 2021, Vol. 42
2021, Vol. 42



2. 中国科学院南京地理与湖泊研究所湖泊与环境国家重点实验室, 南京 210008
2. State Key Laboratory of Lake and Environment, Nanjing Institute of Geography&Limnology, Chinese Academy of Sciences, Nanjing 210008, China
氮是藻类生长的重要限制因子之一, 水体氮浓度升高会导致藻类大量增殖, 引起水体富营养化和蓝藻水华的发生[1], 给水生生态系统带来巨大危害[2].近几十年来, 关于地表水环境中活性氮去除过程的相关研究备受国内外学者广泛关注.长期以来, 反硝化一直被认为是水体去除无机氮的唯一生物途径. 20世纪90年代发现的厌氧氨氧化反应打破了这一传统观念[3].目前国际上对海洋中厌氧氨氧化过程的研究已经十分成熟, 发现其在海洋氮循环中的脱氮贡献率可高达79%[4], 但目前国内外对淡水系统厌氧氨氧化反应及其对脱氮贡献率的研究偏少.
在浅水湖泊中, 由于底泥与水体接触比例更高, 反硝化脱氮的能力更强[5].而且浅水湖泊更容易发生富营养化[6], 湖泊的富营养化程度会影响湖体内的反硝化作用.有研究表明, 富营养化湖泊的反硝化速率明显大于贫营养化湖泊和中度营养化湖泊[7].但是关于浅水, 富营养化湖泊中厌氧氨氧化作用对湖泊脱氮的贡献研究较少.反硝化菌是异养菌, 厌氧氨氧化菌是自养菌, 有研究表明, 浅水湖泊中反硝化对氮去除的贡献率要远高于厌氧氨氧化[8], 因为沉积物与水界面更容易受到溶解氧和外源有机污染物的干扰, 导致厌氧氨氧化菌活性受到抑制而被淘汰[9].不同湖泊或同一湖泊不同湖区受人类活动的影响程度不同, 其富营养化程度也不同, 其对反硝化和厌氧氨氧化速率的影响还需要深入研究.
太湖是一个大型浅水湖泊, 是长三角地区的重要水源地之一[10].近几十年来, 由于人类活动的影响, 日益增加的工业, 农业和生活污水排放加速了太湖水体富营养化进程[11, 12], 不同湖区受人类影响程度的不同, 其富营养化程度也大不相同.竺山湾、大浦口和梅梁湾等河道入湖的湖区污染尤为严重, 而湖心区受人类影响小, 污染相对较小.此外, 不同湖区由于水质和沉积物的性质差异巨大, 其主导的水生植物也大不相同, 北部和西部为藻型湖区, 以浮游植物为主, 而东部胥口湾和东太湖为草型湖区, 以沉水植物为主[10].大量研究表明蓝藻和沉水植物都能显著影响到水体氮的脱除[8, 13], 因此研究不同类型湖区的脱氮速率对深入了解氮的收支有重要的意义.关于太湖反硝化和厌氧氨氧化已经开展了相关研究工作[13, 14], 但是研究多集中在单个湖区, 缺乏对于不同湖区之间脱氮能力的比较.同时, 以往的很多研究多是使用泥浆培养的方法测定反硝化和厌氧氨氧化潜力[15, 16].泥浆实验的曝气过程破坏了底泥的微环境, 改变了原位条件下的溶解氧, 同时底泥中大量的氨氮被搅起从而高估了厌氧氨氧化对脱氮的贡献率.相比之下, 无扰动泥柱流动培养的方法能更大程度地模拟湖泊的原位条件, 减少了人为操作的影响[17].徐徽等[14]使用流动培养结合同位素示踪的方法测定了冬季梅梁湾反硝化和厌氧氨氧化的速率和贡献率, 但是在计算反硝化速率时使用的是Rysgaard等[18]提出的反硝化公式, 其将厌氧氨氧化产生的29N2归结到了反硝化作用中从而低估了厌氧氨氧化对脱氮的贡献率.
基于以上问题, 于2020年4月初和6月末采集了太湖不同湖区的无扰动泥柱进行流动培养, 采用15N同位素示踪和膜进样质谱仪结合的技术测定反硝化和厌氧氨氧化速率, 分析造成不同湖区脱氮速率差异的原因, 以期深入认识湖泊系统氮素的归趋和循环机制.
