 2021, Vol. 42
2021, Vol. 42



2. 西北大学生命科学学院, 西安 710127;
3. 西北农林科技大学农学院, 杨凌 712100
2. College of Life Sciences, Northwest University, Xi'an 710127, China;
3. College of Agronomy, Northwest A&F University, Yangling 712100, China
森林土壤碳库是陆地生态系统中最大的碳库, 约占全球土壤碳库的40%, 因此即使森林SOC库发生微小的变化, 都会对大气CO2含量和全球碳平衡产生深远影响[1, 2].土壤团聚体作为土壤重要的组成部分和土壤结构的基本单位, 可以在SOC和土壤微生物之间形成空间隔离[3], 从而减少微生物对SOC的分解利用并降低土壤CO2的排放.此外, 土壤团聚体还具有维持土壤孔隙、减少侵蚀、防止水土流失[4]和影响微生物群落结构[5]等作用, 其稳定性是衡量土壤质量以及土壤结构和土壤可持续利用的重要指标[6].因此探究森林土壤团聚体的稳定性对降低森林土壤的CO2排放, 增加其固碳能力具有重要意义.
在森林生态系统中, 土壤团聚体稳定性很大程度上受海拔梯度、植被类型和微生物等因素影响[3].海拔梯度作为一种自然地理变化, 对植被带谱分布、土壤理化性质以及微生物组成等都有影响[7, 8]. Zhu等[9]在四川雪宝顶山研究发现, 高海拔呈现出较高的土壤团聚体稳定性; 而Murugan等[10]和吴梦瑶等[7]分别在阿尔卑斯山和贺兰山的研究中发现, 中海拔具有较高的土壤团聚体稳定性, 其稳定性具有随着海拔呈先增高后降低的趋势.因此, 学者们对于土壤团聚体稳定性在不同海拔之间的研究还存在有结果的不一致性.并且, 关于土壤团聚体稳定性随海拔变化的调控因子尚未达成共识.因为其不仅受土壤理化性质(如土壤温度、湿度)[11]和植被群落[12]的影响, 还受多种微生物活动的影响, 如微生物生物量, 胞外酶活性和微生物养分利用率[13, 14].国内外学者就土壤有机质对团聚体稳定性的影响已做了大量的研究工作, 王兴等[15]和Deng等[16]的研究结果均表明土壤团聚体稳定性与有机质水平的高低密切相关, 而多项研究表明生物变量(如微生物量和胞外酶)在土壤团聚体的形成和稳定性中起重要作用[14, 17].因此, 在海拔梯度上开展团聚体稳定性及影响因素的研究显得重要而迫切, 这对于研究森林土壤有机碳的储存和提高土壤可持续发展具有重要意义.
秦岭林区作为我国重要的生态屏障, 是我国南北气候的天然分界线和长江、黄河的分水岭, 也是我国暖温带森林系统的典型代表.尤其秦岭主峰太白山林区不仅垂直植被带谱分布完整、跨度大, 且生物多样性丰富、植被固碳功能潜力巨大, 在中国森林碳汇中占有重要地位, 为温室气体减排提供重要保障, 也为人们开展森林土壤团聚体稳定性机制的研究提供天然的试验平台[18].鉴于此, 本文选择太白山不同海拔梯度的锐齿栎林、辽东栎林和红桦林(1 503~2 451 m)这3种典型植被下的土壤为研究对象, 阐明土壤团聚体稳定性在海拔上的变化规律及其影响因子, 探究不同粒级的团聚体的胞外酶对土壤团聚体稳定性的潜在影响.
1 材料与方法 1.1 研究区概况秦岭是我国南北气候的天然分界线, 是我国重要的生态屏障.其主峰太白山位于秦岭山脉中部(33°45′~34°10′N, 107°19′~107°58′E), 海拔3 767.2 m.太白山属温带季风气候, 年平均气温11.4℃, 年平均降水量910.6 mm, 降水主要集中在7~9月.太白山植被垂直带谱分布完整且跨度大, 植被类型丰富, 不同典型垂直植被林带以锐齿栎林(1 253~1 856 m)、辽东栎林(1 900~2 265 m)、桦木林(2 356~2 500 m)、冷杉林(2 800~3 000 m)和落叶松(3 000~3 400 m)为主.样地基本信息见表 1.
|
|
表 1 样地基本信息 Table 1 Basic information of the plot |
1.2 土壤样品采集
2019年7月, 在太白山锐齿栎(1 503 m)、辽东栎(1 915 m)和红桦(2 405 m)这3个植被带上选取地形、地貌、坡度和坡向等基本一致的标准样地(每个植被带3个重复样地), 每个样地内设3个样方(20 m×20 m), 共设9个样地27个样方.去除地表凋落物和腐殖层后, 采用S型10点采样法采集各样方表层(0~10 cm)土壤, 将同一样方内10个土壤样品均匀混合后装入土壤袋中, 立即放置在便携式冷藏箱内.将上述样品带回实验室后, 剔除石块、根系和动植物残体等杂物, 碾压过2 mm筛后分成两部分, 一部分土壤鲜样置于4℃冰箱保存, 一周内进行土壤酶活性和微生物量测定.其余土壤自然风干, 测定其他土壤理化性质.
