 2021, Vol. 42
2021, Vol. 42



2. 河海大学浅水湖泊综合治理与资源开发教育部重点实验室, 南京 210098
2. Key Laboratory of Integrated Regulation and Resource Development on Shallow Lakes, Ministry of Education, Hohai University, Nanjing 210098, China
近年来, 水体重金属污染事件频繁发生, 严重威胁着河湖水环境生态质量[1].这些重金属进入河流、湖泊等水环境中, 通过吸附和络合等过程最终会沉积富集在底泥中, 使得底泥成为重金属污染的“蓄积库”[2, 3].一旦水环境条件发生变化, 底泥中的重金属就会被释放到上覆水中, 通过食物链逐级向上富集, 最终进入人体, 对人类以及水生生物的健康产生严重威胁[4, 5].针对这一问题, 目前国内外研发出多种河湖底泥重金属污染的修复技术, 可分为原位修复和异位修复两大类.异位修复包括淋洗和电化学等, 通常来说这些方法所需要的工作量较大且成本较高, 同时存在破坏原地生态结构的风险[6, 7].原位修复与异位修复相比具有成本低和影响小等特点, 这其中针对浅水湖泊和湿地污染的植物修复技术由于具有较好的修复效果和应用潜力而逐渐受到人们的青睐[1, 2, 8].
沉水植物作为水生环境的重要组成部分, 具有修复效率高、单位成本低、生物量大和取材广泛等优点.同时相较于其他类别的水生植物, 沉水植物全株生长在水中, 与污染源的接触面积大, 因此可以同时去除底泥和水体中的污染物, 是水环境中重要的污染净化体[9, 10]. Xing等[11]在长江中下游24个重金属污染湖泊中发现大茨藻(Najas marina)是As和Cd的超累积植物, 最大富集量分别达到1 117.65 mg·kg-1和463.48mg·kg-1, 同时也发现金鱼藻(Ceratophyllum demersum)是Co、Cr和Fe的超累积植物.张饮江等[12]研究了金鱼藻、苦草和伊乐藻这3种沉水植物的去镉效果, 结果表明伊乐藻对水体镉的去除能力最强(去除率92%), 金鱼藻次之(去除率90%), 苦草最少(去除率66%).由此可以发现, 沉水植物对重金属有良好的富集能力, 且自身能够缓解重金属的毒害作用.然而, 目前对沉水植物的研究多集中在水体环境中[12, 13], 鲜见将底泥镉污染的耐受和富集机制结合研究, 同时缺乏对植物修复具体应用的建议[14~16].
目前很多关于植物重金属的研究多以定量检测的方法为主, 但是重金属在不同植物细胞中的分布并不相同, 仅依靠单位干/湿重下重金属的含量来表征植物对重金属的富集能力, 所得的研究结论并不全面[17].组织化学分析法可以为定量分析的结果提供补充, 有助于理解植物内重金属的分布和转移机制, 同时也可以更直观地了解植物细胞受毒害程度, 以便植物修复的实际应用[18, 19].基于此, 本文将以4种典型沉水植物黑藻(Hydrilla verticillata)、伊乐藻(Elodea canadensis)、菹草(Potamogeton crispus)和金鱼藻(Ceratophyllum demersum)为研究对象, 通过对植物各生长指标以及抗氧化酶活性的检测来分析植物对镉的耐受效应, 并借助生物/底泥富集因子和植物转运因子分析来了解植物的富集能力, 最后通过植物组织化学的方法来揭示镉在沉水植物体内的分布情况, 以期为沉水植物的实际应用提供理论支持.
1 材料与方法 1.1 实验材料及实验设计本研究所用水生植物为黑藻、伊乐藻、金鱼藻和菹草, 采自江苏省无锡市生态湿地园区, 挑选生长良好的幼苗, 在水生植物培育池中用1/4 Hoagland营养液预培养一周.实验所用底泥为市购池塘泥, 风干后去除其中的粗杂质, 并过100目筛后均质化.用CdCl2·2H2O(分析纯)配制5 g·L-1的镉储存液, 稀释不同的倍数倒入底泥中, 均匀混合并风干, 最终制成0、10、20和50 mg·kg-1这4个含量梯度的镉污染实验底泥(Cd-0、Cd-10、Cd-20和Cd-50)备用[16].实验采用高约20 cm的塑料花盆来模拟静态条件下的水体环境.每个花盆中称取80 g的实验底泥, 随后加入3 L Hoagland营养液.选取长势相同的植物幼苗以每盆5株的密度植入, 于室内培养42 d, 培养温度为(18±3)℃, 每天用2 000 μmol·(m2·s)-1的光照12 h/黑暗12 h, 每隔3 d用1/4 Hoagland营养液补充蒸发的水分, 实验设置3个平行.