1 材料与方法 1.1 采样点位太湖是一个生态类型较为复杂的大型浅水湖泊, 既有藻型湖区, 又有草型湖区, 还有草藻过渡湖区.梅梁湾, 竺山湾和大浦口是藻型湖区, 同时竺山湾和大浦口又是河道湖水入流的湖区, 胥口湾是草型湖区, 贡湖湾是过渡区, 湖心区是沉积物较少的湖区[10]. 4月的采样点位覆盖了4个湖区, 包括梅梁湾、贡湖湾、竺山湾和湖心区, 6月增加了两个湖区(大浦口和胥口湾), 其中梅梁湾和湖心区由于原来的点位未采到形状良好的泥柱而有所变动.
4月的采样点位为梅梁湾8号、湖心区7号、贡湖湾3号和竺山湾4号. 6月的采样点位选择梅梁湾1号(8号点位由于清淤工程未采到性状良好的泥柱而改为1号点)、贡湖湾3号、竺山湾4号、大浦口2号、湖心区5号(7号点未采到性状良好的泥柱而改为5号)和胥口湾6号(图 1).

|
图 1 泥柱采样点位示意 Fig. 1 Sampling sites of sediment cores |
2020年的4月和6月于太湖部分湖区用柱状采泥器采集无扰动泥柱, 每个点位采集3根泥柱作为平行, 泥柱内径90 mm, 泥柱中沉积物高度约为10 cm左右.同步采集50 L上覆水, 使用YSI 6600 V2(美国Yellow Stone公司)多参数水质测定仪测定上覆水pH、DO和温度等理化性质.
1.3 室内培养实验为了尽量模拟太湖的原位环境, 本研究采用流动培养法开展实验.流动培养法是目前被广泛应用的接近于原位情况下的反硝化厌氧氨氧化速率的测定方法.在流动培养的实验中, 除了额外添加的15N外, 其他条件都尽量与原位环境相一致(包括无扰动泥柱的性状, 除硝氮以外的其他营养盐浓度, 有机碳浓度和溶氧浓度), 设置的温度与采样时的原位温度一致, 并进行了恒温控制.在每个泥柱中都添加了等量的15N, 使硝氮本底值低的湖区加完15N后依然低, 而本底值高的湖区加完15N后依然高.
泥柱尽量无扰动运回以避免破坏沉积物的性状和避免氧气的侵蚀, 运回泥柱后避光预培养一天以消耗沉积物表面的硝酸盐.泥柱采用流动培养的方法测定反硝化和厌氧氨氧化速率[16].在有机玻璃管上端塞入带有O型硅胶圈的活塞, 使沉积物柱样上端形成无顶空的气密环境, 上覆水体积约为270 mL, 活塞上连接硅胶管作为进水管和出水管, 保证硅胶管连接处不漏气, 进水管距表层沉积物约1 cm, 进水管低于出水管, 通过流动培养的进出水循环形成垂直及水平方向的混合水流, 使上覆水充分混合.流速通过蠕动泵调节, 蠕动泵放置在出水口, 流速设置为1.5 mL·min-1, 保证上覆水溶解氧变化较小, 而且此流速下界面形态稳定.每根泥柱配备一个10 L的进水袋, 进水袋中的原位水添加Na15NO3进行处理, 使添加完后15N的浓度为2 mg·L-1, 15N的最终丰度根据添加同位素后的硝氮浓度与未添加同位素的浓度的差值计算[15]. 泥柱置于恒温水浴槽中进行恒温培养, 温度设置为采样时湖水的温度.培养时间3 d, 每24 h取一次样, 使用12 mL的Labco质谱专用玻璃顶空瓶采集进水和出水样品, 每个玻璃瓶内注射0.1 mL质量分数为50%的ZnCl2杀死微生物来保存样品, 用50mL塑料离心管采集进水和出水样品, 用0.45 μm的滤膜过滤以备测定无机营养盐和DOC, 在测定前水样保存于-20℃的冰箱中.