同时, 采集各样方内表层(0~10 cm)原状土壤, 装入铝盒带回实验室, 按照土壤自然纹理轻柔掰开, 剔除石块、根系和动植物残体等, 全部过8 mm筛并将同一样地样品混合.采用干筛法将土壤筛分为大团聚体(>2 mm), 中团聚体(0.25~2 mm)与微团聚体(< 0.25 mm)3个等级.将筛分后的各团聚体土壤一部分放于4℃冰箱保存, 一周内进行土壤酶活性和微生物量测定; 其余部分自然风干, 测定其他土壤理化性质.
1.3 植物信息采集在每个植被带调查样方(20 m×20 m)内观测记录包括乔木层、灌木层和草本层植物.记录样方内出现的全部乔木种, 测量所有DBH≥3 cm植株高度和胸径; 于乔木层样方设置2个10 m×10 m的对角小样方进行灌木层调查, 记录样方内出现的全部灌木种, 对其中的全部灌木分别计数, 并测量基径和高度; 于乔木层样方的中心和四角位设置5个1 m×1 m的草本层小样方, 记录样方内出现的所有草本种类, 记录每种草本植物的多度、频度、高度和盖度.
1.4 样品测定土壤有机碳(SOC)的测定采用重铬酸钾-浓硫酸外加热法[19], 土壤全氮(TN)和全磷(TP)分别经H2SO4和H2SO4-HClO4消解后, 利用AA3型连续流动分析仪测定(Bran+Luebbe AAIII, 德国)[20], 土壤微生物量(MBC、MBN、MBP)采用氯仿熏蒸-硫酸钾浸提法[21, 22], 参照Saiya-Cork等[23]描述的96微孔板荧光法测量土壤中β-葡萄糖苷酶(β-glucosidase, EC 3.2.1.21, BG)、β-N-乙酰氨基葡萄糖酶(β-N-Acetyl-glucosaminidase, EC 3.2.1.52, NAG)、亮氨酸氨基肽酶(Leucine arylamidase, LAP)和酸性磷酸酶(Acid phosphatase, EC 3.1.3.2, ACP)的活性, 利用多功能酶标仪(Tecan Infinite M200, 奥地利)检测荧光值.
1.5 数据分析采用物种丰富度(species richness)、Shannon-Wiener多样性指数和Simpson多样性指数对不同海拔的植被多样性进行分析, 计算公式如下.
物种丰富度(S): 出现在样方内的物种数
Shannon-Wiener多样性指数(H):
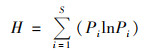
|
Simpson多样性指数(P):
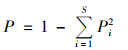
|
式中, Pi为第i个物种的个体数占所有物种个体数的百分比.
土壤微生物群落和土壤资源(SOC、TN、TP)间的关系是通过土壤C: N(C: P)与微生物量C: N(C: P)的比率计算的.为评估不同海拔微生物C: N, C: P稳态(H′), 使用以下方程式[24]:
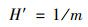
|
式中, m为土壤资源ln(SOC: TN)与微生物ln(MBC: MBN)的斜率或土壤资源ln(SOC: TP)与微生物ln(MBC: MBP)的斜率.当H′≫1或拟合结果不显著(P>0.05)时, 土壤微生物群落呈现强稳态, 而当H′≈1时, 呈现弱稳态.
为了探索不同海拔下的微生物如何调节元素利用效率, 使用了以下方程式计算元素阈值(Ter C: N和Ter C: P), 将测定的微生物量和胞外酶活性联系起来, 阐明土壤当前的养分限制[25]:
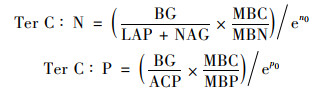
|
式中, BG为β-葡萄糖苷酶的酶活性, LAP为亮氨酸氨基肽酶的酶活性, NAG为β-N-乙酰氨基葡萄糖酶的酶活性, ACP为酸性磷酸酶的酶活性, MBC、MBN和MBP分别表示微生物量碳、微生物量氮和微生物量磷含量.n0表示lnBG对ln (LAP+NAG)的标准主轴回归分析(standardized major axis, SMA)所得到方程的截距, p0表示lnBG对lnACP的标准主轴回归分析所得到的方程截距.标准主轴回归分析在Rstudio中完成.
土壤团聚体稳定性指标选取平均重量直径(mean weight diameter, MWD), 几何平均直径(geometric mean diameter, GMD)[26].
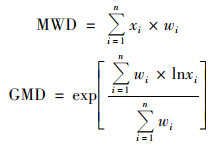
|
式中, xi为第i级团聚体平均直径(mm), wi为第i级团聚体所占质量分数.
偏最小二乘法路径模型(partial least squares path models, PLS-PM)是一种研究观测变量的数据分析方法, 该方法对样本数量要求低且具备高水平统计能力[27], 因此应用PLS-PM探究土壤团聚体稳定性指标MWD和GMD与土壤理化性质、微生物生物量、胞外酶活性及其化学计量比、元素阈值间的关系.
利用单因素方差分析(one-way ANOVA)检验不同海拔混合土壤和团聚体酶活性、酶活性计量比、团聚体稳定性和元素阈值的差异, 利用逐步回归探究团聚体酶活性对团聚体稳定性的驱动因子, 所有分析处理在SPSS 19.0中进行, 用Origin 8.5软件作图.
