 2021, Vol. 42
2021, Vol. 42



2. 西南大学水产学院, 淡水鱼类资源与生殖发育教育部重点实验室, 重庆 400715
2. Key Laboratory of Freshwater Fish Reproductive and Development, Ministry of Education, College of Fisheries, Southwest University, Chongqing 400715, China
浮游生物是水环境中主要的生物类群之一, 主要包括浮游植物和浮游动物, 在水生生态系统中的物质循环和能量流动中起着决定性的作用[1].浮游植物是水环境中的初级生产者, 其群落结构直接或间接地受到物理、化学和生物等环境因子的影响[2], 同时其种类组成和分布等又能客观反映水环境的变化规律, 具有重要的指示作用[3], 因此, 众多研究中往往将浮游植物作为重要指标来评价水体质量[4].浮游动物作为水生生态系统的初级消费者, 在食物链中扮演着重要的角色, 其可能是滤食性鱼类捕食压力的有用指标[5], 影响水体环境质量.浮游动物的适应能力和对环境变化的敏感性与其种类有关[6], 如低营养级浮游动植物生态动力过程对环境变化的响应非常敏感[7].目前已有学者通过典范对应分析[8, 9](CCA)、冗余分析[10~12](RDA)、聚类分析[13~15]、Pearson相关性分析[16]、逐步回归分析[17]、相似性分析[18](ANOSIM)和非度量多维尺度分析[15, 19](NMDS)等方法对不同水域浮游动植物群落结构与环境因子的复杂关系进行了研究.
三峡水库蓄水以来, 受长江干流的顶托作用, 次级河流的流速减缓, 水体环境发生改变, 导致入库支流及回水区水体富营养化进而影响浮游动植物群落结构等, 这一系列问题引起了科研人员的密切关注[20].龙溪河是三峡库区一级次级河流, 属于三峡水库的典型支流.长寿湖水库(狮子滩水库)位于龙溪河下游, 是三峡水库上游最大且最重要的典型内陆水库型湖泊, 具有饮用、养殖、发电、旅游和防洪等多功能, 其水质的状况直接影响着三峡库区的水环境与生态安全.近年来, 由经济发展带来的外源污染物以及历史遗留的内源污染物共同导致了长寿湖水质的恶化, 已有研究表明, 长寿湖出现了水体富营养化[21]和水体重金属污染[22]等环境问题, 这会对水体浮游动植物的群落结构产生影响.雷波等[18]曾分析了该湖泊浮游藻类群落变化特征及主要环境影响因子, 但浮游动物的分布特征及影响因子, 以及利用浮游动植物进行的水质评价鲜见报道.因此, 本研究于不同季节在长寿湖采集水样和浮游动植物样品, 通过研究长寿湖浮游植物和浮游动物的群落结构以及生态学特征及其与环境因子的关系, 评价长寿湖目前的水质状况, 以期为了解长寿湖生态系统结构与功能以及对该区域生态环境保护等提供科学依据, 同时长寿湖作为三峡库区上游最重要的大型湖泊系统, 本研究对于了解库区其他湖泊系统而言, 具有重要的科学价值.
1 材料与方法 1.1 研究区概况长寿湖水库(29°54′~30°04′N, 107°14′~107°27′E, 图 1)是我国西南地区最大的人工淡水湖泊, 其位于重庆市长寿区境内, 于20世纪50年代建成.长寿湖属于中亚热带湿润季风气候区, 其气候特征为:四季分明、气候温和且雨水充沛.其年平均气温17.7℃, 年平均降水量超过1 000 mm.长寿湖水域面积为65.5 km2, 总库容积为10亿m3, 其最大水深和平均水深分别为40 m和15 m, 多年平均流量为47.5 m3·s-1, 径流深496.4 mm.

|
图 1 研究区域示意 Fig. 1 Schematic diagram of the study area |
本研究于不同季节:冬季(2016年12月)、春季(2017年4月)和夏季(2017年7月)在长寿湖选取5个采样点(S1~S5)采集水样和浮游动植物样品(图 1).采样点的设置根据水库的地形地貌设在大坝出水口(S1)、西北区(S2)、滞水区(S3)、东北区(S4)和湖心区(S5).
将25#(64 μm)浮游植物网置于水体表层0.5 m, 同时在水体表层1 m处放置13#(112 μm)浮游动物网[1], 以1 m·s-1的船速缓慢在湖中拖网收集浮游植物与浮游动物定性样品, 拖网时间根据浮游动植物的多少而定.同时, 浮游植物定量分析样品在各站点于水体表层0.5 m处用有机玻璃采水器采集1 L水样, 浮游动物定量分析样品于水体表层0.5 m和1.0 m均匀混合采集20 L水样, 并用25#(64 μm)浮游生物网过滤浓缩后收集.用于浮游植物与浮游动物定性和定量分析的样品, 分别按照体积比1.5%加入鲁哥氏液(Lugol's solution)和5%福尔马林溶液固定, 并尽快运送至实验室, 沉淀浓缩后在显微镜下进行鉴定和计数, 方法参考文献[23].