1.2 植物、底泥及水体中镉含量测定本实验结束后取出植物样本, 用去离子水冲洗2~3遍, 擦干植物表面水分后置于烘箱中, 在60℃烘干至恒重, 磨碎后称取0.05 g(精确到0.000 1 g)进行微波消解.底泥样本在去除烂根及碎屑后, 冷冻干燥并研磨过100目筛, 随后称取0.12 g(精确至0.000 1 g)进行消解.样本消解后, 用ICP-OES(5100型, 美国安捷伦科技有限公司)测定镉含量[20].此外, 取培养结束后的上覆水样10 mL, 用0.45 μm滤膜过滤后, 同样测定其中的镉含量.
1.3 植物各指标的测定本研究采用直接浸提法测量叶绿素含量[21].取不包括叶脉的叶片0.25 g于比色管, 加入适量丙酮和乙醇, 避光静置后用紫外分光光度计进行测量, 进而计算出叶绿素a(Chla)、叶绿素b(Chlb)和总叶绿素(Chla+b)的含量(mg·g-1).
另取0.4 g植物鲜样, 加入缓冲液研磨后定容至10 mL, 离心后取上清液作为粗酶液待测.参考陈国梁[22]的实验方法, 分别用氮蓝四唑光化还原法、愈创木酚法和紫外吸光度法测定超氧化物歧化酶(SOD)活性、过氧化物酶(POD)活性和过氧化氢酶(CAT)活性.
1.4 植物组织内镉分布双硫腙显色法双硫腙可以与镉反应生成红褐色沉淀, 利用这一性质可以定性分析镉在植物各部分的富集量[23].分别取0.5 g植物根、茎和叶的鲜样, 用去离子水清洗3遍后, 放入染色剂中染色15 min, 染色剂由3 mg双硫腙、6 mL丙酮、2 mL蒸馏水与1~2滴冰醋酸制成.染色结束后用蒸馏水漂洗, 用徕卡光学显微镜观察并拍摄红褐色的镉双硫腙沉淀.
1.5 数据处理实验数据处理所需公式如下:
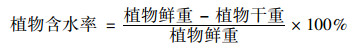
|
(1) |
生物/底泥富集因子(biota-sediment accumulation factor, BSAF)是衡量植物对底泥中重金属的富集累积情况, 可以反映植物对该重金属的去除能力.富集因子越大, 植物对重金属的去除效果就越强[24], 其公式为[25]:
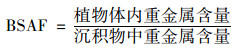
|
(2) |
转运系数(translocation factor, TF)可以表征植物根部向地上部分转移重金属的能力, 其公式为[26]:
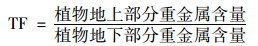
|
(3) |
用Excel 2013、Origin 8.0和SPSS18.0统计分析软件进行数据处理.图由Origin 8.0绘制, 表由Excel 2013生成, 利用SPSS 18.0进行单因素方差分析.
2 结果与分析 2.1 沉水植物生长情况及叶绿素含量变化情况暴露在不同浓度的底泥镉环境中42 d后, 金鱼藻在各浓度梯度中的生长状态均为最佳, 而菹草在各加标实验组中表现出了叶片失绿、枯萎和茎杆易断的现象.
由图 1可知黑藻和金鱼藻在Cd-10中的Chla和Chla+b含量均大于对照组(P < 0.05), 随后呈下降趋势, 而菹草的Chla和Chla+b含量则随浓度升高而逐渐降低.黑藻和伊乐藻的Chlb含量随镉浓度的升高而降低, 但菹草表现为一直上升, 从对照组的0.352mg·g-1到Cd-50中的0.376mg·g-1.黑藻和伊乐藻的Chla/Chlb比值呈现先上升后下降的趋势而菹草则表现为随浓度升高而下降的特点, 其中在Cd-10中Chla/Chlb比值有明显下降, 随后随底泥镉浓度升高而缓慢下降.金鱼藻的Chla/Chlb比值无明显变化趋势.

|
H为黑藻, Y为伊乐藻, Z为菹草, J为金鱼藻; 0、10、20和50代表含有不同含量(mg·kg-1)的镉底泥; **表示差异极显著(P < 0.01), *表示差异显著(P < 0.05), 下同 图 1 4种典型沉水植物在不同底泥镉浓度中的叶绿素含量 Fig. 1 Chlorophyll content of four typical submerged macrophytes in sediments with different cadmium concentrations |
黑藻和伊乐藻在Cd-10中超氧化物歧化酶(SOD)活性与对照组相比均有上升[图 2(a), (P < 0.05)], 而菹草和金鱼藻并没有明显变化.在Cd-50中, 除金鱼藻以外的3种植物的SOD活性相较对照组都有显著下降(P < 0.01).在本实验验中, 金鱼藻的SOD活性变化并不明显.