2 结果与分析 2.1 秦岭海拔梯度上土壤理化性质及植被群落特征土壤有机碳、全氮和全磷含量在海拔梯度上变化特征基本一致(表 2), 均随着海拔升高而显著增加(P < 0.05).土壤C: N、C: P、N: P、容重和pH均表现为海拔梯度Ⅰ显著大于Ⅲ(P < 0.05), 相反, 土壤含水率表现为海拔梯度Ⅲ显著大于海拔梯度Ⅰ(P < 0.05).不同海拔植被群落特征见表 3.
|
|
表 2 不同海拔的土壤理化性质1) Table 2 Physical and chemical properties of soil at different altitudes |
|
|
表 3 不同海拔的植被群落特征 Table 3 Characteristics of vegetation communities at different altitudes |
2.2 秦岭海拔梯度土壤酶活性及计量比值变化
BG、NAG和ACP酶活性在海拔梯度上的变化趋势基本一致, 均为海拔梯度Ⅰ显著大于Ⅱ和Ⅲ, 其中BG酶活性Ⅰ分别显著大于Ⅱ 116.14%(P < 0.05, 95%CI:84.32%~147.96%)和Ⅲ 101.16%(P < 0.05, 95%CI:71.55%~130.78%); NAG酶活性Ⅰ分别显著大于Ⅱ 249.11%(P < 0.05, 95%CI:211.23%~286.98%)和Ⅲ 319.93%(P < 0.05, 95%CI:274.37%~365.49%); ACP酶活性Ⅰ分别显著大于Ⅱ 260.06%(P < 0.05, 95%CI:190.11%~330.01%)和Ⅲ 206.89%(P < 0.05, 95%CI:147.27%~266.51%).而LAP酶活性表现为Ⅲ显著大于Ⅰ 35.49%(P < 0.05, 95%CI:13.54%~57.44%), 在Ⅰ和Ⅱ(P>0.05, 95%CI:4.93%~31.53%)或Ⅱ和Ⅲ(P>0.05, 95%CI:1.91%~26.03%)之间差异性不显著(图 1).土壤酶活性计量比在不同海拔间都具有显著差异(P < 0.05).总体上来说, 海拔梯度Ⅲ具有相对较高BG: (NAG+LAP)值、BG: ACP值和相对较低的(NAG+LAP): ACP值; 而海拔梯度Ⅰ具有相对较低的BG: (NAG+LAP)值、BG: ACP值和相对较高的(NAG+LAP): ACP值(图 2).

|
海拔梯度Ⅰ为锐齿栎林, 1 503 m, 海拔梯度Ⅱ为辽东栎林, 1 915 m, 海拔梯度Ⅲ为红桦林, 2 405 m; 不同小写字母代表海拔间差异显著(P < 0.05), 下同; BG表示β-1, 4-葡萄糖苷酶, NAG表示β-1, 4-N-乙酰葡糖氨糖苷酶, LAP表示亮氨酸氨基肽酶, ACP表示酸性磷酸酶 图 1 不同海拔土壤酶活性 Fig. 1 Soil enzyme activities at different altitudes |

|
BG: (NAG+LAP)表示碳氮比, BG: ACP表示碳磷比, (NAG+LAP): ACP表示氮磷比 图 2 不同海拔土壤酶活性比差异 Fig. 2 Variation of soil enzymatic ratios at different altitudes |
土壤团聚体稳定性指标GMD在海拔梯度Ⅰ、Ⅱ和Ⅲ上分别为1.66 mm、1.39 mm和1.32 mm; MWD在海拔梯度Ⅰ、Ⅱ和Ⅲ上分别为2.17 mm、1.83 mm和1.82 mm; 二者均表现为海拔梯度Ⅰ显著大于Ⅱ和Ⅲ(P < 0.05, 图 3).稳态分析结果表明, 在海拔梯度上, ln(SOC: TN)与ln(MBC: MBN)和ln(SOC: TP)与ln(MBC: MBP)之间存在H′大于1或不显著相关性(表 4).对微生物养分限制而言, 海拔梯度Ⅲ的元素阈值Ter C: N为6.54, 显著高于Ⅰ的2.51和Ⅱ的3.15(P < 0.05), 而海拔梯度Ⅰ、Ⅱ和Ⅲ的Ter C: P分别为11.03、10.49和12.08, 三者Ter C: P的差距未达到显著水平(P>0.05, 图 4).

|
(a) GMD表示几何平均直径, (b) MWD表示平均重量直径 图 3 不同海拔的团聚体稳定性指标 Fig. 3 Stability index of aggregates at different altitudes |
|
|
表 4 不同海拔土壤微生物群落水平上C: N和C: P的化学计量稳态1) Table 4 Stoichiometric steady state of C: N and C: P at the level of soil microbial communities at different altitudes |

|
Ter C:N表示碳氮阈值元素比, Ter C:P表示碳磷阈值元素比 图 4 不同海拔的元素阈值(Ter C: N和Ter C: P) Fig. 4 Threshold elemental ratio at different altitudes |
本研究的结果表明乔木、灌木和草本多样性在海拔上的变化不会显著影响土壤团聚体稳定性在海拔上的变化(P>0.05, 表 5).偏最小二乘路径模型(partial least squares path models, PLS-PM)结果表明, 海拔梯度的变化不会直接显著影响土壤团聚体稳定性, 但会通过影响土壤理化性质、微生物量和胞外酶活性等环境因子间接影响土壤团聚体稳定性.其中, 土壤团聚体稳定性指标与土壤理化性质(SOC、C: N和C: P, 路径系数为-0.176)和元素阈值(路径系数为-0.725)呈负相关关系, 与微生物量(MBN、MBP和MBC: MBN, 路径系数为0.158)和胞外酶活性[BG、NAG、BG: ACP和(NAG+LAP): ACP, 路径系数为1.398]均呈正相关关系.此外, 元素阈值除对团聚体稳定性有直接影响外, 也通过影响胞外酶活性而间接影响团聚体稳定性(路径系数为1.331, 图 5).
