population[J]. Scandinavian Journal of Statistics, 1984, 11(4): 265-270. |
| [22] | Chao A N, Ma M C, Yang M C K. Stopping rules and estimation for recapture debugging with unequal failure rates[J]. Biometrika, 1993, 80(1): 193-201. DOI:10.1093/biomet/80.1.193 |
| [23] | Shannon C E. A mathematical theory of communication[J]. The Bell System Technical Journal, 1948, 27(3): 379-423. DOI:10.1002/j.1538-7305.1948.tb01338.x |
| [24] | Simpson E H. Measurement of diversity[J]. Nature, 1949, 163(4148): 688. DOI:10.1038/163688a0 |
| [25] | Levins R. Evolution in changing environments: some theoretical explorations[M]. Princeton: Princeton University Press, 1968. |
| [26] | Pianka E R. The structure of lizard communities[J]. Annual Review of Ecology and Systematics, 1973, 4: 53-74. DOI:10.1146/annurev.es.04.110173.000413 |
| [27] |
王硕, 杨涛, 李小平, 等. 渭河流域浮游动物群落结构及其水质评价[J]. 水生生物学报, 2019, 43(6): 1333-1345. Wang S, Yang T, Li X P, et al. Zooplankton community structure and the water quality in the Wei River Basin[J]. Acta Hydrobiologica Sinica, 2019, 43(6): 1333-1345. |
| [28] |
王靖淇, 王书平, 张远, 等. 高通量测序技术研究辽河真核浮游藻类的群落结构特征[J]. 环境科学, 2017, 38(4): 1403-1413. Wang J Q, Wang S P, Zhang Y, et al. Community structure characteristics of eukaryotic planktonic algae in Liaohe River through high-throughput sequencing[J]. Environmental Science, 2017, 38(4): 1403-1413. |
| [29] | Gabaldón C, Devetter M, Hejzlar J, et al. Seasonal strengths of the abiotic and biotic drivers of a zooplankton community[J]. Freshwater Biology, 2019, 64(7): 1326-1341. DOI:10.1111/fwb.13308 |
| [30] | Zhou J, Qin B Q, Han X X. The synergetic effects of turbulence and turbidity on the zooplankton community structure in large, shallow Lake Taihu[J]. Environmental Science and Pollution Research, 2018, 25(2): 1168-1175. DOI:10.1007/s11356-017-0262-1 |
| [31] | Portinho J L, Perbiche-Neves G, Nogueira M G. Zooplankton community and tributary effects in free-flowing section downstream a large tropical reservoir[J]. International Review of Hydrobiology, 2016, 101(1-2): 48-56. DOI:10.1002/iroh.201501798 |
| [32] | Chen W D, Ren K X, Isabwe A, et al. Stochastic processes shape microeukaryotic community assembly in a subtropical river across wet and dry seasons[J]. Microbiome, 2019, 7(1). DOI:10.1186/s40168-019-0749-8 |
| [33] | Carter J L, Schindler D E, Francis T B. Effects of climate change on zooplankton community interactions in an Alaskan lake[J]. Climate Change Responses, 2017, 4(1). DOI:10.1186/s40665-017-0031-x |
| [34] | Dupont Y L, Olesen J M. Ecological modules and roles of species in heathland plant-insect flower visitor networks[J]. Journal of Animal Ecology, 2009, 78(2): 346-453. DOI:10.1111/j.1365-2656.2008.01501.x |
| [35] | Jordán F. Keystone species and food webs[J]. Philosophical Transactions of the Royal Society B: Biological Sciences, 2009, 364(1524): 1733-1741. DOI:10.1098/rstb.2008.0335 |
| [36] | Yang S, Pan T, Wang X, et al. Dominant aquatic species and their hydrological niches in freshwater ecosystems in a developing city[J]. Marine and Freshwater Research, 2019, 70(5): 615-624. DOI:10.1071/MF18336 |
| [37] | Voutilainen A, Jurvelius J, Lilja J, et al. Associating spatial patterns of zooplankton abundance with water temperature, depth, planktivorous fish and chlorophyll[J]. Boreal Environment Research, 2016, 21(1-2): 101-114. |