水体理化指标测定方法如下:水体总氮(TN)、总磷(TP)、叶绿素a(Chla)、硝态氮(NO3--N)、亚硝态氮(NO2--N)和铵态氮(NH4+-N)的测定方法见文献[24]; 使用Sievers InnovOx Laboratory TOC Analyzer(GE, USA)测定溶解性有机碳(DOC); 采用多参数水质分析仪现场测定水体pH、氧化还原电位(Eh)、水温(T)和溶解氧(DO)等指标.
1.3 数据处理与分析本文中, 用优势度(Y)来表示优势种, 将Y大于0.02的物种定为优势种[6], 优势度[25]计算见公式(1).采用生物多样性指数包括Shannon-Wiener指数(H′)、Pielous均匀度指数(J)和Margalef丰富度指数(D)对水质进行评价, 具体计算公式如下[18]:

|
(1) |
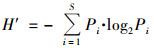
|
(2) |
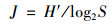
|
(3) |
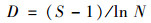
|
(4) |
式中, 第i物种的出现频率用fi表示, 第i物种个体数量与总个体数量的比值用Pi表示, N表示同一样品中浮游动植物的总个体数, Ni为第i种浮游动植物的个体数, S是样品中浮游动植物的总种类数.
Pearson相关性分析:选取10个生物指标包括浮游动植物丰度(ZA、PA)、浮游动植物生物量(ZB、PB)、浮游动植物Shannon-Wiener指数(ZH、PH)、浮游动植物Pielous均匀度指数(ZJ、PJ)和浮游动植物Margalef丰富度指数(ZD、PD), 以及TN、TP、Chla、NH4+-N、NO3--N、NO2--N、DOC、pH、Eh、T和DO共11个理化指标, 进行标准差标准化处理后采用SPSS21.0软件对长寿湖浮游动植物与环境因子做Pearson相关性分析, 其中R值表示相关性大小, P值表示两个变量间的显著性, 一般要求P < 0.05有统计学意义[4].
运用软件SPSS 21.0进行数据处理和方差分析, 用Origin 9.0软件作图.
2 结果与讨论 2.1 长寿湖水体理化特性长寿湖不同季节的水体理化特性如表 1所示, 除NH4+-N和TP之外, 其他环境因子均呈现出显著的季节性差异(P < 0.05).如, 冬季pH、Eh和NO2--N质量浓度显著低于春季和夏季(P < 0.05), TN质量浓度为(2.24±0.34)mg·L-1, 显著高于春季(P < 0.05); 春季NO3--N质量浓度为(0.13±0.07)mg·L-1, 显著低于冬季和夏季(P < 0.05), DOC、DO和Chla质量浓度显著高于夏季(P < 0.05); 夏季T值为(28.98±0.43)℃, 显著高于冬春两季(P < 0.05).但从空间分布上来看, 整个研究期间各采样点的水体环境因子无显著性差异(P>0.05).
|
|
表 1 长寿湖不同季节水体理化特性1) Table 1 Physical and chemical indexes of water body of Changshou Lake in different seasons |
2.2 长寿湖浮游动植物群落结构 2.2.1 物种组成
长寿湖浮游动、植物的鉴定结果表明, 浮游植物共鉴定出8门107种, 其最主要组成群落为蓝藻门, 相对丰度达61.03%; 其次是隐藻门, 相对丰度为19.04%.长寿湖共鉴定出纤细席藻、点状平裂藻和类颤藻鱼腥藻3种优势浮游植物种群.浮游动物共鉴定出4门82种, 主要由轮虫组成, 相对丰度达88.11%; 其次为原生动物, 相对丰度为6.08%.统计分析发现, 长寿湖浮游动物的优势种为无节幼体、长肢多肢轮虫、萼花臂尾轮虫、螺形龟甲轮虫、前节晶囊轮虫和盖氏晶囊轮虫.
另外, 不同季节的浮游动植物相对丰度和优势种群存在较大差异(图 2和表 2).冬季浮游植物为绿藻-蓝藻-硅藻群落, 相对丰度分别为34.03%、27.85%和22.58%, 其中优势种群包括点状平裂藻、颗粒直链藻和纤细席藻等6种浮游植物(表 2).到了春季转变为隐藻-硅藻-绿藻群落, 其相对丰度分别为26.24%、22.69%和17.90%, 优势种群也发生了较大变化, 主要包括卵形隐藻、啮蚀隐藻和短圆柱单肠藻等7种浮游植物.而到了夏季蓝藻相对丰度上升至71.57%, 成为主要的优势群落[图 2(a)], 优势种群主要包括纤细席藻、钝顶螺旋藻和卷曲鱼腥藻等7种浮游植物(表 2), 这是因为蓝藻在较高的温度、强光和静水环境中更具有生长优势[23].本研究中长寿湖冬、春季节硅藻群落有一定的优势, 这与冬、春季节水温较低有利于硅藻生长有关[23].春季水体NH4+-N、DOC和NO2--N的浓度较高, 因此, 喜生于有机物和氮丰富水体的隐藻更具有生长优势.有研究结果表明, 绿藻在富营养化湖泊中占优势[26], 与本研究中冬、春季绿藻为优势群落的结果一致.

