 2021, Vol. 42
2021, Vol. 42

2. 陕西省水工环地质调查中心, 西安 710068;
3. 农业部西北植物营养与农业环境重点实验室, 杨凌 712100
 , QI Ming-xing1 , WANG Min1 , LIU Na-na1 , Pornpimol Kleawsampanjai1 , ZHOU Fei1 , ZHAI Hui1 , WANG Meng-ke1 , REN Rui2 , LIANG Dong-li1,3
, QI Ming-xing1 , WANG Min1 , LIU Na-na1 , Pornpimol Kleawsampanjai1 , ZHOU Fei1 , ZHAI Hui1 , WANG Meng-ke1 , REN Rui2 , LIANG Dong-li1,3
2. Shaanxi Hydrogeological Engneering Geology and Environment Geology Survey Center, Xi'an 710068, China;
3. Key Laboratory of Plant Nutrition and the Agi-environment in Northwest China, Ministry of Agriculture, Yangling 712100, China
镉是环境中毒性较强的重金属元素之一, 具有易移动和难降解等特点[1].据统计, 我国农田镉污染土壤面积超过28万hm2, 镉含量超标的农产品每年高达150万t[2, 3].镉被植物根系吸收后会破坏植物体内的营养平衡, 降低与叶绿素合成相关的酶的活性, 破坏叶绿体的结构, 使叶绿素含量降低[4, 5], 进而导致光合速率下降, 最终抑制植物生长[6].此外, 镉还会降低植物对水分的吸收, 导致气孔导度降低, CO2的固定过程受阻, 净光合速率也随之下降, 进而导致作物产量与品质的下降[7, 8].而农产品中镉的残留和富集则会通过食物链危及人体健康[9].因此, 土壤镉污染防治是目前亟待解决的重要的土壤环境问题之一.
硒是人体和动物必需的微量营养元素[10, 11].适量的硒可以通过调节活性氧代谢, 从而减轻重金属胁迫对植物产生的氧化应激反应[12, 13].硒还可以通过调节光合系统和修复受损细胞来减轻重金属的毒性[14].Sun等[15]的研究发现, 施入硒能够减缓镉对黄瓜光合作用的抑制, 提高植物的净光合速率.Zhang等[16]的研究却发现, 当施硒量>50 g·hm-2反而降低了镉污染土壤中水稻的净光合速率、蒸腾速率和胞间CO2浓度.由此可见, 外源硒对植物镉胁迫下光合作用的影响不尽相同, 有必要进一步查明镉胁迫下植物对不同含量外源硒的响应.
土壤中的亚硒酸盐和硒酸盐是植物吸收硒的主要形式[17].且适量的两种硒均能降低植物对镉的积累, 但现有的研究结果多为单一施入硒酸盐或亚硒酸盐[7, 18], 对不同种类外源硒对植物镉的解毒作用尚未见系统报道.小白菜常被用作重金属污染相关研究的指示作物, 且作为十字花科植物具有较强的富硒能力, 也常被用于富硒蔬菜的研究[19].因此, 本文采用盆栽试验, 以天然镉污染土壤为供试土壤, 小白菜为研究对象, 探究不同外源硒对镉胁迫下小白菜生长和光合作用及对镉吸收和转运的影响, 通过寻求适宜的缓解镉毒害的外源硒种类及施硒量, 以期为土壤镉污染控制与农作物安全生产提供科学依据.
1 材料与方法 1.1 试验材料供试土壤为西安某区的镉污染土壤(0~20 cm).土壤自然风干, 过5 mm筛供盆栽试验使用, 并过0.1、0.25和1 mm筛用于土壤理化性质的测定.测定结果为:pH 8.14; 阳离子交换量13.88 cmol·kg-1; 碳酸钙27.99 g·kg-1; 电导率115 dS·m-1; 有机质15.83 g·kg-1; 无定型铁0.92 g·kg-1; 游离态铁0.92 g·kg-1.土壤总镉为5.36 mg·kg-1, 总硒为0.37 mg·kg-1.
小白菜种子由西北农林科技大学种子公司提供.外源硒为亚硒酸钠(Na2SeO3)和硒酸钠(Na2SeO4), 均购自天津化学试剂厂.