1.4 样品测定与分析水体营养盐的测定使用Skalar连续流动分析仪(荷兰Skalar公司, SAN++型), DOC浓度通过TOC测定仪测定, 叶绿素a使用热乙醇提取法测定[19]. 采用膜进样质谱仪测定培养实验中水体28N2、29N2和30N2[20].数据分析和作图使用Origin 9.0和Arcgis 10.2软件.
1.5 反硝化与厌氧氨氧化速率及贡献率的计算反硝化速率计算公式[21]:
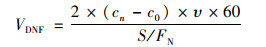
|
(1) |
式中, VDNF为反硝化速率[μmol·(m2·h)-1], cn为第n次取样时30N2的浓度(μmol·L-1), c0为初始样品的30N2浓度(μmol·L-1), υ为蠕动泵流速(mL·min-1), S为泥柱底面积(m2), FN为15N丰度, 60为时间换算因子.
厌氧氨氧化速率计算公式[22]:
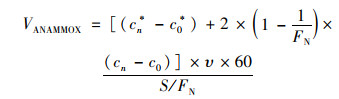
|
(2) |
式中, VANAMMOX为反硝化速率[μmol·(m2·h)-1], cn为第n次取样时30N2的浓度(μmol·L-1), c0为初始样品的30N2浓度(μmol·L-1), cn*为第n次取样时29N2的浓度(μmol·L-1), c0*为初始样品的29N2浓度(μmol·L-1), υ为蠕动泵流速(mL·min-1), S为泥柱底面积(m2), FN为15N丰度, 60为时间换算因子.
反硝化和厌氧氨氧化对脱氮贡献率的计算:
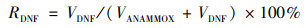
|
(3) |
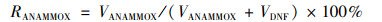
|
(4) |
式中, RDNF为反硝化贡献率, RANAMMOX为厌氧氨氧化贡献率.
1.6 统计分析为了探究太湖不同湖区脱氮速率的控制因子, 将通过流动培养法所测得的反硝化, 厌氧氨氧化速率和它们对脱氮的相对贡献率与采样时对应的原位环境因子进行相关性分析.统计分析软件为SPSS 19.
2 结果与分析 2.1 太湖水质的空间差异不同湖区的水质情况如表 1所示.春季水温在20℃左右, 总体来看竺山湾的营养盐浓度最高, 其中硝氮的浓度达到1.5 mg·L-1, 氨氮和亚硝氮分别达到0.226 mg·L-1和0.098 mg·L-1, 正磷酸盐浓度也最高, 达到0.068 mg·L-1, 叶绿素a浓度为65.46 μg·L-1, 远超其他3个湖区.竺山湾DOC浓度达到4.443 mg·L-1, 也高于其它湖区.梅梁湾硝态氮、氨态氮、亚硝态氮、磷酸盐的浓度分别为0.571、0.057、0.007和0.012 mg·L-1, DOC浓度为4.277 mg·L-1.贡湖湾水质情况也比较差, 其中硝氮, 氨氮, 亚硝氮和磷酸盐浓度分别为0.963、0.103、0.010和0.060mg·L-1, 湖心区水质相对较好, 硝氮、氨氮、亚硝氮和磷酸盐浓度分别为0.526、0.052、0.009和0.008 mg·L-1.
|
|
表 1 各个点位上覆水水质参数 Table 1 Parameters of overlying water at the sampling sites |
夏季平均水温超过28℃(表 1), 比春季要高9℃左右.总体上, 夏季不同湖区氨氮和磷酸盐浓度要高于春季, 分别为0.076~0.352 mg·L-1和0.006~0.056 mg·L-1.而硝氮的浓度远低于春季(0.003~1.069mg·L-1, 表 1).