|
|
表 5 植物群落特征与土壤团聚体稳定性的相关性分析1) Table 5 Correlation analysis of plant community characteristics and soil aggregate stability |

|
椭圆内数字表示R2; 箭头上数字表示正(正数)或负(负数)影响的路径系数; 土壤理化性质指有机碳、碳氮比和碳磷比; 微生物量指微生物量氮、微生物量磷和微生物量碳氮比; 元素阈值指Ter C: N和Ter C: P; 胞外酶活性指BG、NAG和BG: ACP 图 5 土壤团聚体稳定性驱动因素的偏最小二乘路径模型 Fig. 5 Partial least squares path models (PLS-PM) of the drivers of soil aggregate stability |
团聚体分级对土壤酶活性有显著影响.总体上来说, 微团聚体酶活性显著高于大团聚体和中团聚体(图 6). 2.2节的分析结果表明, 酶活性是影响土壤团聚体稳定性的重要环境因子.因此本节进一步探究团聚体酶对GMD和NWD的影响.逐步回归结果表明(表 6), 土壤团聚体稳定性指标GMD、MWD与中团聚体NAG酶活性和小团聚体BG酶活性呈极显著相关(P < 0.01).

|
BG表示β-1, 4-葡萄糖苷酶, NAG表示β-1, 4-N-乙酰葡糖氨糖苷酶, LAP表示亮氨酸氨基肽酶, ACP表示酸性磷酸酶 图 6 不同海拔土壤团聚体酶活性 Fig. 6 Soil aggregate enzyme activity at different altitudes |
|
|
表 6 土壤团聚体酶活性对土壤团聚体稳定性的逐步回归方程1) Table 6 Stepwise regression equation of the relationship between soil aggregate enzyme activity and soil aggregate stability |
3 讨论 3.1 土壤理化性质及团聚体稳定性沿海拔梯度差异性
本研究的结果表明, 土壤有机碳、全氮和全磷含量随着海拔的升高而增加(表 2), 这与李丹维等[28]的研究结果一致.这主要是由于随着海拔的升高, 温度降低, 微生物活动减弱, 土壤有机碳全氮矿化速率减慢, 动植物残体分解速率减缓, 使得土壤中有机碳和全氮累积量增加[29].土壤团聚体稳定性结果表明, 低海拔(海拔梯度I)土壤团聚体稳定性显著高于高海拔土壤团聚体稳定性(图 3), 该研究结果与Zhu等[9]在四川雪宝顶山得出的结果相反.出现这种相异结果的原因一方面可能是由于研究区域的气候、纬度等方面存在差异, 另一方面也有可能是不同土壤微生物的群落构成、养分限制等引起的.土壤团聚体稳定性的影响因素复杂多样, 需综合考虑影响因素.
3.2 海拔梯度上土壤酶活性变化特征本研究中土壤碳获取酶(BG)、氮获取酶(NAG+LAP)和磷获取酶(ACP)的活性随着海拔升高而降低(图 2), 这与笔者以前的研究结果一致[30].值得注意的是, NAG和LAP同属于氮获取酶, 但二者对海拔的响应却截然不同(图 1).对此, 本研究提出了两种可能的解释.第一, 根据Miochaelis-Menten理论, 胞外酶的活性取决于底物的可利用性[12, 31].NAG主要在几丁质和其他β-1, 4-连接的葡糖胺聚合物中起作用, 而LAP主要水解多肽氮末端的亮氨酸和其他疏水氨基酸[32].由于两种N获取酶水解的底物不同, 微生物群落的策略也随之改变, 即低海拔的土壤更倾向可以产生NAG酶的微生物, 而高海拔的土壤更适合可以产生LAP酶的微生物.第二, 由海拔梯度引起的土壤微气候[33, 34]、凋落物输入[35, 36]和植物群落组成[37, 38]等的变化都会引起微生物群落的改变, 从而导致NAG和LAP在海拔上存在差异.
3.3 影响土壤团聚体稳定性的因素分析PLS-PM表明, 土壤团聚体稳定性在海拔上的变化主要受到了土壤胞外酶活性的影响(图 5).可能的解释是土壤微生物作为土壤团聚体形成最活跃的生物因素[6], 其生产的胞外酶活性可以指示微生物群落结构和养分需求的变化[25].具体而言, 低海拔土壤可能通过增加微生物量和多样性、根系生物量和分泌物、菌根菌丝体及其分泌的多糖类物质等多方面促进团聚体间的黏结作用, 从而提高团聚体稳定性[39].