|
图 2 长寿湖不同季节各门类浮游植物和浮游动物相对丰度 Fig. 2 Relative abundances of phytoplankton and zooplankton in different seasons in Changshou Lake |
|
|
表 2 长寿湖不同季节浮游动植物优势种及优势度 Table 2 Dominant species and the degrees of zooplankton and phytoplankton in different seasons in Changshou Lake |
对于浮游动物而言, 不同季节其丰度和优势群落变化不大, 从图 2(b)可以看出, 冬春夏3个季节轮虫均为主要组成群落, 这与童子湖水库浮游动物的研究结果一致[27], 其相对丰度均最高, 分别为61.66%、85.76%和89.62%.这是由于轮虫个体小, 发育时间快, 生命周期短, 鱼类对其影响较小, 对水体营养水平的变化敏感, 轮虫的种类和数量随湖泊营养水平的上升逐渐增加[28], 而长寿湖目前处于富营养化状态[18], 轮虫因此成为其优势群落.浮游动物优势种群在不同季节有差异, 冬季主要包括无节幼体和长额象鼻溞等5种浮游动物, 春季包括螺形龟甲轮虫和前节晶囊轮虫等8种浮游动物, 夏季包括剪形臂尾轮虫和萼花臂尾轮虫等7种浮游动物(表 2).
2.2.2 浮游动植物丰度、生物量和多样性指数长寿湖浮游动植物的丰度、生物量的统计结果(图 3)和时空分布差异性分析表明, 浮游动植物丰度和生物量呈现了显著的季节性差异(P < 0.05), 这与其他研究结果一致[10, 29~31], 但整个研究期间, 空间差异均不显著(P>0.05).如图 3(a)所示, 夏季的浮游植物丰度最高, 春季次之, 冬季最低, 而浮游动物春季丰度最高, 和冬、夏季有显著的差异性.由图 3(b)可知, 春季浮游动、植物生物量均显著高于冬季.由图 3可知, 浮游动植物的丰度和生物量随季节变化的趋势大致相同, 这与浮游动物常滤食浮游植物有关.按照国内湖泊富营养化评价标准[32], 即浮游植物丰度小于3×105 cells·L-1为贫营养, 介于(3~10)×105 cells·L-1为中营养, 大于10×105 cells·L-1为富营养, 可知长寿湖在冬季为中营养状态, 春、夏季为富营养状态.同时选取长寿湖的TN、TP和Chla为评价指标, 根据OECD富营养化单因子(Chla) 评价标准[33, 34], 即Chla < 3mg·m-3为贫营养; Chla介于3~11 mg·m-3为中营养; Chla介于11~78 mg·m-3为富营养; Chla>78 mg·m-3为严重富营养化, 得到长寿湖在冬季为中营养状态, 春季为严重富营养化状态, 夏季为富营养状态, 与利用浮游植物丰度评价水质富营养化的结果相一致.并根据水体富营养化的临界浓度(TN为0.2mg·L-1, TP为0.02 mg·L-1)[33]可得, 长寿湖冬、春季均远远超过对应的TN和TP的临界浓度值, 水体基本处于富营养化状态.而本研究中浮游动植物优势种为富营养型湖泊水库中常见的种类, 如颗粒直链藻、卷曲鱼腥藻、啮蚀隐藻、螺形龟甲轮虫、前节晶囊轮虫和剪形臂尾轮虫等, 符合水体营养状态的评价结果.

|
图 3 长寿湖不同季节浮游动植物的丰度和生物量 Fig. 3 Abundances and biomass of zooplankton and phytoplankton in Changshou Lake in different seasons |
浮游动植物的种类数以及生物多样性指数的时空差异性分析表明, 整个研究期间, 其空间差异均不显著(P>0.05), 浮游动物多样性指数季节性差异不显著(P>0.05), 这与张冬融等[17]对浮游动物的研究结果一致, 但浮游植物存在显著的季节性差异(P < 0.05).由表 3可得, 冬春两季浮游植物生物多样性指数均显著高于夏季(P < 0.05), 这可能是由于夏季丰水期的到来破坏了长寿湖水体稳定性[18], 干扰漂浮性浮游植物的生长, 以及夏季较高的温度和光照条件有利于蓝藻类优势种群大量生长, 从而抑制其他种类的生长, 导致生物多样性减少, 如蓝藻的大量生长抑制硅藻的生长[35].浮游动物的种类数春季最大, 与冬夏两季差异显著(P < 0.05), 这主要由于冬季温度较低, 不利于浮游动物的生长, 以及夏季轮虫优势种群的大量生长抑制了其他浮游动物的生长, 导致其种类数降低.