2.2 不同湖区反硝化与厌氧氨氧化脱氮的相对贡献率流动培养结果显示(图 2), 春季竺山湾的脱氮能力最强, 反硝化速率为(142.43±35.54)μmol·(m2·h)-1, 厌氧氨氧化速率为(17.95±8.66)μmol·(m2·h)-1, 反硝化对脱氮的贡献率在4个湖区中最低, 厌氧氨氧化对脱氮的贡献率在4个湖区中最高, 分别为(88.80±4.62)%和(11.20±1.53)%; 贡湖湾次之, 反硝化速率为(72.78±11.83)μmol·(m2·h)-1, 厌氧氨氧化速率为(7.65±3.65)μmol·(m2·h)-1, 反硝化和厌氧氨氧化对脱氮的贡献率分别为(90.48±5.56)%和(9.52±1.52)%; 湖心区反硝化速率为(50.79±9.24)μmol·(m2·h)-1, 厌氧氨氧化速率为(4.96±2.62)μmol·(m2·h)-1, 反硝化和厌氧氨氧化对脱氮的贡献率分别为(91.10±5.64)%和(8.90±1.62)%; 梅梁湾反硝化速率最低, 为(27.74±8.45)μmol·(m2·h)-1, 厌氧氨氧化速率为(2.35±1.06)μmol·(m2·h)-1, 反硝化对脱氮的贡献率最高, 厌氧氨氧化对脱氮的贡献率最低, 分别为(92.18±4.69)%和(7.82±1.71)%.

|
图 2 春季和夏季不同湖区脱氮速率与脱氮贡献率 Fig. 2 Nitrogen removal rates and contribution rates of different lake areas in spring and summer |
夏季竺山湾的脱氮能力最强, 反硝化速率为(165.68±62.14)μmol·(m2·h)-1, 厌氧氨氧化速率为(33.56±10.66)μmol·(m2·h)-1, 反硝化对脱氮的贡献率在所有湖区中最低, 厌氧氨氧化对脱氮的贡献率在所有湖区中最高, 分别为(83.15±5.54)%和(16.85±1.78)%; 其它5个采样湖区的脱氮速率相差不大, 反硝化速率为(25.47±10.46)~(42.50±16.46)μmol·(m2·h)-1, 厌氧氨氧化速率为(2.65±0.94)~(5.95±2.65)μmol·(m2·h)-1, 反硝化和厌氧氨氧化对脱氮的贡献率分别为(86.38±5.65)%~(92.76±5.57)%和(13.62±1.95)%~(7.24±1.78)%.
2.3 反硝化和厌氧氨氧化速率与环境因子的相关性反硝化与厌氧氨氧化速率与无机营养盐和溶解氧都呈现出了显著的相关性, 其中, 反硝化速率与硝氮和亚硝氮浓度相关性最好, 厌氧氨氧化与亚硝氮和氨氮相关性最好.温度、pH和叶绿素a的浓度与反硝化和厌氧氨氧化速率无明显相关性(表 2).
|
|
表 2 反硝化和厌氧氨氧化速率与环境因子的相关性1) Table 2 Correlation between the denitrification and ANAMMOX rates and environmental factors |
3 讨论 3.1 氮浓度是限制春季和夏季反硝化和厌氧氨氧化速率的主要因素
相关性分析显示, 反硝化速率与硝氮和亚硝氮浓度的相关性最好(P < 0.01, 表 2), 说明底物的浓度对反硝化速率的影响最大.在污染严重的竺山湾地区, 硝酸盐的浓度要远高于其他几个采样湖区, 所以反硝化速率也最高.在春季, 不同湖区硝氮的浓度差异比较大, 反硝化速率也呈现了非常显著的差异, 而在夏季, 在采样的6个湖区中, 除了竺山湾的硝氮浓度很高, 其他5个湖区的硝氮浓度都处在一个很低的水平, 所以竺山湾的反硝化速率很高, 而其他几个湖区的反硝化速率很低, 而且差异不很明显.此外还发现, 尽管夏季的温度比较高, 但是除了竺山湾外, 其他几个湖区夏季的反硝化速率明显要比春季低, 进一步说明夏季反硝化速率主要受氮浓度限制.溶解氧也是影响反硝化速率的重要因素(P < 0.05).大多数的反硝化菌是厌氧菌, 它们对氧气很敏感, 所以低溶解氧能促进反硝化菌的生长[23].