1.2 试验设计本研究采用盆栽试验, 将供试土壤装入塑料花盆(直径23 cm、高16.5 cm, 每盆装土5 kg), 然后将亚硒酸钠或硒酸钠以溶液的形式均匀喷入土壤中.两种外源硒的施用量为0、0.5、1和2.5 mg·kg-1[20], 共7个处理, 每个处理重复3次, 共21盆.外源硒施入土壤后平衡21 d[21], 期间保持土壤水分含量为田间持水量的70%.
平衡结束后, 每盆施入氮肥(150 mg·kg-1)和磷肥(80 mg·kg-1), 并浇水至100%田间持水量后播种, 种植前小白菜种子用0.5%次氯酸钠溶液浸泡10 min后用去离子水清洗3次.每盆播种15粒.作物出苗7 d后间苗, 10 d后定苗5株.小白菜生长期间, 保持土壤湿度为田间持水量的70%, 并于播种后的第35 d收获.
1.3 测定指标及方法 1.3.1 小白菜根长、株高和生物量收获后的小白菜用自来水、蒸馏水洗涤3次, 吸水纸吸干水分, 分为地上部和地下部.并测量其根长和株高.后将其于95℃下杀青30 min, 50℃烘至恒重并称重, 烘干的样品用球磨机研磨后密封储存备用.
1.3.2 小白菜SPAD值及光合指标作物收获前, 于08:30~11:30使用SPAD-502叶绿素仪测定叶绿素(SPAD), 用便携式光合速率测定仪(LI-6400XT, 美国)测定同一叶位(第三片展开叶)叶片的光合参数, 包括:净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)和蒸腾速率(Tr).
1.3.3 小白菜镉含量将烘干后的样品用4∶1(体积比)的HNO3-HClO4混酸进行消解, 待消解完全后定容并过滤, 使用电感耦合等离子体质谱仪(ICP-MS, 美国)测定上清液镉含量.
1.3.4 土壤理化性质及镉和硒含量土壤基本理化性质根据鲍士旦[22]的方法测定, 土壤总镉测定根据GB/T 17141-1997测定, 土壤总硒含量根据Peng等[23]的方法测定.
1.4 质量控制为保证测定结果的准确性, 在测定植物镉含量时, 每批次试验均分别设置3个空白及重复.测定过程中使用小麦(GBW10011)作为质量控制样品, 镉标准值为(18±4) μg·kg-1, 实测值为(20±3) μg·kg-1.
1.5 数据统计分析使用转运系数(TF)反映小白菜根部吸收镉后转运到地上部的能力, 计算公式(1)如下:
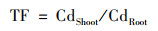
|
(1) |
式中, CdRoot和CdShoot分别指小白菜地下部和地上部镉含量(mg·kg-1).
采用Excel 2013进行数据处理及图表绘制, SPSS 22.0进行统计分析, 单因素方差分析(one-way ANOVA)探究硒对各指标影响的差异显著性.
2 结果与分析 2.1 不同外源硒对镉污染土壤小白菜生长的影响 2.1.1 对根长和株高的影响如图 1所示, 与对照相比, 外源施硒对小白菜的根长均无显著影响(P>0.05), 但低硒处理(≤1.0 mg·kg-1)却不同程度地增加了小白菜的株高.且施硒量为1.0 mg·kg-1时的促进作用最为显著, 亚硒酸盐和硒酸盐处理分别较对照提高了18.3%和12.7%, 而高硒处理(2.5 mg·kg-1)则对其无显著影响(P>0.05).由此可见, 镉胁迫下低硒处理促进了小白菜地上部的生长, 且亚硒酸盐的促进作用大于硒酸盐.

|
图例表示不同的施硒量(mg·kg-1), 下同 图 1 镉污染土壤施硒对小白菜根长和株高的影响 Fig. 1 Effects of Se application on root length and shoot height of pak choi in Cd contaminated soil |
施硒对小白菜生物量的影响如图 2所示.与对照相比, 除1 mg·kg-1亚硒酸盐处理使小白菜地下部生物量显著提高了13%(P < 0.05), 其他处理对地下部生物量均无显著影响(P>0.05).对小白菜地上部生物量而言, 除2.5 mg·kg-1的硒酸盐处理对其无显著影响外(P>0.05), 其他处理的小白菜地上部生物量均有不同程度地提高, 且以1 mg·kg-1的亚硒酸盐处理促进作用最显著(P < 0.05).此外, 当施硒量相同时, 硒酸盐处理的小白菜地上部和地下部生物量低于相同的亚硒酸盐处理.