|
|
表 3 长寿湖不同季节浮游动植物的生物多样性指数1) Table 3 Biodiversity indexes of zooplankton and phytoplankton in different seasons in Changshou Lake |
生物多样性指数是评价水体质量状况的指标之一, 已有学者基于此进行水质评价的研究[36].本研究利用浮游植物群落的生物多样性指数对长寿湖进行水质评价(表 4), 3项指数的值越大, 表明多样性越高[37], 水质越好.由Shannon-Wiener指数结果可得, 长寿湖冬、春季处于清洁-寡污状态, 夏季水质类型为β-中污型水质; Pielous均匀度指数结果表明, 冬春夏3个季节水质属于清洁-寡污型; 根据Margalef丰富度指数发现, 长寿湖水质冬、春季处于中度污染, 夏季处于重污染, 但有向中度污染过渡的趋势. 总体而言, 长寿湖冬、春季生物多样性指数差异不显著, 二者均显著大于夏季(P < 0.05), 冬、春季节长寿湖水质处于清洁-寡污状态, 夏季处于中度污染状态.由周边地区工农业的快速发展带来的大量营养盐、有机物等外源污染物可能是导致长寿湖水质变差的主要原因.
|
|
表 4 基于Shannon-Wiener指数、Pielous指数和Margalef指数的水质评价标准[36] Table 4 Water quality evaluation criteria of the Shannon-Wiener index, Pielou's index, and Margalef index |
2.3 长寿湖浮游动植物群落结构与环境因子的响应关系
湖泊多个环境因子在时间和空间上影响浮游动植物的群落结构. 有研究表明, 浮游动植物群落结构受水温、营养盐、盐度、pH、水深、溶解氧、总氮、叶绿素a、透明度和总磷等[31, 38, 39]的影响, 如适当的温度可促进浮游动植物群落发展[40], 溶解的微量元素供应可改变浮游植物的群落结构, 增加人为来源的养分负荷, 降低生态多样性和加剧群落结构的演替[41].
本研究中Pearson相关性分析结果表明(表 5), 冬季ZB与PB显著正相关, PH、ZH与Eh均具有显著的相关性, PA与DO、T有相关关系, PJ与DO、TP具有显著的负相关关系, ZB与Chla呈现显著的正相关, ZH与NO2--N表现为显著的负相关关系.春季浮游动物生物指标均与浮游植物丰度和生物量没有显著相关的关系, PA和ZH与NO3--N显著正相关, PA、PB与Chla显著正相关, ZH与DO显著负相关. 夏季ZA与PB显著负相关, ZH、ZD均与PA具有显著正相关关系, PA、ZH和ZD与NH4+-N, 以及PB与NO3--N之间均具有显著的正相关关系, PA、PD和ZD均与DOC呈现显著的负相关关系.
|
|
表 5 浮游动植物的生物指标与环境因子的相关性1) Table 5 Correlations between the biological indexes of zooplankton and phytoplankton and environmental factors |
浮游动植物之间的关系密切而复杂, 是水体中所有环境因子共同作用的结果, 浮游动物对浮游植物的下行控制效应, 对浮游动植物的群落结构有重要的影响.本研究中, 冬季浮游动物生物量与浮游植物生物量具有显著的正相关关系, 春季浮游动植物生物指标之间无显著的相关关系, 夏季浮游动物丰度与浮游植物生物量显著负相关, 浮游动物生物多样性指数与浮游植物丰度显著正相关, 说明长寿湖浮游动物对浮游植物的下行控制效应存在季节性差异, 夏季该效应尤为突出, 夏季浮游动物的生物多样性与浮游植物密切相关, 可能是夏季有利于浮游动植物的生长, 浮游动物对浮游植物的摄食压力增大导致的.DO是衡量水体自净能力的指标之一, 浮游动植物的生长均需要消耗氧气进行呼吸作用, 本研究中冬季浮游植物Pielous均匀度指数与DO正相关, 张丹[42]的研究结果也得到了相同的结论.本研究发现, 环境因子NO2--N不利于浮游动物的生长, 冬、春季浮游动植物指标与TP、DO存在显著的负相关关系, 这表明NO2--N、TP和DO在特定环境下对浮游动植物的生长也存在抑制作用.窦勇等[1]的研究中也提出了氮盐对浮游动物的生长可能存在一定的毒害作用.本研究中, 夏季DOC对浮游动植物的生长有影响, DOC代表水体中溶解性有机质(DOM)的含量, DOM通过吸收紫外光从而对浮游植物的光合作用产生影响, 同时DOM和水体的物质发生反应, 以及含有的弱酸弱碱官能团影响水体的pH[43], 导致水环境中物质的生物有效性发生改变, 从而直接或间接地影响浮游植物的群落结构.夏季浮游动植物指标与铵态氮、硝态氮有关, 氮盐作为调节浮游植物生长的营养元素之一, 对浮游植物的生长有一定的促进作用; 冬季浮游动植物的生物指标与Eh有关, 因为Eh可能通过影响氨氮转化速率与效率[44]等控制浮游植物的生长, 李德亮等[45]对大通湖的研究结果也表明, Eh是影响浮游植物群落结构的主要因子.冬、春季浮游动植物生物量与Chla正相关, 已有研究表明水体中浮游植物现存量的变化可通过Chla的浓度来反映[46].且浮游动物滤食浮游植物, 因此, 可推断DOC、氮盐、Eh和Chla通过改变浮游植物群落结构, 从而间接影响浮游动物的群落结构.