微生物群落对养分的需求在海拔上的变化(即元素阈值Ter的变化)将通过调控胞外酶的产生, 来间接地影响土壤团聚体稳定性在海拔上的变化趋势.生态化学计量理论(ecological stoichiometry theory, EST)表明, 胞外酶的产生极大程度上受到微生物群落的养分限制的影响, 这是由于土壤养分(如SOC、TN和TP)存在海拔上的空间分布差异(表 2), 导致不同海拔的微生物将采取不同的策略调整其群落结构和功能, 以达到适应这种差异[40~42].具体来说, 微生物有以下3种方式来进行调整:①微生物通过储存过量养分或改变群落中优势物种进行调整[43, 44].这种调整取决于微生物群落的物种分布和生长速率, 使微生物群落趋向弱稳态或稳态[45].稳态分析表明, 在海拔梯度上, ln(SOC: TN)与ln(MBC: MBN)和ln(SOC: TP)与ln(MBC: MBP)之间存在H′大于1或无显著相关性(表 4), 这表明在本研究区域内3个海拔在微生物群落水平C: N和C: P均处于稳态.因此, 笔者认为本研究中没有发生此类调整.即微生物群落稳态在海拔上的变化不会影响土壤团聚体稳定性在海拔上的变化.②微生物通过调节其胞外酶的产量来进行调整[25, 46, 47].由于胞外酶的产生需要消耗能量和氮, 因此只有当调整后的收益远大于其调整成本, 才会发生这类调整.本研究的结果表明, BG: (NAG+LAP)平均值分别为1.25、1.83和2.27, 这意味着高海拔土壤具有相对较低的N获取酶活性(基于全球尺度值1.41), 这或许从侧面反映了随着海拔的升高, 微生物通过这类调整, N限制的状况得以改善.因此, 笔者认为, 本研究中发生了此类调整, 并且在高海拔上, 此类调整更为明显.这表明, 碳和氮获取酶(BG、NAG和LAP)的活性在海拔上的变化显著地影响团聚体稳定性在海拔上的变化.③微生物通过调节自身的生理能力(例如释放过量元素), 从而改变元素利用效率, 以达到调整的目的[48].EST理论表明, 较高的元素阈值Ter C: N伴随着更高的C利用效率和更低的N利用效率[44, 49].研究区域内(1 503~2 405 m)随着海拔的上升, SOC/TN的平均值分别为24.51、20.70和20.99, 而Ter C: N的值为2.51、3.15和6.54(图 4).表明微生物群落由较高的N利用率转变为较高的C利用率, 即高海拔微生物群落的N限制作用减弱.因此, 笔者认为, 本研究中也发生了此类调整, 这类调整旨在减少微生物对N的限制, 并且这类调整在高海拔更加明显.这表明, 微生物群落通过调节元素阈值(Ter C: N)也会影响土壤团聚体稳定性在海拔上的变化.
值得注意的是, 不同等级团聚体通过控制空气、水分以及有机质的可接触性影响土壤酶活性[17].本研究发现, 土壤微团聚体(< 0.25 mm)4种胞外酶活性显著高于大团聚体(>2 mm)酶活性(图 6).这或许是由于团聚体物理结构差异影响的, 微团聚体孔径较小, 养分和水分通过扩散作用进入的速度较快[50], 为酶促反应提供更多可利用的基质[51], 从而增强酶活性; 而大团聚体孔隙大, 团聚体稳定性低, 土壤酶活性易受环境变化影响[52], 从而导致酶活性降低.这与雷蕾等[53]在三峡库区得出的研究结论一致.进一步通过逐步回归分析得出, 中团聚体(0.25~2 mm)NAG酶和微团聚体(< 0.25 mm)BG酶是影响团聚体稳定性的重要因子(表 6).这可能是由于NAG酶主要水解几丁质[32], 而这些物质在中团聚体的形成和稳定性中起重要作用[7]; BG酶主要水解纤维素, 而多糖类物质在土壤微团聚体的稳定性中起关键作用, 能增加团聚体的稳定性[14, 54].
4 结论(1) 本研究反映了秦岭太白山区域不同海拔(1 503~2 451m)土壤团聚体稳定性指标GMD和MWD随着海拔的上升而降低, 其在海拔上的变化趋势主要受到土壤胞外酶活性的调控.
(2) 微生物通过调节胞外酶的相对产量和改变元素利用效率来进行调整, 使高海拔的N限制状况得以改善, 这种调整也会影响土壤团聚体稳定性在海拔上的变化.
(3) 中团聚体NAG酶和微团聚体BG酶是影响土壤团聚体稳定性的主要驱动因子, 团聚体的稳定性主要依靠多糖类和疏水氨基酸等物质的胶结作用.