| [38] |
侯朝伟, 孙西艳, 刘永亮, 等. 烟台近海浮游动物优势种空间生态位研究[J]. 生态学报, 2020, 40(16): 5822-5833. Hou C W, Sun X Y, Liu Y L, et al. Spatial niches of dominant zooplankton species in the Yantai offshore waters[J]. Acta Ecologica Sinica, 2020, 40(16): 5822-5833. |
| [39] | Richmond J Q, Ota H, Grismer L L, et al. Influence of niche breadth and position on the historical biogeography of seafaring scincid lizards[J]. Biological Journal of the Linnean Society, 2021, 132(1): 74-92. DOI:10.1093/biolinnean/blaa172 |
| [40] | Huang S Q, Zhou Q, Gu L C, et al. Modeling and simulation study of the stoichiometric niche space and niche overlap based on the copula method[J]. Environmental and Ecological Statistics, 2020, 27(1): 115-149. DOI:10.1007/s10651-019-00437-0 |
| [41] |
郑挺, 林元烧, 曹文清, 等. 北部湾北部生态系统结构与功能——浮游动物空间生态位及其分化[J]. 生态学报, 2014, 34(13): 3635-3649. Zheng T, Lin Y S, Cao W Q, et al. Ecosystem structure and function in northern Beibu Gulf: zooplankton spatial niche and its differentiation[J]. Acta Ecologica Sinica, 2014, 34(13): 3635-3649. |
| [42] |
付显婷, 杨薇, 赵彦伟, 等. 白洋淀浮游动物群落结构与水环境因子的关系[J]. 农业环境科学学报, 2020, 39(6): 1271-1282. Fu X T, Yang W, Zhao Y W, et al. Community structure of zooplankton and its relationship with water environmental factors in Baiyangdian Lake[J]. Journal of Agro-Environment Science, 2020, 39(6): 1271-1282. |
| [43] | Chaloner T M, Gurr S J, Bebber D P. Geometry and evolution of the ecological niche in plant-associated microbes[J]. Nature Communications, 2020, 11(1). DOI:10.1038/s41467-020-16778-5 |
| [44] | Manlick P J, Pauli J N. Human disturbance increases trophic niche overlap in terrestrial carnivore communities[J]. Proceedings of the National Academy of Sciences of the United States of America, 2020, 117(43): 26842-26848. DOI:10.1073/pnas.2012774117 |
| [45] |
杨佳, 周健, 秦伯强, 等. 太湖梅梁湾浮游动物群落结构长期变化特征(1997~2017年)[J]. 环境科学, 2020, 41(3): 1246-1255. Yang J, Zhou J, Qin B Q, et al. Long-term variation characteristics of zooplankton community structure in Meiliang Bay, Lake Taihu[J]. Environmental Science, 2020, 41(3): 1246-1255. DOI:10.3969/j.issn.1000-6923.2020.03.036 |
| [46] | Ișkιn U, Filiz N, Cao Y, et al. Impact of nutrients, temperatures, and a heat wave on zooplankton community structure: an experimental approach[J]. Water, 2020, 12. DOI:10.3390/w12123416 |
| [47] |
孙胜浩, 陈娟, 王沛芳, 等. 澜沧江硅藻的地理分布模式与关键驱动因素[J]. 环境科学, 2020, 41(12): 5458-5469. Sun S H, Chen J, Wang P F, et al. Biogeographic distribution patterns of diatoms in Lancang River and their key Drivers[J]. Environmental Science, 2020, 41(12): 5458-5469. |
| [48] | de Souza C A, Vieira L C G, Legendre P, et al. Damming interacts with the flood pulse to alter zooplankton communities in an Amazonian river[J]. Freshwater Biology, 2019, 64(5): 1040-1053. DOI:10.1111/fwb.13284 |
| [49] |
杜彩丽, 杨丽, 赵诣, 等. 淀山湖浮游动物群落时空分布特征及其与环境因子的关系[J]. 环境科学, 2019, 40(10): 4513-4522. Du C L, Yang L, Zhao Y, et al. Temporal and spatial variation of zooplankton community structure and its relationship with environmental factors in Dianshan Lake, Shanghai[J]. Environmental Science, 2019, 40(10): 4513-4522. |
| [50] | Siqueira T, Saito V S, Bini L M, et al. Community size can affect the signals of ecological drift and niche selection on biodiversity[J]. Ecology, 2020, 101(6). DOI:10.1002/ecy.3014 |