3 结论(1) 本次对浮游动植物的鉴定结果表明, 浮游植物共有8门107种, 其群落结构呈现季节性的变化规律, 体现为绿藻-蓝藻-硅藻群落(冬季)转变为隐藻-硅藻-绿藻群落(春季), 而夏季以蓝藻群落为主.浮游动物共鉴定出4门82种, 轮虫为其最主要组成群落.
(2) 通过浮游动植物的指示作用以及利用生物多样性指数进行水质评价, 可得冬、春季节长寿湖水质处于清洁-寡污状态, 夏季处于中度污染状态, 长寿湖整体处于富营养化状态.
(3) 影响长寿湖浮游动植物群落结构的环境因子包括Chla、DOC、TP、NO2--N、NO3--N、NH4+-N、DO、Eh和T.不同季节影响浮游动植物群落结构的环境因子存在差异, 因此不同时期需综合考虑浮游动植物与环境因子相互影响的关系, 采取有效措施防治长寿湖的环境污染.
| [1] |
窦勇, 霍达, 姜智飞, 等. 海河入海口表层水体浮游生物群落特征及与环境因子的相关性研究[J]. 生态环境学报, 2016, 25(4): 647-655. Dou Y, Huo D, Jiang Z F, et al. Study of the community characteristics of plankton and their correlation with the environmental factors in surface water in Haihe estuary[J]. Ecology and Environmental Sciences, 2016, 25(4): 647-655. |
| [2] |
薄芳芳, 杨虹, 左倬, 等. 上海公园水体夏季浮游植物群落与环境因子的关系[J]. 生态学杂志, 2009, 28(7): 1259-1265. Bo F F, Yang H, Zuo Z, et al. Relationship between phytoplankton community in summer and environmental factors in Shanghai park lakes[J]. Chinese Journal of Ecology, 2009, 28(7): 1259-1265. |
| [3] | Lepistö L, Holopainen A L, Vuoristo H. Type-specific and indicator taxa of phytoplankton as a quality criterion for assessing the ecological status of Finnish boreal lakes[J]. Limnologica, 2004, 34(3): 236-248. DOI:10.1016/S0075-9511(04)80048-3 |
| [4] |
李娜, 周绪申, 孙博闻, 等. 白洋淀浮游植物群落的时空变化及其与环境因子的关系[J]. 湖泊科学, 2020, 32(3): 772-783. Li N, Zhou X S, Sun B W, et al. Spatiotemporal variation of phytoplankton community and its relationship with environmental factors in the Lake Baiyangdian[J]. Journal of Lake Sciences, 2020, 32(3): 772-783. |
| [5] | Li Y, Chen F Z. Are zooplankton useful indicators of water quality in subtropical lakes with high human impacts?[J]. Ecological Indicators, 2020, 113. DOI:10.1016/j.ecolind.2020.106167 |
| [6] |
夏品华, 马健荣, 李存雄, 等. 红枫湖水库冬春季浮游生物群落与环境因子的典范对应分析[J]. 环境科学研究, 2011, 24(4): 378-386. Xia P H, Ma J R, Li C X, et al. Canonical correspondence analysis between the plankton community and environmental factors in Hongfeng Lake reservoir in winter and spring[J]. Research of Environmental Sciences, 2011, 24(4): 378-386. |
| [7] |
魏皓, 赵伟, 罗晓凡, 等. 北冰洋浮游生物空间分布及其季节变化的模拟[J]. 海洋学报, 2019, 41(9): 65-79. Wei H, Zhao W, Luo X F, et al. Simulation of spatial distribution and seasonal variation of plankton in the Arctic Ocean[J]. Acta Oceanologica Sinica, 2019, 41(9): 65-79. DOI:10.3969/j.issn.0253-4193.2019.09.006 |
| [8] |
杨丽, 张玮, 尚光霞, 等. 淀山湖浮游植物功能群演替特征及其与环境因子的关系[J]. 环境科学, 2018, 39(7): 3158-3167. Yang L, Zhang W, Shang G X, et al. Succession characteristics of phytoplankton functional groups and their relationships with environmental factors in Dianshan Lake, Shanghai[J]. Environmental Science, 2018, 39(7): 3158-3167. |
| [9] |
王海邻, 刘玉飞, 任玉芬, 等. 北京市河流秋季浮游动物群落特征分析[J]. 环境科学, 2019, 40(8): 3568-3576. Wang H L, Liu Y F, Ren Y F, et al. Analysis of river zooplankton community characteristics in autumn in Beijing[J]. Environmental Science, 2019, 40(8): 3568-3576. |
| [10] |
杨杰青, 欧阳珑玲, 唐峰华, 等. 海南西北部近岸海域浮游动物群落结构与环境因子的关系[J]. 中国水产科学, 2020, 27(2): 236-249. Yang J Q, Ouyang L L, Tang F H, et al. Relationship between the zooplankton community structure and en-vironmental factors offshore of northwest Hainan Island, China[J]. Journal of Fishery Sciences of China, 2020, 27(2): 236-249. |
| [11] |