| [1] | Davidson E A, Trumbore S E, Amundson R. Biogeochemistry: soil warming and organic carbon content[J]. Nature, 2000, 408(6814): 789-790. DOI:10.1038/35048672 |
| [2] |
陈仕奇, 吕盛, 高明, 等. 缙云山不同林分下土壤有机碳及矿化特征[J]. 环境科学, 2019, 40(2): 953-960. Chen S Q, Lü S, Gao M, et al. Characteristics of soil organic carbon and mineralization with different stands in Jinyun Mountain[J]. Environmental Science, 2019, 40(2): 953-960. |
| [3] | Six J, Conant R T, Paul E A, et al. Stabilization mechanisms of soil organic matter: Implications for C-saturation of soils[J]. Plant and Soil, 2002, 241(2): 155-176. DOI:10.1023/A:1016125726789 |
| [4] | Guo Z C, Zhang Z B, Zhou H, et al. The effect of 34-year continuous fertilization on the SOC physical fractions and its chemical composition in a Vertisol[J]. Scientific Reports, 2019, 9(1). DOI:10.1038/s41598-019-38952-6 |
| [5] | Carrillo-Saucedo S M, Gavito M E. Resilience of soil aggregation and exocellular enzymatic functions associated with arbuscular mycorrhizal fungal communities along a successional gradient in a tropical dry forest[J]. Mycorrhiza, 2020, 30(1): 109-120. DOI:10.1007/s00572-019-00928-9 |
| [6] |
王清奎, 汪思龙. 土壤团聚体形成与稳定机制及影响因素[J]. 土壤通报, 2005, 36(3): 415-421. Wang Q K, Wang S L. Forming and stable mechanism of soil aggregate and influencing factors[J]. Chinese Journal of Soil Science, 2005, 36(3): 415-421. DOI:10.3321/j.issn:0564-3945.2005.03.031 |
| [7] | 吴梦瑶, 陈林, 庞丹波, 等. 贺兰山不同海拔植被下土壤团聚体分布及其稳定性研究[J]. 水土保持学报, 2021, 35(2): 210-216. |
| [8] | Bordoloi R, Das B, Yam G, et al. Modeling of water holding capacity using readily available soil characteristics[J]. Agricultural Research, 2019, 8(3): 347-355. DOI:10.1007/s40003-018-0376-9 |
| [9] | Zhu M K, Yang S Q, Ai S H, et al. Artificial soil nutrient, aggregate stability and soil quality index of restored cut slopes along altitude gradient in southwest China[J]. Chemosphere, 2020, 246. DOI:10.1016/j.chemosphere.2019.125687 |
| [10] | Murugan R, Djukic I, Keiblinger K, et al. Spatial distribution of microbial biomass and residues across soil aggregate fractions at different elevations in the Central Austrian Alps[J]. Geoderma, 2019, 339: 1-8. DOI:10.1016/j.geoderma.2018.12.018 |
| [11] | Li G L, Vogel J, He Z L, et al. Association of soil aggregation with the distribution and quality of organic carbon in soil along an elevation gradient on Wuyi Mountain in China[J]. PLoS One, 2016, 11(3). DOI:10.1371/journal.pone.0150898 |
| [12] |
瞿晴, 徐红伟, 吴旋, 等. 黄土高原不同植被带人工刺槐林土壤团聚体稳定性及其化学计量特征[J]. 环境科学, 2019, 40(6): 2904-2911. Qu Q, Xu H W, Wu X, et al. Soil aggregate stability and its stoichiometric characteristics in Robinia pseudoacacia forest within different vegetation zones on the Loess Plateau, China[J]. Environmental Science, 2019, 40(6): 2904-2911. |
| [13] | Steffens M, Kölbl A, Kögel-Knabner I. Alteration of soil organic matter pools and aggregation in semi-arid steppe topsoils as driven by organic matter input[J]. European Journal of Soil Science, 2009, 60(2): 198-212. DOI:10.1111/j.1365-2389.2008.01104.x |
| [14] | Six J, Bossuyt H, Degryze S, et al. A history of research on the link between (micro)aggregates, soil biota, and soil organic matter dynamics[J]. Soil and Tillage Research, 2004, 79(1): 7-31. DOI:10.1016/j.still.2004.03.008 |
| [15] |
王兴, 钟泽坤, 张欣怡, 等. 长期撂荒恢复土壤团聚体组成与有机碳分布关系[J]. 环境科学, 2020, 41(5): 2416-2424. Wang X, Zhong Z K, Zhang X Y, et al. Relationship between the composition of soil aggregates and the distribution of organic carbon under long-term abandoned restoration[J]. Environmental Science, 2020, 41(5): 2416-2424. |
| [16] | Deng L, Kim D G, Peng C H, et al. Controls of soil and aggregate-associated organic carbon variations following natural vegetation restoration on the Loess Plateau in China[J]. Land Degradation & Development, 2018, 29(11): 3974-3984. |
| [17] | Six J, Paustian K. Aggregate-associated soil organic matter as an ecosystem property and a measurement tool[J]. Soil Biology and Biochemistry, 2014, 68: A4-A9. DOI:10.1016/j.soilbio.2013.06.014 |
| [18] |
唐志尧, 方精云, 张玲. 秦岭太白山木本植物物种多样性的梯度格局及环境解释[J]. 生物多样性, 2004, 12(1): 115-122. Tang Z Y, Fang J Y, Zhang L. Patterns of woody plant species diversity along environmental gradients on Mt. Taibai, Qinling Mountains[J]. Biodiversity Science, 2004, 12(1): 115-122. DOI:10.3321/j.issn:1005-0094.2004.01.014 |