|
图 2 镉污染土壤施硒对小白菜生物量的影响 Fig. 2 Effects of Se application on biomass of pak choi in Cd contaminated soil |
施硒对小白菜光合作用的影响如表 1所示.与对照相比, 无论亚硒酸盐处理还是硒酸盐处理, 小白菜的SPAD值随施硒量的增加均呈先增加后逐渐下降的规律.此外, 低硒处理较对照显著提高了小白菜的SPAD值, 且亚硒酸盐处理对SPAD值增加(30.4%~32.9%, P < 0.05)显著高于硒酸盐处理(7.7%~15.4%, P < 0.05).但高硒处理对小白菜的SPAD值均无显著影响(P>0.05).随着亚硒酸盐和硒酸盐施用量的增加, 小白菜的净光合速率、气孔导度和胞间CO2浓度均呈先增加后降低的趋势, 且施硒量为1.0 mg·kg-1时提高效果最为显著(胞间CO2浓度除外, P < 0.05).亚硒酸盐处理小白菜叶片的净光合速率、气孔导度和胞间CO2浓度与对照相比分别提高了39.2%、121%和17.8%, 而硒酸盐处理分别提高了25.2%、67.5%和22.3%.此外, 叶片的蒸腾速率随亚硒酸盐施用量的升高而增加, 且在施硒量为2.5 mg·kg-1时达到最高(与未施硒相比升高了31.8%); 与亚硒酸盐处理相反, 硒酸盐处理小白菜叶片的蒸腾速率随施硒量的升高逐渐下降.
|
|
表 1 镉污染土壤施硒对小白菜SPAD值及叶片光合指标的影响1) Table 1 Effects of Se application on SPAD value and leaf photosynthetic parameters of pak choi in Cd contaminated soil |
2.3 外源硒对镉污染土壤中小白菜镉吸收、转运的影响
施硒对小白菜地上、地下镉含量的影响如图 3所示, 小白菜地上部镉含量高于地下部.与对照相比, 施加亚硒酸盐显著降低了小白菜地下部和地上部的镉含量(P < 0.05).其中, 小白菜地下部镉含量随亚硒酸盐施用量的升高呈显著下降趋势(P < 0.05), 并在亚硒酸盐为2.5 mg·kg-1时最低, 较对照处理下降了57%.而小白菜地上部的镉含量随亚硒酸盐施入水平呈先降低, 在1.0 mg·kg-1时达到最低而后升高的趋势.与此不同, 硒酸盐处理小白菜地上部镉含量在0.5 mg·kg-1处理时达最低, 而后随硒酸盐施用量的增加地上部镉含量呈上升趋势.由以上可知, 在镉胁迫下施用适量的亚硒酸盐和硒酸盐均可以降低小白菜对镉的吸收, 且1.0 mg·kg-1的亚硒酸盐更能降低镉在小白菜地上部的积累.

|
图 3 镉污染土壤施硒对小白菜镉吸收及转运的影响 Fig. 3 Effects of Se application on the uptake and translocation of pak choi in Cd contaminated soil |
TF值用来评价植物将镉离子从地下部向地上部转移和富集的能力.TF值越大, 转运能力越强.由图 3(c)可知, 对于亚硒酸盐, 低硒处理对小白菜的TF值无显著影响(P>0.05), 但高硒处理显著提高了小白菜的TF值.与此不同, 施用0.5 mg·kg-1的硒酸盐显著降低了小白菜的TF值, 而施用1.0和2.5 mg·kg-1的硒酸盐却显著提高了小白菜的TF值(P < 0.05).由此可见, 亚硒酸盐和硒酸盐对小白菜转运镉的影响存在差异.