许志, 陈小华, 沈根祥, 等. 上海河道浮游植物群落结构时空变化特征及影响因素分析[J]. 环境科学, 2020, 41(8): 3621-3628. Xu Z, Chen X H, Shen G X, et al. Spatial and temporal variation of phytoplankton community structure and its influencing factors in Shanghai river channels[J]. Environmental Science, 2020, 41(8): 3621-3628. DOI:10.3969/j.issn.1000-6923.2020.08.043 |
| [12] |
王振方, 张玮, 杨丽, 等. 异龙湖不同湖区浮游植物群落特征及其与环境因子的关系[J]. 环境科学, 2019, 40(5): 2249-2257. Wang Z F, Zhang W, Yang L, et al. Characteristics of phytoplankton community and its relationship with environmental factors in different regions of Yilong Lake, Yunnan Province, China[J]. Environmental Science, 2019, 40(5): 2249-2257. |
| [13] |
柳丽华, 左涛, 陈瑞盛, 等. 2004年秋季长江口海域浮游植物的群落结构和多样性[J]. 海洋水产研究, 2007, 28(3): 112-119. Liu L H, Zuo T, Chen R S, et al. Community structure and diversity of phytoplankton in the estuary of Yangtse River in autumn[J]. Marine Fisheries Research, 2007, 28(3): 112-119. |
| [14] |
毕洪生, 孙松, 高尚武, 等. 渤海浮游动物群落生态特点Ⅰ. 种类组成与群落结构[J]. 生态学报, 2000, 20(5): 715-721. Bi H S, Sun S, Gao S W, et al. The ecological characteristics of the zooplankton community in the Bohai sea Ⅰ. Species composition and community structure[J]. Acta Ecologica Sinica, 2000, 20(5): 715-721. DOI:10.3321/j.issn:1000-0933.2000.05.001 |
| [15] | Sidik M J, Rashed-Un-Nabi M, Hoque A. Distribution of phytoplankton community in relation to environmental parameters in cage culture area of Sepanggar Bay, Sabah, Malaysia[J]. Estuarine, Coastal and Shelf Science, 2008, 80(2): 251-260. DOI:10.1016/j.ecss.2008.08.004 |
| [16] | Lv X M, Zhang J, Liang P, et al. Phytoplankton in an urban river replenished by reclaimed water: features, influential factors and simulation[J]. Ecological Indicators, 2020, 112. DOI:10.1016/j.ecolind.2020.106090 |
| [17] |
张冬融, 徐佳奕, 徐兆礼, 等. 杭州湾南岸海域春秋季浮游动物分布特征与主要环境因子的关系[J]. 生态学杂志, 2014, 33(8): 2115-2123. Zhang D R, Xu J Y, Xu Z L, et al. Distribution pattern of zooplankton in the south part of Hangzhou Bay during spring and autumn in relation to main environmental factors[J]. Chinese Journal of Ecology, 2014, 33(8): 2115-2123. |
| [18] |
雷波, 刘朔孺, 张方辉, 等. 三峡水库上游长寿湖浮游藻类的季节变化特征及关键环境影响因子[J]. 湖泊科学, 2017, 29(2): 369-377. Lei B, Liu S R, Zhang F H, et al. Seasonal phytoplankton dynamics and its relationship with key environmental factors in Lake Changshou, upper reaches of the Three Gorges Reservoir[J]. Journal of Lake Sciences, 2017, 29(2): 369-377. |
| [19] |
张庆吉, 王业宇, 王金东, 等. 骆马湖浮游植物演替规律及驱动因子[J]. 环境科学, 2020, 41(4): 1648-1656. Zhang Q J, Wang Y Y, Wang J D, et al. Succession pattern of phytoplankton and its drivers in Lake Luoma, Jiangsu Province[J]. Environmental Science, 2020, 41(4): 1648-1656. |
| [20] |
潘晓洁, 刘诚, 万成炎, 等. 三峡水库典型支流河口浮游植物群落与水环境相关性分析[J]. 水生态学杂志, 2016, 37(6): 7-13. Pan X J, Liu C, Wan C Y, et al. Phytoplankton community structure in typical tributary coves of Three Gorges Reservoir and its relationship to the water environment[J]. Journal of Hydroecology, 2016, 37(6): 7-13. |
| [21] |
杨志敏, 熊海灵, 张晟, 等. 重庆长寿湖富营养化评价及氮磷平衡研究[J]. 水土保持学报, 2005, 19(2): 73-75, 109. Yang Z M, Xiong H L, Zhang S, et al. Assessment of eutrophication and balance of N and P in Changshou Reservoir[J]. Journal of Soil and Water Conservation, 2005, 19(2): 73-75, 109. DOI:10.3321/j.issn:1009-2242.2005.02.019 |
| [22] |