|
图 2 4种典型沉水植物在不同底泥镉浓度中的抗氧化酶活性 Fig. 2 Antioxidant enzyme activities of four typical submerged macrophytes in sediments with different cadmium concentrations |
黑藻、伊乐藻和菹草的过氧化物酶(POD)活性在Cd-10中均高于无污染的对照组[图 2(b)], 而在Cd-20中, 黑藻和伊乐藻的酶活性为16.03U·(min·g)-1和33.73U·(min·g)-1, 分别略高于对照组的15.77U·(min·g)-1和33.67U·(min·g)-1.在Cd-50中, 黑藻、伊乐藻和菹草的POD活性有明显下降.从整体来看, 金鱼藻的POD活性在Cd-10和Cd-20中随浓度增加有小幅度上升, 而在Cd-50中略有下降.
黑藻和伊乐藻的过氧化氢酶(CAT)活性在Cd-10和Cd-20都有所提高[图 2(c)].在Cd-50中, 两者的酶活性均下降, 其中伊乐藻的酶活性却仍高于对照组.菹草的CAT酶活性表现为先上升后下降的趋势, 在Cd-10中达到最大值.金鱼藻CAT的活性并没有表现出明显的变化.总的来说, 黑藻、伊乐藻和菹草的SOD、POD和CAT的活性都表现出先升高后下降的趋势, 而金鱼藻变化趋势较不明显.
2.3 沉水植物对底泥中镉的富集量如表 1所示, Cd-10和Cd-20的4种植物BSAF的大小均为:菹草>黑藻>伊乐藻>金鱼藻, 其中菹草在Cd-10下的BSAF达到2.58.在Cd-50中, 黑藻的富集能力最大, 伊乐藻其次, 菹草的BSAF为1.16, 富集能力有显著下降(P < 0.05).金鱼藻的BSAF一直呈下降趋势, 显著低于同等条件下的其它植物(P < 0.05), 且在3个浓度下的BSAF均未超过1.总体来说, 除金鱼藻外的3种沉水植物均表现出一定的底泥镉污染富集能力.
|
|
表 1 4种典型沉水植物体和底泥中的镉含量及生物/底泥富集因子(BSAF) Table 1 Cadmium content and biological/sediment enrichment factor (BSAF) in four typical submerged macrophytes and sediments |
本研究同时还分析了在Cd-50中生长的水生植物的地上部分和地下部分的镉含量, 结果如表 2所示.由于金鱼藻没有明显根系, 因此将其视为整株.从结果可以发现, 3种植物的地下部分镉含量都要大于地上部分, 转运系数(TF)均小于1, 同时其大小关系为:黑藻>伊乐藻>菹草, 说明这3种植物依靠植物根滤的机制来富集镉.值得注意的是:黑藻的TF最高, 为0.77; 菹草最低, 仅为0.28.
|
|
表 2 高镉污染环境中黑藻、伊乐藻和菹草地上和地下部分的镉含量 Table 2 Cadmium content of H.verticillata, E.canadensis, and P.crispus in high cadmium polluted environment |
为了更好地了解实验期间, 底泥中的镉向上覆水释放情况, 同时也对水样中的镉含量进行了测定.结果发现水样中基本没有检出镉污染物质, 仅在底泥镉污染浓度为50 mg·kg-1的黑藻和菹草实验组发现有少量镉存在, 检出浓度均为0.02 μg·L-1, 与底泥相比污染物含量很低, 因此本研究不考虑水体中的镉污染.
2.4 镉在沉水植物各部分的分布为了进一步探究重金属镉进入植物体内后的转运机制, 以及各植物对镉的富集机制, 本研究以Cd-50中的植物作为研究对象, 通过双硫腙显色法来观察镉在沉水植物根茎叶的分布情况, 同时又通过分析镉在沉水植物叶片中的亚细胞含量来揭示植物的富集机制.双硫腙试剂可与镉反应, 生成可以在光学显微镜下观察到的不溶性红棕色沉淀.根据这个原理, 将4种植物洗净并分成根、茎和叶这3部分, 分别进行双硫腙染色后, 用光学显微镜观察并拍照.