3 讨论 3.1 施硒对镉污染土壤中小白菜镉的解毒作用本研究发现, 外源施硒显著降低了小白菜根系对镉的吸收[图 3(a)], 且地下部镉含量与施硒量呈极显著负相关(P < 0.01, 表 2), 表明外源硒与土壤镉的相互作用降低了植物根系对镉的吸收.Zhang等[24]通过对玉米的研究也认为, 土壤中的硒和镉能够形成难以被植物吸收的Cd-Se复合物, 进而抑制植物对镉的吸收.本研究还发现, 施入0.5 mg·kg-1硒酸盐显著降低了小白菜镉的TF值[图 3(c)], 说明适量的外源硒可以通过抑制镉向地上部转运进而降低镉在可食部位的积累.Wan等[25]在碱性土壤中施入外源硒后发现镉在水稻地上部的积累降低.这是由于镉被根系吸收后, 通过质外体途径进入维管柱并向地上部运输[26], 而硒可以促进水稻根系内皮凯氏带的形成, 阻断皮层与维管柱之间的质外体运输途径, 进而减少了镉进入维管柱[25].
|
|
表 2 小白菜的生长生理指标与施硒量和镉含量间的相关关系1) Table 2 Correlation between the growth physiological indices and selenium application rate and cadmium concentration of pak choi |
生物量的变化通常是衡量植物对外界胁迫的耐性或者抗性的重要指标之一[27].适量外源硒能缓解重金属对植物的毒害作用, 提高重金属胁迫下作物的生物量[28, 29].本研究发现, 无论亚硒酸盐还是硒酸盐, 低硒处理不同程度地提高了小白菜地上部生物量和株高(图 1和图 2), 与Wan等[30]的研究结果相一致, 低硒处理小白菜地上部镉含量较对照显著下降了18%~40%[图 3(b)].主要原因有以下两点:一方面, 适量的硒可以降低植物体内活性氧的积累, 改善营养平衡从而促进植物的生长[12]; 另一方面, 硒可以通过降低小白菜对镉的吸收, 减轻镉产生的氧化胁迫, 使镉胁迫导致的小白菜叶片的净光合速率、气孔导度、胞间CO2浓度的下降得到缓解(表 1), 并促进小白菜的生长.
植物的光合作用对重金属非常敏感, 低含量的镉就能抑制水稻、玉米等植物的光合作用, 使植物的生长受阻, 生物量降低[31, 32]. Cd2+可以通过Ca2+、Mg2+、Zn2+等离子的通道进入细胞或阻滞细胞通道, 同时干扰其各项功能.此外, 镉还能抑制原叶绿素脂还原酶的活性, 干扰叶片中叶绿素的合成[33].本研究中, 亚硒酸盐和硒酸盐施用量≤1.0 mg·kg-1时分别使小白菜的SPAD值提高了30.4%~32.9%和7.7%~15.5%(表 1).这可能是由于外源施硒后小白菜地上部的镉含量下降, 减少了Cd2+对Ca2+、Mg2+和Zn2+等离子的替代作用[34, 35].此外, 本研究发现亚硒酸盐或硒酸盐≤1.0 mg·kg-1时能不同程度提高植物的净光合速率、气孔导度和胞间CO2浓度(表 1), 且小白菜的净光合速率、气孔导度和胞间CO2浓度与叶片的镉含量呈负相关(表 2), 这与Gao等[7]的研究结果一致, 即镉可以抑制植物的光合作用, 而施硒后这种抑制作用得到缓解.究其原因主要有以下两点:第一, 适量的硒可以防止叶绿素降解并在一定程度上抵消镉的破坏效应[6].本研究中发现适量的硒能够提高小白菜叶片硒含量(数据未给出)及SPAD值(表 1), 且小白菜的SPAD值与净光合速率、气孔导度和胞间CO2浓度呈显著正相关关系(P < 0.05, 表 2), 也进一步证明了硒可以通过提高叶绿素的含量而促进光合作用.第二, 硒能诱导光合作用相关蛋白的积累, 这些蛋白可以介导电子转移, 作为光受体或在叶绿体中的蛋白质生物合成中起作用, 从而使镉胁迫下光合作用得到改善[15], 本研究中低硒处理小白菜叶片的净光合速率、气孔导度和胞间CO2浓度均显著高于对照(P < 0.05, 表 1)也印证了这一点.