周莉萍, 金相灿, 卢少勇, 等. 重庆市长寿湖重金属污染评价[J]. 农业环境科学学报, 2010, 29(7): 1355-1361. Zhou L P, Jin X C, Lu S Y, et al. Access of heavy metals pollution of Changshouhu in Chongqing, China[J]. Journal of Agro-Environment Science, 2010, 29(7): 1355-1361. |
| [23] | 赵文. 水生生物学[M]. 北京: 中国农业出版社, 2005. |
| [24] | 国家环境保护总局. 水和废水监测分析方法[M]. (第四版). 北京: 中国环境科学出版社, 2002. |
| [25] |
郭沛涌, 沈焕庭, 刘阿成, 等. 长江河口浮游动物的种类组成、群落结构及多样性[J]. 生态学报, 2003, 23(5): 892-900. Guo P Y, Shen H T, Liu A C, et al. The species composition, community structure and diversity of zooplankton in Changjiang estuary[J]. Acta Ecologica Sinica, 2003, 23(5): 892-900. DOI:10.3321/j.issn:1000-0933.2003.05.008 |
| [26] |
张洁, 杨娟. 三峡库区汛期浮游藻类与水质特征研究[J]. 生态科学, 2017, 36(1): 55-63. Zhang J, Yang J. A study on phytoplankton and water quality characteristics in the Three Gorges Reservoir at flood season[J]. Ecological Science, 2017, 36(1): 55-63. |
| [27] | 胡莲, 彭建华, 邹曦, 等. 童子湖水库生态监测与湿地恢复的生态效应研究[A]. 见: 2018中国环境科学学会科学技术年会论文集(第一卷)[C]. 合肥: 中国环境科学学会, 2018. 9. |
| [28] |
林秋奇, 胡韧, 段舜山, 等. 广东省大中型供水水库营养现状及浮游生物的响应[J]. 生态学报, 2003, 23(6): 1101-1108. Lin Q Q, Hu R, Duan S S, et al. Reservoir trophic states and the response of plankton in Guangdong Province[J]. Acta Ecologica Sinica, 2003, 23(6): 1101-1108. DOI:10.3321/j.issn:1000-0933.2003.06.010 |
| [29] |
林青, 由文辉, 徐凤洁, 等. 滴水湖浮游动物群落结构及其与环境因子的关系[J]. 生态学报, 2014, 34(23): 6918-6929. Lin Q, You W H, Xu F J, et al. Zooplankton community structure and its relationship with environmental factors in Dishui Lake[J]. Acta Ecologica Sinica, 2014, 34(23): 6918-6929. |
| [30] |
刘晓曦, 陈丽, 蒋伊能, 等. 抚仙湖浮游植物群落时空变化特征及其与环境因子的关系[J]. 湖泊科学, 2020, 32(3): 793-803. Liu X X, Chen L, Jiang Y N, et al. Spatiotemporal variation of phytoplankton communities and their relationship with environmental factors in Lake Fuxian[J]. Journal of Lake Sciences, 2020, 32(3): 793-803. |
| [31] |
吴利, 唐会元, 龚云, 等. 三峡水库正常运行下库区干流浮游动物群落特征研究[J]. 水生态学杂志, 2021, 42(1): 58-65. Wu L, Tang H Y, Gong Y, et al. Temporal-spatial distribution of zooplankton community in the main stem of Three Gorges Reservoir under normal operation[J]. Journal of Hydroecology, 2021, 42(1): 58-65. |
| [32] |
原居林, 尹文林, 沈锦玉, 等. 南太湖浮游植物特征及其富营养化评价[J]. 生态学杂志, 2009, 28(11): 2197-2201. Yuan J L, Yin W L, Shen J Y, et al. Phytoplankton and its relations with eutrophication of southern Taihu Lake[J]. Chinese Journal of Ecology, 2009, 28(11): 2197-2201. |
| [33] |
李如忠, 刘科峰, 钱靖, 等. 合肥市区典型景观水体氮磷污染特征及富营养化评价[J]. 环境科学, 2014, 35(5): 1718-1726. Li R Z, Liu K F, Qian J, et al. Nitrogen and phosphate pollution characteristics and eutrophication evaluation for typical urban landscape waters in Hefei City[J]. Environmental Science, 2014, 35(5): 1718-1726. |
| [34] |
陈豪, 左其亭, 张永勇, 等. 淮河中上游浮游植物群落结构特征及营养状况评价[J]. 中国农村水利水电, 2019(2): 1-6, 10. Chen H, Zuo Q T, Zhang Y Y, et al. Phytoplankton community structure and nutrition status evaluation in the upper and middle reaches of the Huaihe River[J]. China Rural Water and Hydropower, 2019(2): 1-6, 10. |
| [35] |
刘静, 杜桂森, 刘晓端, 等. 密云水库的浮游生物群落[J]. 西北植物学报, 2004, 24(8): 1485-1488. Liu J, Du G S, Liu X D, et al. Study on plankton community in Miyun reservoir[J]. Acta Botanica Boreali-Occidentalia Sinica, 2004, 24(8): 1485-1488. DOI:10.3321/j.issn:1000-4025.2004.08.024 |
| [36] |