| [19] | Nelson D W, Sommers L E. Total carbon, organic carbon, and organic matter[A]. In: Sparks D L, Page A L, Helmke P A, et al (Eds. ). Methods of Soil Analysis[M]. Madison: Soil Science Society of America, 1996. 961-1010. |
| [20] |
钟泽坤, 杨改河, 任成杰, 等. 黄土丘陵区撂荒农田土壤酶活性及酶化学计量变化特征[J]. 环境科学, 2020, 42(1): 411-421. Zhong Z K, Yang G H, Ren C J, et al. Effects of farmland abandonment on soil enzymatic activity and enzymatic stoichiometry in the Loess Hilly Region, China[J]. Environmental Science, 2020, 42(1): 411-421. |
| [21] | Brookes P C, Landman A, Pruden G, et al. Chloroform fumigation and the release of soil nitrogen: a rapid direct extraction method to measure microbial biomass nitrogen in soil[J]. Soil Biology and Biochemistry, 1985, 17(6): 837-842. DOI:10.1016/0038-0717(85)90144-0 |
| [22] | Vance E D, Brookes P C, Jenkinson D S. An extraction method for measuring soil microbial biomass C[J]. Soil Biology and Biochemistry, 1987, 19(6): 703-707. DOI:10.1016/0038-0717(87)90052-6 |
| [23] | Saiya-Cork K R, Sinsabaugh R L, Zak D R. The effects of long term nitrogen deposition on extracellular enzyme activity in an Acer saccharum forest soil[J]. Soil Biology and Biochemistry, 2002, 34(9): 1309-1315. DOI:10.1016/S0038-0717(02)00074-3 |
| [24] | Sterner R W, Elser J J. Ecological stoichiometry: the biology of elements from molecules to the biosphere[M]. Princeton: Princeton University Press, 2002. |
| [25] | Sinsabaugh R L, Hill B H, Follstad Shah J J. Ecoenzymatic stoichiometry of microbial organic nutrient acquisition in soil and sediment[J]. Nature, 2009, 462(7274): 795-798. DOI:10.1038/nature08632 |
| [26] | Kemper W D, Koch E J. Aggregate stability of soils from western United States and Canada: measurement procedure, correlations with soil constituents[R]. Washington: USDA, 1996. |
| [27] | Ali H E, Reineking B, Münkemüller T. Effects of plant functional traits on soil stability: intraspecific variability matters[J]. Plant and Soil, 2017, 411(1-2): 359-375. DOI:10.1007/s11104-016-3036-5 |
| [28] |
李丹维, 王紫泉, 田海霞, 等. 太白山不同海拔土壤碳、氮、磷含量及生态化学计量特征[J]. 土壤学报, 2017, 54(1): 160-170. Li D W, Wang Z Q, Tian H X, et al. Carbon, nitrogen and phosphorus contents in soils on Taibai mountain and their ecological stoichiometry relative to elevation[J]. Acta Pedologica Sinica, 2017, 54(1): 160-170. |
| [29] |
张广帅, 邓浩俊, 杜锟, 等. 泥石流频发区山地不同海拔土壤化学计量特征——以云南省小江流域为例[J]. 生态学报, 2016, 36(3): 675-687. Zhang G S, Deng H J, Du K, et al. Soil stoichiometry characteristics at different elevation gradients of a mountain in an area with high frequency debris flow: a case study in Xiaojiang Watershed, Yunnan[J]. Acta Ecologica Sinica, 2016, 36(3): 675-687. |
| [30] |
胡汗, 马寰菲, 白红英, 等. 秦岭太白山不同林带土壤微生物呼吸速率及其影响因素[J]. 生态学报, 2021, 41(1): 135-148. Hu H, Ma H F, Bai H Y, et al. The soil microbial respiration rate and its influencing factors in different forest belts of Taibai Mountain, Qinling Mountains[J]. Acta Ecologica Sinica, 2021, 41(1): 135-148. |
| [31] | Sinsabaugh R L, Lauber C L, Weintraub M N, et al. Stoichiometry of soil enzyme activity at global scale[J]. Ecology Letters, 2008, 11(11): 1252-1264. DOI:10.1111/j.1461-0248.2008.01245.x |
| [32] | Sinsabaugh R L, Gallo M E, Lauber C, et al. Extracellular enzyme activities and soil organic matter dynamics for northern hardwood forests receiving simulated nitrogen deposition[J]. Biogeochemistry, 2005, 75(2): 201-215. DOI:10.1007/s10533-004-7112-1 |
| [33] | Domínguez M T, Holthof E, Smith A R, et al. Contrasting response of summer soil respiration and enzyme activities to long-term warming and drought in a wet shrubland (NE Wales, UK)[J]. Applied Soil Ecology, 2017, 110: 151-155. DOI:10.1016/j.apsoil.2016.11.003 |
| [34] | Zhou X Q, Chen C R, Wang Y F, et al. Warming and increased precipitation have differential effects on soil extracellular enzyme activities in a temperate grassland[J]. Science of the Total Environment, 2013, 444: 552-558. DOI:10.1016/j.scitotenv.2012.12.023 |
| [35] | Bhattacharyya P, Roy K S, Neogi S, et al. Influence of elevated carbon dioxide and temperature on belowground carbon allocation and enzyme activities in tropical flooded soil planted with rice[J]. Environmental Monitoring and Assessment, 2013, 185(10): 8659-8671. DOI:10.1007/s10661-013-3202-7 |