3.2 不同外源硒对镉的解毒作用的差异虽然外源硒能够提高小白菜产量, 并降低其地上部镉含量, 但本研究中高硒处理的地上部的生物量和株高均低于低硒处理.这是由于过量的硒非特异性地取代组织蛋白中的硫并形成硒代蛋氨酸和硒代半胱氨酸, 干扰机体内正常的蛋白功能从而对植物造成毒性, 并产生氧化应激作用造成的[36, 37].此外, 过量的硒会损伤光合细胞器, 与土壤中镉产生协同毒害作用[28], 对小白菜的光合作用产生抑制(表 1), 进而导致小白菜生物量下降.Chang等[38]的研究发现, 施用8.0 mg·kg-1的亚硒酸盐对水稻各部位镉含量降低程度显著大于1.0 mg·kg-1.而郭锋等[39]的研究发现, 在土壤镉含量为0.5 mg·kg-1时, 添加< 2.0 mg·kg-1的亚硒酸盐后菠菜叶片的镉含量随施硒量的增加而降低, 而当施硒量>2.0 mg·kg-1时菠菜叶片镉含量却升高.因此, 在一定的施硒范围内, 硒镉之间存在拮抗作用, 且这个施硒范围因土壤条件、植物品种不同而异[40].
在本研究中, 高硒处理的亚硒酸盐和硒酸盐处理较对照处理小白菜镉的TF值显著增加[图 3(c)].由于植物体内存在积累和区隔化机制可以将根系吸收的重金属多数转运到地上部分然后对其进行解毒作用[41].对于亚硒酸盐, 为维持植物体内代谢物质的平衡, 迫使镉向地上部转移分配促进植物本身的积累和区隔化机制进行解毒作用[42].而硒酸盐本身就具有较强的向地上部分转运的能力[20], 因此可能带动镉向地上部分转运, 但该机制还需进一步试验.
值得注意的是, 当施硒量为1.0 mg·kg-1时, 亚硒酸盐处理对小白菜生长、净光合速率的改善效果均优于相同处理的硒酸盐(图 1、图 2和表 1), 且亚硒酸盐对小白菜镉吸收的抑制作用大于硒酸盐(图 3), 造成这一差异的原因有二:一是硒酸盐在土壤中移动性较大, 而亚硒酸盐则更容易被土壤中的碳酸盐、铁锰氧化物、有机质等吸附[23], 从而使亚硒酸盐更多地残留在土壤中, 更容易与镉形成难以被植物吸收的Cd-Se复合物.二是亚硒酸盐和硒酸盐进入植物细胞后, 硒酸盐的形态基本不发生改变, 而亚硒酸盐则在根部被快速转化为硒代蛋氨酸、硒代半胱氨酸等有机硒[43].镉进一步取代有机硒中的硒, 形成镉的有机化合物, 从而降低镉的毒性[9].因此, 在利用外源硒缓解镉对植物的毒害作用时要选取适宜的施用量和添加形态.
4 结论(1) 低硒处理(≤1.0 mg·kg-1)能缓解镉对小白菜光合作用的胁迫, 促进镉污染土壤中小白菜的生长, 提高生物量.
(2) 施硒可以降低小白菜各部位的镉含量, 小白菜地上部的镉随施硒量的增加呈先降低后升高趋势, 地下部则呈降低趋势, 且亚硒酸盐的作用更强.
(3) 推荐施用1.0 mg·kg-1的亚硒酸盐来降低镉污染地区小白菜镉吸收或缓解镉对小白菜的毒性.