厌氧氨氧化速率的空间变化与反硝化速率的变化比较同步, 反硝化速率高的点位厌氧氨氧化速率也高.统计分析结果显示, 厌氧氨氧化速率明显也是受底物氨氮, 硝氮和亚硝氮的浓度限制(P < 0.01), 同样, 较低的溶解氧也有利于厌氧氨氧化反应的进行(P < 0.05)[24], 而且厌氧氨氧化速率与溶解氧的相关性显著高于反硝化速率与溶解氧的相关性, 说明厌氧氨氧化相比于反硝化对低氧环境的依赖性更强.但是与反硝化速率相比, 厌氧氨氧化速率明显要低很多.这是因为厌氧氨氧化菌是自养型厌氧菌, 而太湖作为一个平均水深只有1.9 m的浅水湖泊, 并没有明显的氧跃层, 而且沉积物与水界面极易受到外界的干扰, 比如溶氧的侵蚀, 外源有机污染的输入, 这些都能显著抑制厌氧氨氧化菌的活性[24].
有趣的是, 反硝化速率与氨氮浓度, 厌氧氨氧化速率与硝氮浓度都呈现出了显著的正相关(P < 0.05, 表 2), 这似乎不合常理.实际上, 在自然水环境中, 反硝化和硝化反应往往是同时发生的, 而硝氮除了河道入水和降雨等物理来源之外, 只有氨氮硝化这一条生化途径[25], 高浓度氨氮的硝化过程为反硝化提供了充足的硝氮和亚硝氮底物.厌氧氨氧化与硝氮呈现显著正相关是因为氨氮耦合了硝氮反硝化生成的亚硝氮而实现了厌氧氨氧化过程.此外也有研究表明硝氮能作为电子受体代替亚硝氮参与厌氧氨氧化过程[26].
3.2 蓝藻堆积有利于促进脱氮过程除了温度升高的原因外[27], 蓝藻水华也很可能是促进湖泊脱氮潜力, 导致夏季水体的脱氮速率受到氮限制的另一个重要原因.有研究表明, 蓝藻水华使水库沉积物表面的厌氧氨氧化菌的丰度高了9~25倍且厌氧氨氧化菌丰度的改变与氮(如氨氮)和有机碳有关[28].另外有多篇文献报道蓝藻能促进反硝化的发生[13, 29].蓝藻可能是通过改变水体溶解氧, 或者为反硝化和厌氧氨氧化提供反应的底物而促进脱氮过程的.蓝藻的堆积会使下层的藻无法接触阳光而转为有氧呼吸, 当水柱中蓝藻的呼吸作用大于光合作用时, 水体的溶解氧就会降低, 蓝藻降解后会为水体提供大量的氨氮和有机碳, 保证了厌氧氨氧化和反硝化的发生.同时有研究报导蓝藻能为反硝化菌提供附着的载体, 有大量蓝藻颗粒的水柱反硝化潜力很强[30].
此外, 本研究发现反硝化和厌氧氨氧化速率都与磷浓度显著相关(P < 0.01).磷和氮是影响水体富营养化的重要因子, 磷浓度高的地方通常氮浓度也很高.而且磷虽然不直接参与反硝化和厌氧氨氧化, 但作为限制水体初级生产力的因子[31], 可能会间接影响到脱氮过程.比如通过引起蓝藻水华来加速湖泊脱氮过程.
3.3 蓝藻水华促进厌氧氨氧化对脱氮的贡献率厌氧氨氧化对脱氮的贡献率并不高(图 2), 这说明反硝化作用是太湖脱氮的主要形式.夏季厌氧氨氧化对脱氮的贡献率明显比春季要大, 而且水华严重的竺山湾厌氧氨氧化对脱氮的贡献率也明显高于其他几个采样湖区.统计分析表明, 厌氧氨氧化对脱氮的贡献率与叶绿素a的浓度显著相关(P < 0.05)这说明蓝藻水华对厌氧氨氧化脱氮贡献率的变化有很大的影响.Lansdown等[32]的研究发现尽管厌氧氨氧化需要缺氧条件, 但是其可以通过耦合硝化反应产生的亚硝氮而达到一个可观的脱氮速率.本研究发现夏季反硝化速率普遍低于春季, 但是厌氧氨氧化速率相比于春季并没有明显地下降.这是因为尽管夏季硝氮的含量普遍低于春季, 但是氨氮和亚硝氮相比于春季却没有明显地下降, 甚至有所升高, 而且夏季水体的溶解氧并不低, 这说明水柱中可能存在较强的硝化作用, 而硝化作用为厌氧氨氧化提供了底物亚硝氮, 硝化作用耦合厌氧氨氧化作用的发生可能是夏季厌氧氨氧化速率并没有明显降低的原因之一.有研究报道, 蓝藻死亡降解会导致沉积物氨氮的大量释放[33], 同时, 蓝藻自身死亡降解也会产生大量的有机氮和氨氮[34], 这说明蓝藻水华可以通过改变水体溶解氧、pH和营养盐浓度而直接或间接影响到氨氮浓度, 最后影响到厌氧氨氧化对脱氮的贡献率.