由于金鱼藻没有明显的根系, 因此只对3种植物的根部进行了染色实验.结果表明, 3种植物根部均呈现不同程度的红褐色沉淀, 其中菹草白根的红褐色沉淀最为明显, 且形成沉淀较大[图 3(c)], 说明其根系表现出了良好的阻隔镉向地上运输的作用.从茎细胞发现黑藻、伊乐藻和菹草中均能明显看到红褐色沉淀, 同时黑藻的细胞壁上附着有红褐色沉淀[图 3(a)].在菹草中, 植株茎细胞中的叶绿体密集均匀地分布有红褐色沉淀[图 3(c)], 说明此时镉已经对植株的叶绿体产生影响, 开始影响植株正常的生理活动.金鱼藻由于没有吸收过多重金属镉, 因此在茎部并没有看到很明显的红褐色沉淀.在4种植物叶片中均能看到明显的红棕色沉淀.伊乐藻叶片的某些细胞均被染色, 其中细胞壁的颜色最深, 同时还可以发现, 叶片中的某些细胞中叶绿体和液泡均有不同程度的显色[图 3(b)].在菹草叶片的叶脉中出现一个明显的深色沉淀[图 3(c)], 而在金鱼藻叶片中, 形成的沉淀较小且多聚集在细胞壁边[图 3(d)].

|
白色箭头所指为显微镜观察到的红褐色沉淀 图 3 4种典型沉水植物各部分双硫腙-镉沉淀光学显微镜照片 Fig. 3 Optical micrographs of dithizone-cadmium precipitation in various parts of four typical submerged macrophytes |
本研究从叶绿素含量以及抗氧化酶活性方面来分析4种沉水植物对底泥镉污染的耐受能力.叶绿素主要分布在叶绿体中, 是帮助植物吸收太阳光并进行光合作用的核心物质, 其含量的变化直接影响了植物的生长与发育.在本研究中, 除了菹草一直受到抑制以外, 其它3种植物各叶绿素含量均呈现先上升后下降的趋势, 这符合植物在镉环境中“低促高抑”的特点[27].该现象是由于镉对植物细胞分裂产生的干扰所致, 即在低浓度的镉环境中, 植物细胞的分裂速度会加快, 进而提高植物代谢能力[28].虽然低浓度的镉可以促进植物生长, 但是镉作为植物生长发育的非必需元素, 具有很强的生物毒性[29].当底泥镉浓度较高时(Cd≥20mg·kg-1), 4种植物的叶绿素含量已经受到不同程度的抑制, 直观表现为叶片失绿、茎杆萎蔫, 这种现象在Cd-50的菹草中尤为明显.抗氧化酶系统是植物的解毒机制之一, 主要包括过氧化物酶(POD)、过氧化氢酶(CAT)和超氧化物歧化酶(SOD)等[30].在本研究中, 黑藻和伊乐藻在较低的污染环境里(Cd-10和Cd-20)各氧化酶活性都有不同程度的升高, 说明植物自身解毒机制已经被激活.值得注意的是, 菹草在Cd-10环境下3种酶活性都有不同程度升高, 但是其生长增量却低于对照组, 这可能是由于镉对菹草的毒害作用已经超过了抗氧化系统自身调节能力, 使得植株的生长受到抑制[31, 32].
综合以上分析, 从底泥含镉浓度20mg·kg-1开始, 菹草的生长就已经受到抑制, 耐受能力在4种植物中最低, 同时金鱼藻的生长情况与底泥污染浓度并没有明显关系, 因此在本实验中, 植物的耐受能力为:黑藻>伊乐藻>菹草.
由于本研究是静态实验, 各样本的水体中均未检测出镉含量(部分Cd-50样本中检出有少量镉的存在, 但含量较少), 因此底泥是唯一镉污染源, 而在实际应用中, 当水体环境发生变化时, 底泥中的重金属镉会被释放, 此时植物地上部分也与污染物直接接触.关于沉水植物对水体镉的耐受能力, 王晓美等[15]的实验结果表明, 菹草叶片在0.5mg·L-1的镉溶液中开始出现腐烂发黑的现象, 而黑藻表现出来的抑制浓度为1 mg·L-1; 在林海等[33]的实验中, 黑藻在15mg·L-1的镉溶液中依然有50%的去除效果.同时又通过查阅相关研究, 总结了近年我国东南部河流湖泊不同介质的镉污染情况, 结果如下:①淮河淮南段的水体最高镉浓度为1.57 μg·L-1, 平均0.288 μg·L-1 [34]; ②太湖流域典型水系的底泥中镉浓度为0.479 mg·kg-1 [35]; ③太湖梅梁湾的底泥镉浓度为2.54mg·kg-1, 水体中镉浓度为0.065 μg·L-1 [36]; ④杭州市某河道底泥最高镉浓度为14.04 mg·kg-1, 平均3.61 mg·kg-1 [37]; ⑤山东省泰安市东平湖底泥最高镉浓度为0.348mg·kg-1, 平均0.285 mg·kg-1 [38].通过对比可以发现, 实际情况下的水体与底泥的镉污染浓度要远低于室内实验中沉水植物表现出来的抑制浓度.因此笔者推测, 本实验所选沉水植物都可以在大部分镉污染流域中生长.