| [1] | Treesubsuntorn C, Dhurakit P, Khaksar G, et al. Effect of microorganisms on reducing cadmium uptake and toxicity in rice (Oryza sativa L.)[[J]. Environmental Science and Pollution Research, 2018, 25(26): 25690-25701. DOI:10.1007/s11356-017-9058-6 |
| [2] |
彭鸥, 李丹阳, 刘寿涛, 等. 镉胁迫对水稻生长发育的影响及伤流液与稻米镉含量的相关性研究[J]. 生态与农村环境学报, 2019, 35(1): 76-82. Peng O, Li D Y, Liu S T, et al. Effects of cadmium stress on rice growth and correlation between bleeding sap and cadmium content in rice[J]. Journal of Ecology and Rural Environment, 2019, 35(1): 76-82. |
| [3] |
刘霄霏, 李惠英, 陈良, 等. 外源硒对镉胁迫下黑麦草生长和生理的影响[J]. 草地学报, 2020, 28(1): 72-79. Liu X F, Li H Y, Chen L, et al. Effects of exogenous selenium on growth and physiology of ryegrass under cadmium stress[J]. Acta Agrestia Sinica, 2020, 28(1): 72-79. |
| [4] | Qian H F, Li J J, Pan X J, et al. Photoperiod and temperature influence cadmium's effects on photosynthesis-related gene transcription in Chlorella vulgaris[J]. Ecotoxicology and Environmental Safety, 2010, 73(6): 1202-1206. DOI:10.1016/j.ecoenv.2010.07.006 |
| [5] | Souza V L, De Almeida A A F, Lima S G C, et al. Morphophysiological responses and programmed cell death induced by cadmium in Genipa americana L. (Rubiaceae)[[J]. Biometals, 2011, 24(1): 59-71. DOI:10.1007/s10534-010-9374-5 |
| [6] | Liu W X, Shang S H, Feng X, et al. Modulation of exogenous selenium in cadmium-induced changes in antioxidative metabolism, cadmium uptake, and photosynthetic performance in the 2 tobacco genotypes differing in cadmium tolerance[J]. Environmental Toxicology and Chemistry, 2015, 34(1): 92-99. DOI:10.1002/etc.2760 |
| [7] | Gao M, Zhou J, Liu H L, et al. Foliar spraying with silicon and selenium reduces cadmium uptake and mitigates cadmium toxicity in rice[J]. Science of the Total Environment, 2018, 631-632: 1100-1108. DOI:10.1016/j.scitotenv.2018.03.047 |
| [8] | Wang Y, Jiang X, Li K, et al. Photosynthetic responses of Oryza sativa L. seedlings to cadmium stress: physiological, biochemical and ultrastructural analyses[J]. Biology of Metals, 2014, 27(2): 389-401. DOI:10.1007/s10534-014-9720-0 |
| [9] | Ismael M A, Elyamine A M, Moussa M G, et al. Cadmium in plants: uptake, toxicity, and its interactions with selenium fertilizers[J]. Metallomics, 2019, 11(2): 255-277. DOI:10.1039/C8MT00247A |
| [10] | Zhai H, Xue M Y, Du Z K, et al. Leaching behaviors and chemical fraction distribution of exogenous selenium in three agricultural soils through simulated rainfall[J]. Ecotoxicology and Environmental Safety, 2019, 173: 393-400. DOI:10.1016/j.ecoenv.2019.02.042 |
| [11] | Zhou F, Yang W X, Wang M K, et al. Effects of selenium application on Se content and speciation in Lentinula edodes[J]. Food Chemistry, 2018, 265: 182-188. DOI:10.1016/j.foodchem.2018.05.087 |
| [12] | Lin L, Zhou W H, Dai H X, et al. Selenium reduces cadmium uptake and mitigates cadmium toxicity in rice[J]. Journal of Hazardous Materials, 2012, 235-236: 343-351. DOI:10.1016/j.jhazmat.2012.08.012 |
| [13] | Wu Z C, Wang F H, Liu S, et al. Comparative responses to silicon and selenium in relation to cadmium uptake, compartmentation in roots, and xylem transport in flowering Chinese cabbage (Brassica campestris L. ssp. chinensis var. utilis) under cadmium stress[J]. Environmental and Experimental Botany, 2016, 131: 173-180. DOI:10.1016/j.envexpbot.2016.07.012 |