| [36] | Yin H J, Li Y F, Xiao J, et al. Enhanced root exudation stimulates soil nitrogen transformations in a subalpine coniferous forest under experimental warming[J]. Global Change Biology, 2013, 19(7): 2158-2167. DOI:10.1111/gcb.12161 |
| [37] | Kardol P, Cregger M A, Campany C E, et al. Soil ecosystem functioning under climate change: plant species and community effects[J]. Ecology, 2010, 91(3): 767-781. DOI:10.1890/09-0135.1 |
| [38] | Steinauer K, Tilman D, Wragg P D, et al. Plant diversity effects on soil microbial functions and enzymes are stronger than warming in a grassland experiment[J]. Ecology, 2015, 96(1): 99-112. DOI:10.1890/14-0088.1 |
| [39] | Pérès G, Cluzeau D, Menasseri S, et al. Mechanisms linking plant community properties to soil aggregate stability in an experimental grassland plant diversity gradient[J]. Plant and Soil, 2013, 373(1-2): 285-299. DOI:10.1007/s11104-013-1791-0 |
| [40] | Zechmeister-Boltenstern S, Keiblinger K M, Mooshammer M, et al. The application of ecological stoichiometry to plant-microbial-soil organic matter transformations[J]. Ecological Monographs, 2015, 85(2): 133-155. DOI:10.1890/14-0777.1 |
| [41] | Buchkowski R W, Schmitz O J, Bradford M A, et al. Microbial stoichiometry overrides biomass as a regulator of soil carbon and nitrogen cycling[J]. Ecology, 2015, 96(4): 1139-1149. DOI:10.1890/14-1327.1 |
| [42] | Cleveland C C, Liptzin D. C: N: P stoichiometry in soil: is there a "Redfield ratio" for the microbial biomass?[J]. Biogeochemistry, 2007, 85(3): 235-252. DOI:10.1007/s10533-007-9132-0 |
| [43] | Fanin N, Fromin N, Buatois B, et al. An experimental test of the hypothesis of non-homeostatic consumer stoichiometry in a plant litter-microbe system[J]. Ecology Letters, 2013, 16(6): 764-772. DOI:10.1111/ele.12108 |
| [44] | Scott J T, Cotner J B, Lapara T M. Variable stoichiometry and homeostatic regulation of bacterial biomass elemental composition[J]. Frontiers in Microbiology, 2012, 3. DOI:10.3389/fmicb.2012.00042 |
| [45] | Yuan X B, Niu D C, Gherardi L A, et al. Linkages of stoichiometric imbalances to soil microbial respiration with increasing nitrogen addition: evidence from a long-term grassland experiment[J]. Soil Biology and Biochemistry, 2019, 138. DOI:10.1016/j.soilbio.2019.107580 |
| [46] | Allison S D, Vitousek P M. Responses of extracellular enzymes to simple and complex nutrient inputs[J]. Soil Biology and Biochemistry, 2005, 37(5): 937-944. DOI:10.1016/j.soilbio.2004.09.014 |
| [47] | Waring B G, Weintraub S R, Sinsabaugh R L. Ecoenzymatic stoichiometry of microbial nutrient acquisition in tropical soils[J]. Biogeochemistry, 2014, 117(1): 101-113. DOI:10.1007/s10533-013-9849-x |
| [48] | Manzoni S, Porporato A. Soil carbon and nitrogen mineralization: theory and models across scales[J]. Soil Biology and Biochemistry, 2009, 41(7): 1355-1379. DOI:10.1016/j.soilbio.2009.02.031 |
| [49] | Sinsabaugh R L, Manzoni S, Moorhead D L, et al. Carbon use efficiency of microbial communities: stoichiometry, methodology and modelling[J]. Ecology Letters, 2013, 16(7): 930-939. DOI:10.1111/ele.12113 |
| [50] | Six J, Paustian K, Elliott E T, et al. Soil structure and organic matter I. distribution of aggregate-size classes and aggregate-associated carbon[J]. Soil Science Society of America Journal, 2000, 64(2): 681-689. DOI:10.2136/sssaj2000.642681x |
| [51] | Steffens M, Kölbl A, Schörk E, et al. Distribution of soil organic matter between fractions and aggregate size classes in grazed semiarid steppe soil profiles[J]. Plant and Soil, 2011, 338(1-2): 63-81. DOI:10.1007/s11104-010-0594-9 |
| [52] |
钟晓兰, 李江涛, 李小嘉, 等. 模拟氮沉降增加条件下土壤团聚体对酶活性的影响[J]. 生态学报, 2015, 35(5): 1422-1433. Zhong X L, Li J T, Li X J, et al. Early effect of soil aggregates on enzyme activities in a forest soil with simulated N deposition elevation[J]. Acta Ecologica Sinica, 2015, 35(5): 1422-1433. |
| [53] |
雷蕾, 肖文发, 曾立雄, 等. 三峡库区2种马尾松混交林土壤团聚体酶活性分布特征[J]. 生态学报, 2020, 40(17): 6179-6188. Lei L, Xiao W F, Zeng L X, et al. Distribution of enzymatic activities within soil aggregates in two types of Pinus massoniana mixed plantations in the Three Gorges Reservoir area[J]. Acta Ecologica Sinica, 2020, 40(17): 6179-6188. |
| [54] |
罗晓虹, 王子芳, 陆畅, 等. 土地利用方式对土壤团聚体稳定性和有机碳含量的影响[J]. 环境科学, 2019, 40(8): 3816-3824. Luo X H, Wang Z F, Lu C, et al. Effects of land use type on the content and stability of organic carbon in soil aggregates[J]. Environmental Science, 2019, 40(8): 3816-3824. |