3.2 沉水植物对底泥镉污染的富集转运作用从植物的生物/底泥富集因子(BSAF)可以看出, 在底泥含镉浓度小于20mg·kg-1时, 菹草的富集能力要略高于黑藻, 而在Cd-50中, 黑藻表现出来的富集能力要优于菹草.由于在实际情况中, 底泥的含镉浓度很难达到20~50mg·kg-1, 因此从富集能力来说, 黑藻和菹草都是比较理想的底泥修复植物.本研究中的金鱼藻并没有得到与前人在镉污染水体中进行的研究相类似的现象[27], 结果表明其BSAF的值小于1, 可能是由于金鱼藻退化的根系导致其并不能很好地与底泥中的镉有良好的接触[15, 16], 这也说明了根系对于修复底泥污染物的重要性.
从转运能力来说, 黑藻的TF为0.77, 接近Cd超富集植物的临界迁移系数标准(TF=1.00), 相比之下菹草更容易将镉富集在根部(TF=0.28).在实际应用中, 向地上部分转运能力强的黑藻, 可直接收获地上部分来收集重金属镉.而根部富集能力强的菹草, 需要连根拔除来收集污染物, 另外由于种植时间越长, 根系越发达, 在选用菹草修复底泥镉的时候需要及时收割和清理.
3.3 沉水植物体内镉污染的组织化学分布在本研究中, 通过双硫腙染色发现, 镉在不同植物、不同组织部位的分布都表现出了不均一性(如金鱼藻和伊乐藻叶片), 这也说明仅依靠BSAF和TF分析来了解植物的富集转运机制具有一定局限性.转运能力最低的菹草根部在经双硫腙染色之后出现边界明显且体积较大的红棕色沉淀, 这也与定量分析的结果一致.这种结果的出现可能是由于菹草的根部相对于其它水生植物可以产生更多的根际分泌物来固定重金属[39].从根部进入植物体内的镉, 会被茎输送至地上部分, 而茎作为物质传输的部分, 对镉的富集能力较差[15].值得注意的是, 在Cd-50中镉已经较均匀地分布在菹草茎部细胞的叶绿体中, 而在同样培养条件下的黑藻茎部仅可见一些清晰的沉淀附在细胞壁上, 且分布较不均匀.结合菹草的耐受性低于黑藻, 可以推断在一定重金属浓度中, 植物会将重金属固定在某个区域; 而当重金属的浓度超过植物可以承受的范围时, 重金属会对植物全身产生毒害作用.同时也建议在实际应用中, 为避免重金属镉对菹草正常生理活动产生影响, 应及时对菹草进行连根去除.
4 结论根系作用是沉水植物吸收和富集底泥镉污染的关键途径, 而根系严重退化的金鱼藻未能表现出明显的镉污染富集能力.其它3种植物在较低含镉污染底泥(Cd≤20mg·kg-1)的镉富集能力大小为:菹草≈黑藻>伊乐藻, 镉耐受能力大小为:黑藻>伊乐藻>菹草.菹草将多数富集的镉积累在植物根部, 而黑藻和伊乐藻却将多数镉转运至地上部分.进入地上部分的镉通过茎部被运输至叶片, 当毒性超出植物自身解毒能力的时候, 叶片中的镉便会进入细胞器中, 产生毒害作用.综合植物对镉污染的耐受和富集能力, 并结合实际情况, 最终将黑藻和菹草选为修复含镉底泥的理想植物.参考植物的转运能力, 建议在收获黑藻时可仅去除地上部分, 而菹草则需要定期连根去除.