4 结论(1) 太湖氮磷浓度具有明显的季节变化, 夏季水体硝氮浓度显著降低, 而氨氮和磷酸根的浓度有所升高.
(2) 太湖沉积物反硝化速率和厌氧氨氧化速率春季呈现出明显的空间差异, 反硝化速率从高到低依次为竺山湾, 贡湖湾, 湖心区和梅梁湾, 厌氧氨氧化速率从高到低依次为竺山湾, 贡湖湾, 湖心区和梅梁湾; 夏季除了污染较重的竺山湾外, 其他湖区的反硝化和厌氧氨氧化速率无明显的空间差异.
(3) 总体来看, 太湖沉积物夏季反硝化和厌氧氨氧化速率要普遍低于春季, 主要控制因子是水体氮的浓度.
(4) 蓝藻水华严重的湖区厌氧氨氧化对脱氮的贡献率明显高于其他湖区.
| [1] | Zhang X K, Li B L, Xu H, et al. Effect of micronutrients on algae in different regions of Taihu, a large, spatially diverse, hypereutrophic lake[J]. Water Research, 2019, 151: 500-514. DOI:10.1016/j.watres.2018.12.023 |
| [2] |
汪岩山. 蓝藻水华对浮游细菌群落结构的影响及蓝藻生物质应用研究[D]. 合肥: 中国科学技术大学, 2018. Wang Y S. Bacterioplankton community structure in response to algal blooms and applications of algal biomass[D]. Hefei: University of Science and Technology of China, 2018. |
| [3] |
王沙沙, 潘阳, 谭宇杰, 等. 厌氧氨氧化技术现状、研究进展及主要影响因素[J]. 环境卫生工程, 2020, 28(1): 64-69, 73. Wang S S, Pan Y, Tan Y J, et al. Status, research progress and main influence factors of anammox technology[J]. Environmental Sanitation Engineering, 2020, 28(1): 64-69, 73. |
| [4] | Song B, Buckner C T, Hembury D J, et al. Impact of volcanic ash on anammox communities in deep sea sediments[J]. Environmental Microbiology Reports, 2014, 6(2): 159-166. DOI:10.1111/1758-2229.12137 |
| [5] | Reisinger A J, Tank J L, Hoellein T J, et al. Sediment, water column, and open-channel denitrification in rivers measured using membrane-inlet mass spectrometry[J]. Journal of Geophysical Research: Biogeosciences, 2016, 121(5): 1258-1274. DOI:10.1002/2015JG003261 |
| [6] | Lang B, Medeiros A S, Worsley A, et al. Influence of industrial activity and pollution on the paleoclimate reconstruction from a eutrophic lake in lowland England, UK[J]. Journal of Paleolimnology, 2018, 59(4): 397-410. DOI:10.1007/s10933-017-9995-6 |
| [7] | Saunders D L, Kalff J. Denitrification rates in the sediments of Lake Memphremagog, Canada-USA[J]. Water Research, 2001, 35(8): 1897-1904. DOI:10.1016/S0043-1354(00)00479-6 |
| [8] | Roland F A E, Darchambeau F, Borges A V, et al. Denitrification, anaerobic ammonium oxidation, and dissimilatory nitrate reduction to ammonium in an East African Great Lake (Lake Kivu)[J]. Limnology and Oceanography, 2018, 63(2): 687-701. DOI:10.1002/lno.10660 |
| [9] | Xie H T, Ji D D, Zang L H. Effects of inhibition conditions on anammox process[J]. IOP Conference Series: Earth and Environmental Science, 2017, 100. DOI:10.1088/1755-1315/100/1/012149 |
| [10] | 秦伯强, 胡维平, 陈伟民. 太湖水环境演化过程与机理[M]. 北京: 科学出版社, 2004. |
| [11] |
陆沈钧, 姚俊, 曹翔. 浅析太湖流域农业面源污染现状、成因及对策[J]. 水利发展研究, 2020, 20(2): 40-44, 53. Lu S J, Yao J, Cao X. Status, causes and Countermeasures of agricultural non-point source pollution in Taihu Lake Basin[J]. Water Resources Development Research, 2020, 20(2): 40-44, 53. |
| [12] |