| [14] | Feng R W, Wei C Y, Tu S X. The roles of selenium in protecting plants against abiotic stresses[J]. Environmental and Experimental Botany, 2013, 87: 58-68. DOI:10.1016/j.envexpbot.2012.09.002 |
| [15] | Sun H Y, Wang X Y, Wang Y A, et al. Alleviation of cadmium toxicity in cucumber (Cucumis sativus) seedlings by the application of selenium[J]. Spanish Journal of Agricultural Research, 2016, 14(4). DOI:10.5424/sjar/2016144-10008 |
| [16] | Zhang L H, Hu B, Li W, et al. OsPT2, a phosphate transporter, is involved in the active uptake of selenite in rice[J]. New Phytologist, 2014, 201(4): 1183-1191. DOI:10.1111/nph.12596 |
| [17] | Natasha, Shahid M, Niazi N K, et al. A critical review of selenium biogeochemical behavior in soil-plant system with an inference to human health[J]. Environmental Pollution, 2018, 234: 915-934. DOI:10.1016/j.envpol.2017.12.019 |
| [18] | Sun H Y, Dai H X, Wang X Y, et al. Physiological and proteomic analysis of selenium-mediated tolerance to Cd stress in cucumber (Cucumis sativus L.)[J]. Ecotoxicology and Environmental Safety, 2016, 133: 114-126. DOI:10.1016/j.ecoenv.2016.07.003 |
| [19] |
杜振宇, 史衍玺, 王清华. 蔬菜对硒的吸收及适宜补硒食用量[J]. 生态环境, 2004, 13(2): 230-231, 267. Du Z Y, Shi Y X, Wang Q H. Selenium uptake by vegetables and recommendation of edible amounts for selenium supplement[J]. Ecology and Environment, 2004, 13(2): 230-231, 267. DOI:10.3969/j.issn.1674-5906.2004.02.026 |
| [20] |
付冬冬, 段曼莉, 梁东丽, 等. 不同价态外源硒对小白菜生长及养分吸收的影响[J]. 植物营养与肥料学报, 2011, 17(2): 358-365. Fu D D, Duan M L, Liang D L, et al. Effects of selenite and selenate on growth and nutrient absorption of pakchoi[J]. Plant Nutrition and Fertilizer Science, 2011, 17(2): 358-365. |
| [21] | Li J, Peng Q, Liang D L, et al. Effects of aging on the fraction distribution and bioavailability of selenium in three different soils[J]. Chemosphere, 2016, 144: 2351-2359. DOI:10.1016/j.chemosphere.2015.11.011 |
| [22] | 鲍士旦. 土壤农化分析[M]. 北京: 中国农业出版社, 2000. |
| [23] | Peng Q, Wang M K, Cui Z W, et al. Assessment of bioavailability of selenium in different plant-soil systems by diffusive gradients in thin-films (DGT)[J]. Environmental Pollution, 2017, 225: 637-643. DOI:10.1016/j.envpol.2017.03.036 |
| [24] | Zhang Z Z, Yuan L X, Qi S H, et al. The threshold effect between the soil bioavailable molar Se: Cd ratio and the accumulation of Cd in corn (Zea mays L.) from natural Se-Cd rich soils[J]. Science of the Total Environment, 2019, 688: 1228-1235. DOI:10.1016/j.scitotenv.2019.06.331 |
| [25] | Wan Y N, Camara A Y, Yu Y, et al. Cadmium dynamics in soil pore water and uptake by rice: influences of soil-applied selenite with different water managements[J]. Environmental Pollution, 2018, 240: 523-533. DOI:10.1016/j.envpol.2018.04.044 |
| [26] | Lux A, Martinka M, Vaculík M, et al. Root responses to cadmium in the rhizosphere: a review[J]. Journal of Experimental Botany, 2011, 62(1): 21-37. DOI:10.1093/jxb/erq281 |
| [27] | Rizwan M, Ali S, Rehman M Z U, et al. A critical review on the effects of zinc at toxic levels of cadmium in plants[J]. Environmental Science and Pollution Research, 2019, 26(7): 6279-6289. DOI:10.1007/s11356-019-04174-6 |
| [28] | Filek M, Keskinen R, Hartikainen H, et al. The protective role of selenium in rape seedlings subjected to cadmium stress[J]. Journal of Plant Physiology, 2008, 165(8): 833-844. DOI:10.1016/j.jplph.2007.06.006 |
| [29] | Huang B F, Xin J L, Dai H W, et al. Effects of interaction between cadmium (Cd) and selenium (Se) on grain yield and Cd and Se accumulation in a hybrid rice (Oryza sativa) System[J]. Journal of Agricultural and Food Chemistry, 2017, 65(43): 9537-9546. DOI:10.1021/acs.jafc.7b03316 |