| [1] |
李晶, 栾亚宁, 孙向阳, 等. 水生植物修复重金属污染水体研究进展[J]. 世界林业研究, 2015, 28(2): 31-35. Li J, Luan Y N, Sun X Y, et al. Research advances in remediation of heavy metal contaminated water bodies by aquatic plants[J]. World Forestry Research, 2015, 28(2): 31-35. |
| [2] | Giripunje M D, Fulke A B, Meshram P U. Remediation techniques for heavy-metals contamination in lakes: a mini-review[J]. Clean-Soil, Air, Water, 2015, 43(9): 1350-1354. DOI:10.1002/clen.201400419 |
| [3] | 张海锋, 胥焘, 黄应平, 等. 水生植物修复沉积物中重金属污染的机制及影响因素研究进展[J]. 亚热带水土保持, 2015, 27(1): 37-41. DOI:10.3969/j.issn.1002-2651.2015.01.009 |
| [4] | Akcil A, Erust C, Ozdemiroglu S, et al. A review of approaches and techniques used in aquatic contaminated sediments: metal removal and stabilization by chemical and biotechnological processes[J]. Journal of Cleaner Production, 2015, 86: 24-36. DOI:10.1016/j.jclepro.2014.08.009 |
| [5] | Jaiswal A, Verma A, Jaiswal P. Detrimental effects of heavy metals in soil, plants, and aquatic ecosystems and in humans[J]. Journal of Environmental Pathology, Toxicology and Oncology, 2018, 37(3): 183-197. DOI:10.1615/JEnvironPatholToxicolOncol.2018025348 |
| [6] |
梁欣. 旧厂房改造土壤重金属污染的原位与异位修复技术进展[J]. 中国资源综合利用, 2018, 36(5): 85-87. Liang X. Advances in in-situ and ectopic repair techniques for heavy metal pollution in soil of old factory buildings[J]. China Resources Comprehensive Utilization, 2018, 36(5): 85-87. DOI:10.3969/j.issn.1008-9500.2018.05.027 |
| [7] |
杨胜香, 田启建, 梁士楚, 等. 湘西花垣矿区主要植物种类及优势植物重金属蓄积特征[J]. 环境科学, 2012, 33(6): 2038-2045. Yang S X, Tian Q J, Liang S C, et al. Bioaccumulation of heavy metals by the dominant plants growing in huayuan manganese and lead/zinc mineland, Xiangxi[J]. Environmental Science, 2012, 33(6): 2038-2045. |
| [8] | Peng J F, Song Y H, Yuan P, et al. The remediation of heavy metals contaminated sediment[J]. Journal of Hazardous Materials, 2009, 161(2-3): 633-640. DOI:10.1016/j.jhazmat.2008.04.061 |
| [9] |
张萌, 刘足根, 李雄清, 等. 长江中下游浅水湖泊水生植被生态修复种的筛选与应用研究[J]. 生态科学, 2014, 33(2): 344-352. Zhang M, Liu Z G, Li X Q, et al. Studies of the selection and application of suitable hydrophyte species on lake restoration in the middle and lower reaches of Yangtze River[J]. Ecological Science, 2014, 33(2): 344-352. |
| [10] |
高海荣, 陈秀丽, 赵爱娟, 等. 5种沉水植物对重金属富集能力的对比研究[J]. 环境保护科学, 2016, 42(4): 101-105. Gao H R, Chen X L, Zhao A J, et al. Comparison of heavy metal accumulation by five submerged macrophytes[J]. Environmental Protection Science, 2016, 42(4): 101-105. |
| [11] | Xing W, Wu H P, Hao B B, et al. Bioaccumulation of heavy metals by submerged macrophytes: looking for hyperaccumulators in eutrophic lakes[J]. Environmental Science & Technology, 2013, 47(9): 4695-4703. |
| [12] |
张饮江, 易冕, 王聪, 等. 3种沉水植物对水体重金属镉去除效果的实验研究[J]. 上海海洋大学学报, 2012, 21(5): 784-793. Zhang Y J, Yi M, Wang C, et al. Reaserch on phytoremediation of cadmium contaminated water by three submerged macrophytes[J]. Journal of Shanghai Ocean University, 2012, 21(5): 784-793. |
| [13] |
贺艳. 植物材料对重金属污染水体的修复和轮叶黑藻(Hydrilla verticillata L. )对镉积累特性的研究[D]. 南京: 南京农业大学, 2016. He Y. Phytroemediation of contaminated water with plant materials and characterization of Cd accumulation in the aquatic plant Hydrilla verticillata[D]. Nanjing: Nanjing Agricultural University, 2016. |
| [14] |