朱昕阳, 蒋彩萍, 马晓雁, 等. 太湖流域浙江片区出入境水量、水质及污染物通量[J]. 湖泊科学, 2020, 32(3): 629-640. Zhu X Y, Jiang C P, Ma X Y, et al. Water quantity, quality and pollutant flux of inbound and outbound water in Zhejiang area of Taihu Basin[J]. Journal of Lake Sciences, 2020, 32(3): 629-640. |
| [13] |
刘志迎, 许海, 詹旭, 等. 蓝藻水华对太湖水柱反硝化作用的影响[J]. 环境科学, 2019, 40(3): 1261-1269. Liu Z Y, Xu H, Zhan X, et al. Influence of cyanobacterial blooms on denitrification rate in shallow Lake Taihu, China[J]. Environmental Science, 2019, 40(3): 1261-1269. DOI:10.3969/j.issn.1000-6923.2019.03.044 |
| [14] |
徐徽, 张路, 商景阁, 等. 太湖梅梁湾水土界面反硝化和厌氧氨氧化[J]. 湖泊科学, 2009, 21(6): 775-781. Xu H, Zhang L, Shang J G, et al. Denitrification and anammox on the sediment-water interface in the Meiliang Bay of Lake Taihu[J]. Journal of Lake Sciences, 2009, 21(6): 775-781. |
| [15] | Yao Z B, Wang F, Wang C L, et al. Anaerobic ammonium oxidation coupled to ferric iron reduction in the sediment of a eutrophic lake[J]. Environmental Science and Pollution Research, 2019, 26(15): 15084-15094. DOI:10.1007/s11356-019-04907-7 |
| [16] |
陈志浩, 覃云斌, 丁帮璟, 等. 宛山荡农田土壤氮迁移过程反硝化与厌氧氨氧化[J]. 环境科学, 2020, 41(1): 412-419. Chen Z H, Qin Y B, Ding B J, et al. Denitrification and anaerobic ammonium oxidation in soil nitrogen migration process in a farmland of Wanshandang Lake[J]. Environmental Science, 2020, 41(1): 412-419. |
| [17] | Gao H, Schreiber F, Collins G, et al. Aerobic denitrification in permeable Wadden Sea sediments[J]. The ISME Journal, 2010, 4(3): 417-426. DOI:10.1038/ismej.2009.127 |
| [18] | Rysgaard S, Risgaard-Petersen N, Nielsen L P, et al. Nitrification and denitrification in lake and estuarine sediments measured by the15N dilution technique and isotope pairing[J]. Applied and Environmental Microbiology, 1993, 59(7): 2093-2098. DOI:10.1128/AEM.59.7.2093-2098.1993 |
| [19] |
陈明, 孙洁梅, 李敏, 等. 水体中藻类叶绿素a提取方法的比较[J]. 环境监测管理与技术, 2017, 29(4): 57-59. Chen M, Sun J M, Li M, et al. Comparison for extracting chlorophyll-a from algae in water[J]. The Administration and Technique of Environmental Monitoring, 2017, 29(4): 57-59. DOI:10.3969/j.issn.1006-2009.2017.04.014 |
| [20] |
李进芳, 柴延超, 陈顺涛, 等. 利用膜进样质谱仪测定水稻土几种厌氧氮转化速率[J]. 农业环境科学学报, 2019, 38(7): 1541-1549. Li J F, Chai Y C, Chen S T, et al. Measurement of denitrification, Anammox, DNRA rates, and net N2 flux in paddy soil using a membrane inlet mass spectrometer[J]. Journal of Agro-Environment Science, 2019, 38(7): 1541-1549. |
| [21] | Hou L J, Yin G Y, Liu M, et al. Effects of sulfamethazine on denitrification and the associated N2O release in estuarine and coastal sediments[J]. Environmental Science & Technology, 2015, 49(1): 326-333. |
| [22] | Thamdrup B, Dalsgaard T. Production of N2 through anaerobic ammonium oxidation coupled to nitrate reduction in marine sediments[J]. Applied and Environmental Microbiology, 2002, 68(3): 1312-1318. DOI:10.1128/AEM.68.3.1312-1318.2002 |