| [30] | Wan Y A, Yu Y, Wang Q, et al. Cadmium uptake dynamics and translocation in rice seedling: influence of different forms of selenium[J]. Ecotoxicology and Environmental Safety, 2016, 133: 127-134. DOI:10.1016/j.ecoenv.2016.07.001 |
| [31] | Ekmekçi Y, Tanyolaç D, Ayhan B. Effects of cadmium on antioxidant enzyme and photosynthetic activities in leaves of two maize cultivars[J]. Journal of Plant Physiology, 2008, 165(6): 600-611. DOI:10.1016/j.jplph.2007.01.017 |
| [32] | Ying R R, Qiu R L, Tang Y T, et al. Cadmium tolerance of carbon assimilation enzymes and chloroplast in Zn/Cd hyperaccumulator Picris divaricata[J]. Journal of Plant Physiology, 2010, 167(2): 81-87. DOI:10.1016/j.jplph.2009.07.005 |
| [33] |
苏玲, 章永松, 林咸永, 等. 维管植物的镉毒和耐性机制[J]. 植物营养与肥料学报, 2000, 6(1): 106-112. Su L, Zhang Y S, Lin X Y, et al. Cadmium toxicity and tolerance in vascular plants[J]. Plant Nutrition and Fertilizer Science, 2000, 6(1): 106-112. DOI:10.3321/j.issn:1008-505X.2000.01.016 |
| [34] | Noctor G, Veljovic-Jovanovic S, Driscoll S, et al. Drought and oxidative load in the leaves of C3 plants: a predominant role for photorespiration?[J]. Annals of Botany, 2002, 89(7): 841-850. DOI:10.1093/aob/mcf096 |
| [35] | Sharma P, Jha A B, Dubey R S, et al. Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions[J]. Journal of Botany, 2012, 2012. DOI:10.1155/2012/217037 |
| [36] | van Hoewyk D. A tale of two toxicities: malformed selenoproteins and oxidative stress both contribute to selenium stress in plants[J]. Annals of Botany, 2013, 112(6): 965-972. DOI:10.1093/aob/mct163 |
| [37] |
覃思跃, 赵文龙, 李俊, 等. 硒与硫单一及交互作用对小白菜生长及硒生物有效性的影响[J]. 环境科学学报, 2016, 36(4): 1500-1507. Qin S Y, Zhao W L, Li J, et al. Single and combined effects of sulfate and selenate on growth and selenium bioavailability of pak choi[J]. Acta Scientiae Circumstantiae, 2016, 36(4): 1500-1507. |
| [38] | Chang H, Zhou X B, Wang W H, et al. Effects of selenium application in soil on formation of iron plaque outside roots and cadmium uptake by rice plants[J]. Advanced Materials Research, 2013, 750-752: 1573-1576. DOI:10.4028/www.scientific.net/AMR.750-752.1573 |
| [39] |
郭锋, 樊文华, 冯两蕊, 等. 硒对镉胁迫下菠菜生理特性、元素含量及镉吸收转运的影响[J]. 环境科学学报, 2014, 34(2): 524-531. Guo F, Fan W H, Feng L R, et al. Effects of selenium (Se) on the physiological characteristics, element contents, uptake and transportation of Cd in spinach under Cd stress[J]. Acta Scientiae Circumstantiae, 2014, 34(2): 524-531. |
| [40] |
万亚男. 硒对水稻吸收、转运及累积镉的影响机制[D]. 北京: 中国农业大学, 2018. Wan Y N. Mechanisms of selenium influencing cadmium uptake, translocation and accumlation in rice[D]. Beijing: China Agricultural University, 2018. |
| [41] |
李影, 王友保. 4种蕨类草本植物对Cu的吸收和耐性研究[J]. 草业学报, 2010, 19(3): 191-197. Li Y, Wang Y B. Research on Cu uptake and tolerance of four pteridophyta plants[J]. Acta Prataculturae Sinica, 2010, 19(3): 191-197. |
| [42] |
胡斌, 王丹, 王松山, 等. 铜、硒复合污染对小白菜生长及金属吸收和转运的影响[J]. 环境科学学报, 2011, 31(9): 2033-2041. Hu B, Wang D, Wang S S, et al. Uptake and transport of exogenous Cu and Se in pak choi and their effects on growth[J]. Acta Scientiae Circumstantiae, 2011, 31(9): 2033-2041. |
| [43] | Li H F, McGrath S P, Zhao F J. Selenium uptake, translocation and speciation in wheat supplied with selenate or selenite[J]. New Phytologist, 2008, 178(1): 92-102. DOI:10.1111/j.1469-8137.2007.02343.x |