谢佩君, 李铭红, 晏丽蓉, 等. 三种沉水植物对Cu、Pb复合污染底泥的修复效果[J]. 农业环境科学学报, 2016, 35(4): 757-763. Xie P J, Li M H, Yan L R, et al. Remediation of Cu and Pb co-polluted sediments by three submerged plants[J]. Journal of Agro-Environment Science, 2016, 35(4): 757-763. |
| [15] |
王晓美, 高培培, 孙旭淏, 等. 修复白洋淀镉污染水体的沉水植物筛选试验[J]. 广西植物, 2018, 38(10): 1382-1390. Wang X M, Gao P P, Sun X H, et al. Screening of submerged macrophytes for phytoremediation of cadmium-contaminated water in Baiyangdian Lake[J]. Guihaia, 2018, 38(10): 1382-1390. DOI:10.11931/guihaia.gxzw201711015 |
| [16] |
乔云蕾, 李铭红, 谢佩君, 等. 沉水植物对受重金属镉、锌污染的水体底泥的修复效果[J]. 浙江大学学报(理学版), 2016, 43(5): 601-609. Qiao Y L, Li M H, Xie P J, et al. A study on the absorption of cadmium and zinc in the water sediments with submerged plants[J]. Journal of Zhejiang University (Science Edition), 2016, 43(5): 601-609. |
| [17] | Seregin I V, Kozhevnikova A D. Distribution of cadmium, lead, nickel, and strontium in imbibing maize caryopses[J]. Russian Journal of Plant Physiology, 2005, 52(4): 565-569. DOI:10.1007/s11183-005-0084-8 |
| [18] | Seregin I V, Kozhevnikova A D. Histochemical methods for detection of heavy metals and strontium in the tissues of higher plants[J]. Russian Journal of Plant Physiology, 2011, 58(4): 721-727. DOI:10.1134/S1021443711040133 |
| [19] |
朱光旭, 肖化云, 郭庆军, 等. 铅锌尾矿污染区3种菊科植物体内重金属的亚细胞分布和化学形态特征[J]. 环境科学, 2017, 38(7): 3054-3060. Zhu G X, Xiao H Y, Guo Q J, et al. Subcellular distribution and chemical forms of heavy metals in three types of compositae plants from lead-zinc tailings area[J]. Environmental Science, 2017, 38(7): 3054-3060. |
| [20] |
陈秀梅. 酸溶-微波消解-电感耦合等离子体质谱法测定土壤及沉积物中的钽[J]. 环境科学导刊, 2020, 39(1): 85-90. Chen X M. Determination of Ta in soil and sediment samples in mixed acid using microwave digestion method by inductively coupled plasma-mass spectrometry[J]. Environmental Science Survey, 2020, 39(1): 85-90. |
| [21] |
兰心宇, 王军军, 阎蕴运, 等. 水生植物有翅星蕨(Microsorum pteropus)对镉的超富集能力及抗性生理研究[J]. 中国科学: 生命科学, 2017, 47(10): 1113-1123. Lan X Y, Wang J J, Yan Y Y, et al. Hyperaccumulation capacity and resistance physiology of Microsorum pteropus, an aquatic fern to cadmium[J]. Scientia Sinica Vitae, 2017, 47(10): 1113-1123. |
| [22] |
陈国梁. 沉水植物对砷的富集特征及机理研究[D]. 杭州: 浙江大学, 2014. Chen G L. The characteristics and mechanism of arsenic accumulation by submerged plants[D]. Hangzhou: Zhejiang University, 2014. |
| [23] | Clabeaux B L, Navarro D A G, Aga D S, et al. Cd tolerance and accumulation in the aquatic macrophyte, Chara australis: Potential use for charophytes in phytoremediation[J]. Environmental Science & Technology, 2011, 45(12): 5332-5338. |
| [24] | Lafabrie C, Major K M, Major C S, et al. Trace metal contamination of the aquatic plant Hydrilla verticillata and associated sediment in a coastal Alabama creek (Gulf of Mexico-USA)[J]. Marine Pollution Bulletin, 2013, 68(1-2): 147-151. DOI:10.1016/j.marpolbul.2012.11.045 |
| [25] | Huang P C, Tien C J, Sun Y M, et al. Occurrence of phthalates in sediment and biota: relationship to aquatic factors and the biota-sediment accumulation factor[J]. Chemosphere, 2008, 73(4): 539-544. DOI:10.1016/j.chemosphere.2008.06.019 |
| [26] |
何东, 邱波, 彭尽晖, 等. 湖南下水湾铅锌尾矿库优势植物重金属含量及富集特征[J]. 环境科学, 2013, 34(9): 3595-3600. He D, Qiu B, Peng J H, et al. Heavy metal contents and enrichment characteristics of dominant plants in a lead-zinc tailings in xiashuiwan of Hunan province[J]. Environmental Science, 2013, 34(9): 3595-3600. |