董芳, 周贤君, 张辉, 等. 贵阳市小车河浮游生物群落结构与水环境因子的关系[EB/OL]. 基因组学与应用生物学. http://kns.cnki.net/kcms/detail/45.1369.q.20200213.1346.002.html, 2020-10-30. Dong F, Zhou X J, Zhang H, et al. Plankton community structure and its relation to the environmental factors of Xiaoche River in Guiyang City, Guizhou[EB/OL]. Genomics and Applied Biology. http://kns.cnki.net/kcms/detail/45.1369.q.20200213.1346.002.html, 2020-10-30. |
| [37] |
王珊, 于明, 刘全儒, 等. 东江干流浮游植物的物种组成及多样性分析[J]. 资源科学, 2013, 35(3): 473-480. Wang S, Yu M, Liu Q R, et al. Phytoplankton species composition and biodiversity in Dongjiang River[J]. Resources Science, 2013, 35(3): 473-480. |
| [38] |
石晓丹, 阮晓红, 邢雅囡, 等. 苏州平原河网区浅水湖泊冬夏季浮游植物群落与环境因子的典范对应分析[J]. 环境科学, 2008, 29(11): 2999-3008. Shi X D, Ruan X H, Xing Y N, et al. Canonical correspondence analysis between phytoplankton community and environmental factors in winter and summer in Shallow Lakes of plain river network areas, Suzhou[J]. Environmental Science, 2008, 29(11): 2999-3008. |
| [39] |
刘存歧, 孔祥玲, 张治荣, 等. 基于RDA的白洋淀浮游植物群落动态特征分析[J]. 河北大学学报(自然科学版), 2016, 36(3): 278-285. Liu C Q, Kong X L, Zhang Z R, et al. Dynamics of phytoplankton community in Baiyangdian Lake based on the redundancy analysis(RDA)[J]. Journal of Hebei University (Natural Science Edition), 2016, 36(3): 278-285. DOI:10.3969/j.issn.1000-1565.2016.03.010 |
| [40] |
王硕, 杨涛, 李小平, 等. 渭河流域浮游动物群落结构及其水质评价[J]. 水生生物学报, 2019, 43(6): 1333-1345. Wang S, Yang T, Li X P, et al. Zooplankton community structure and the water quality in the Wei River Basin[J]. Acta Hydrobiologica Sinica, 2019, 43(6): 1333-1345. |
| [41] | Nwankwegu A S, Li Y P, Huang Y N, et al. Nutrient addition bioassay and phytoplankton community structure monitored during autumn in Xiangxi Bay of Three Gorges Reservoir, China[J]. Chemosphere, 2020, 247. DOI:10.1016/j.chemosphere.2020.125960 |
| [42] |
张丹. 不同水质类别城市河道浮游生物群落结构分析及其多样性的研究[D]. 上海: 华东师范大学, 2011. Zhang D. Study on community structure and diversity of plankton in urban rivers of different water ouality category[D]. Shanghai: East China Normal University, 2011. |
| [43] |
白英臣. 天然溶解有机质和环境污染物的相互作用——以铜、汞离子和卡马西平为例[D]. 贵阳: 中国科学院地球化学研究所, 2007. Bai Y C. Interaction between natural dissolved organic matter and environmental contaminants: copper, mercury ion and carbamazepine[D]. Guiyang: Institute of Geochemistry, Chinese Academy of Sciences, 2007. |
| [44] |
赵秀侠, 卢文轩, 李静, 等. 青龙湖冬季浮游植物群落结构及其与环境因子的关系[J]. 水生态学杂志, 2020, 41(2): 83-90. Zhao X X, Lu W X, Li J, et al. Winter phytoplankton community structure and its relationships with environmental factors in Qinglong Lake, Anhui Province[J]. Journal of Hydroecology, 2020, 41(2): 83-90. |
| [45] |
李德亮, 张婷, 肖调义, 等. 大通湖浮游植物群落结构及其与环境因子关系[J]. 应用生态学报, 2012, 23(8): 2107-2113. Li D L, Zhang T, Xiao D Y, et al. Phytoplankton's community structure and its relationships with environmental factors in an aquaculture lake, Datong Lake of China[J]. Chinese Journal of Applied Ecology, 2012, 23(8): 2107-2113. |
| [46] |
刘朋超, 王卫军, 骆启豪, 等. 山东半岛南北两侧海域真核浮游生物群落特征及与环境因子的相关性分析[J]. 海洋与湖沼, 2019, 50(4): 876-885. Liu P C, Wang W J, Luo Q H, et al. Characteristics of eukaryotic plankton community and its correlation with environmental factors in the northern and southern seas of Shandong Peninsula[J]. Oceanologia et Limnologia Sinica, 2019, 50(4): 876-885. |