| [23] |
王海燕, 赵远哲, 王文富, 等. 人工湿地脱氮影响因素及强化措施研究进展[J]. 环境工程技术学报, 2020, 10(4): 585-597. Wang H Y, Zhao Y Z, Wang W F, et al. A review of influencing factors and enhanced measures for nitrogen removal of constructed wetlands[J]. Journal of Environmental Engineering Technology, 2020, 10(4): 585-597. |
| [24] | 冯宁, 张文文, 马晨曦. 厌氧氨氧化反应影响因素研究进展[A]. 见: 2018中国环境科学学会科学技术年会论文集(第一卷)[C]. 合肥: 中国环境科学学会, 2018. |
| [25] | Kuypers M M M, Marchant H K, Kartal B. The microbial nitrogen-cycling network[J]. Nature Reviews Microbiology, 2018, 16(5): 263-276. DOI:10.1038/nrmicro.2018.9 |
| [26] | Waki M, Yasuda T, Fukumoto Y, et al. Effect of electron donors on anammox coupling with nitrate reduction for removing nitrogen from nitrate and ammonium[J]. Bioresource Technology, 2013, 130: 592-598. DOI:10.1016/j.biortech.2012.12.101 |
| [27] |
孙小溪, 蒋宏忱. 湖泊微生物反硝化过程及速率研究进展[J]. 微生物学报, 2020, 60(6): 1162-1176. Sun X X, Jiang H C. Progress in microbially mediated denitrifying in lacustrine environments[J]. Acta Microbiologica Sinica, 2020, 60(6): 1162-1176. |
| [28] | Xue Y Y, Yu Z, Chen H H, et al. Cyanobacterial bloom significantly boosts hypolimnelic anammox bacterial abundance in a subtropical stratified reservoir[J]. FEMS Microbiology Ecology, 2017, 93(10). DOI:10.1093/femsec/fix118 |
| [29] |
肖琳, 杨佳怡. 水华对沉积物中氮的转化和去除的影响[J]. 环境科学, 2019, 40(9): 4018-4022. Xiao L, Yang J Y. Water bloom modified sediment nitrogen transformation and removal[J]. Environmental Science, 2019, 40(9): 4018-4022. |
| [30] | Chen X F, Jiang H Y, Sun X, et al. Nitrification and denitrification by algae-attached and free-living microorganisms during a cyanobacterial bloom in Lake Taihu, a shallow Eutrophic Lake in China[J]. Biogeochemistry, 2016, 131(1-2): 135-146. DOI:10.1007/s10533-016-0271-z |
| [31] | Alvarez-Clare S, Mack M C, Brooks M. A direct test of nitrogen and phosphorus limitation to net primary productivity in a lowland tropical wet forest[J]. Ecology, 2013, 94(7): 1540-1551. |
| [32] | Lansdown K, McKew B A, Whitby C, et al. Importance and controls of anaerobic ammonium oxidation influenced by riverbed geology[J]. Nature Geoscience, 2016, 9(5): 357-360. |
| [33] |
朱梦圆, 朱广伟, 王永平. 太湖蓝藻水华衰亡对沉积物氮、磷释放的影响[J]. 环境科学, 2011, 32(2): 409-415. Zhu M Y, Zhu G W, Wang Y P. Influence of scum of algal bloom on the release of N and P from sediments of Lake Taihu[J]. Environmental Science, 2011, 32(2): 409-415. |
| [34] |
余岑涔, 马杰, 许晓光, 等. 太湖近岸带草藻残体分解对水质的影响[J]. 农业环境科学学报, 2018, 37(2): 302-308. Yu C C, Ma J, Xu X G, et al. Influence of decomposition processes of higher aquatic plants and algae on water quality near the shoreline of Lake Taihu[J]. Journal of Agro-environment Science, 2018, 37(2): 302-308. |