| [27] |
陈虹, 张颖, 方元平, 等. 镉对金鱼藻植株生长和抗氧化酶活性的影响[J]. 湖北农业科学, 2012, 51(5): 977-980. Chen H, Zhang Y, Fang Y P, et al. Cadmium-induced changes in development and oxidative metabolism in Ceratophyllum demersum plant[J]. Hubei Agricultural Sciences, 2012, 51(5): 977-980. DOI:10.3969/j.issn.0439-8114.2012.05.035 |
| [28] |
丁继军, 潘远智, 刘柿良, 等. 土壤重金属镉胁迫对石竹幼苗生长的影响及其机理[J]. 草业学报, 2013, 22(6): 77-85. Ding J J, Pan Y Z, Liu P L, et al. Effect and mechanisms of soil cadmium stress on Dianthus chinensis seedling growth[J]. Acta Prataculturae Sinica, 2013, 22(6): 77-85. |
| [29] | 杨红霞, 陈俊良, 刘崴. 镉对植物的毒害及植物解毒机制研究进展[J]. 江苏农业科学, 2019, 47(2): 1-8. |
| [30] |
贾茜茹, 刘奋武, 樊文华. 硅对Cd胁迫下黄瓜苗期光合及抗氧化酶系统的影响[J]. 水土保持学报, 2018, 32(4): 321-326. Jia Q R, Liu F W, Fan W H. Effects of silicon on photosynthesis and antioxidant enzymes of cucumber seedling under cadmium stress[J]. Journal of Soil and Water Conservation, 2018, 32(4): 321-326. |
| [31] |
赵会君, 梁昕昕, 魏玉清. 不同浓度的Cu、Mn、Zn胁迫对商陆叶片光合系统参数及抗氧化酶系统的影响[J]. 北方园艺, 2020(3): 120-127. Zhao H J, Liang X X, Wei Y Q. Effect of different concentrations of Cu, Zn and Mn on the photosynthetic and antioxidative system of pH ytolaccca americana[J]. Northern Horticulture, 2020(3): 120-127. |
| [32] |
万子栋, 高天鹏, 周玉霞, 等. 重金属复合胁迫下碱蓬萌发生长及富集特征[J]. 生物工程学报, 2020, 36(3): 493-507. Wan Z D, Gao T P, Zhou Y X, et al. Seed germination, bud growth and heavy-metal accumulation of Suaeda salsa[J]. Chinese Journal of Biotechnology, 2020, 36(3): 493-507. |
| [33] |
林海, 陈思, 董颖博, 等. 黑藻、狐尾藻对重金属铅、镉、铬、钒污染水体的修复[J]. 中国有色金属学报, 2017, 27(1): 178-186. Lin H, Chen S, Dong Y B, et al. Phytoremediation on heavy metal-polluted water of Pb, Cd, Cr and V by Hydrilla verticillata and Myriophyllum verticillatum[J]. The Chinese Journal of Nonferrous Metals, 2017, 27(1): 178-186. |
| [34] |
陈雅琼. 淮河淮南段重金属含量变化规律研究[J]. 广东化工, 2020, 47(5): 89-90. Chen Y Q. Study on the variation of heavy metal content in Huainan section of Huaihe river[J]. Guangdong Chemical Industry, 2020, 47(5): 89-90. DOI:10.3969/j.issn.1007-1865.2020.05.035 |
| [35] |
张杰, 郭西亚, 曾野, 等. 太湖流域河流沉积物重金属分布及污染评估[J]. 环境科学, 2019, 40(5): 2202-2210. Zhang J, Guo X Y, Zeng Y, et al. Spatial distribution and pollution assessment of heavy metals in river sediments from lake Taihu basin[J]. Environmental Science, 2019, 40(5): 2202-2210. DOI:10.3969/j.issn.1000-6923.2019.05.049 |
| [36] | Ni L X, Li D D, Su L L, et al. Effects of algae growth on cadmium remobilization and ecological risk in sediments of Taihu Lake[J]. Chemosphere, 2016, 151: 37-44. DOI:10.1016/j.chemosphere.2016.02.012 |
| [37] |
丁涛, 田英杰, 刘进宝, 等. 杭州市河道底泥重金属污染评价与环保疏浚深度研究[J]. 环境科学学报, 2015, 35(3): 911-917. Ding T, Tian Y J, Liu J B, et al. Assessment of heavy metal status in sediments and environmental dredging depth in Hangzhou[J]. Acta Scientiae Circumstantiae, 2015, 35(3): 911-917. |
| [38] | Wang Y Q, Yang L Y, Kong L H, et al. Spatial distribution, ecological risk assessment and source identification for heavy metals in surface sediments from Dongping Lake, Shandong, East China[J]. CATENA, 2015, 125: 200-205. DOI:10.1016/j.catena.2014.10.023 |
| [39] | Ma Y, Oliveira R S, Freitas H, et al. Biochemical and molecular mechanisms of plant-microbe-metal interactions: relevance for phytoremediation[J]. Frontiers in Plant Science, 2016, 7. DOI:10.3389/fpls.2016.00918 |