 2021, Vol. 42
2021, Vol. 42


2. 西安建筑科技大学西北水资源与环境生态教育部重点实验室, 西安 710055
2. Key Laboratory of Northwest Water Resource, Environment and Ecology, Ministry of Education, Xi'an University of Architecture and Technology, Xi'an 710055, China
已有研究表明, 城市污水处理厂进水中存在大量的活性硝化菌[1~3].Frigon等[4]的研究发现, 进水硝化菌在迁移至活性污泥后仍能恢复硝化性能, Duan等[5]的研究发现进水硝化菌的迁移会引起系统短程硝化失效, Yu等[6]的研究证明进水硝化菌可以有效提高活性污泥硝化速率; Saunders等[7]的研究比较了3个污水厂进水与活性污泥中硝化菌, 发现二者具有相似的群落结构, 并认为硝化菌的选择机制在污水管网中就已经开始.以上研究说明进水硝化菌对活性污泥系统具有接种且强化活性污泥系统硝化性能的潜力.但目前这些研究仅对进水硝化菌是否具有活性, 其与活性污泥硝化菌群落结构是否相似进行调查.在城市污水处理厂实际运行过程中, 由于硝化菌对温度变化敏感, 曝气池中硝化菌活性与群落结构随季节规律性变化, 并且冬季硝化效率是曝气池容积设计的限制因素.然而, 进水硝化菌对活性污泥系统硝化性能与群落结构的季节性变化规律之间是否具有相关性, 进水硝化菌的迁移能否缓减季节性温度变化对活性污泥系统的影响, 这些问题有待进一步研究.
本研究在未设置初沉池的西安市第二污水处理厂中, 对其进水和活性污泥中氨氧化菌(AOB)的硝化性能和群落结构进行长达12个月的调查分析.研究进水和活性污泥的比氨氧化速率(SAUR)的季节性变化, 并通过活性计算进水AOB对活性污泥系统的接种强度; 采用qPCR检测进水与活性污泥AOB的丰度变化, 揭示进水硝化菌对活性污泥系统的影响; 同时, 通过Illumina MiSeq测序比较进水和活性污泥系统中AOB的群落结构, 以期为优化污水处理厂设计与运行提供一定的理论支持.
1 材料与方法 1.1 实验样品考虑到城市污水处理厂中初沉池污泥停留时间较长, 进水硝化菌可能会在初沉池中被分离[8], 因此本实验样品取自未设置初沉池的西安市第二污水处理厂.西安市第二污水处理厂是西安市服务范围最大的污水处理系统, 其污水管网的水量大约为50万m3·d-1, 污水在管网中的平均停留时间约为十几个小时, 高于该厂曝气池水力停留时间.
实验所用进水样品及活性污泥样品分别取自细格栅出水处及多段多级AAO好氧池末端.设置取样周期为每月一次(每月10日), 并在相同时间段(09:30~12:00)进行取样, 雨天则推迟3 d取样.
1.2 比氨氧化速率(SAUR)及接种强度测定取进水样品10 L, 多次浓缩后用去离子水清洗3次以去除样品中残余基质(5 000 r·min-1, 5 min), 重悬至1 000 mL后用于氨氧化速率(AUR)的测定.取适量活性污泥样品直接进行离心操作, 其预处理步骤与进水样品相同.AUR采用耗氧速率法(OUR)在(20±1)℃条件下测定, 具体方法见文献[9], 并定义AUR与混合液挥发性悬浮固体浓度(MLVSS)的比值为比氨氧化速率(SAUR).
进水硝化菌对活性污泥系统的接种强度计算公式[6]如下:
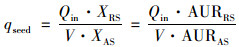
|
(1) |
式中, qseed为硝化菌连续接种强度, g·(g·d)-1; Qin为城市污水处理厂的平均日进水量, L·d-1; XRS为污水厂进水中硝化菌浓度, mg·L-1; V为城市污水处理厂生物反应池容积, L; XAS为活性污泥中硝化菌浓度, mg·L-1; AURRS为进水硝化菌氨氧化活性, mg·(L·h)-1; AURAS为活性污泥硝化菌氨氧化活性, mg·(L·h)-1.
1.3 群落结构测定 1.3.1 定量PCR(qPCR)取适量浓缩后的进水污泥和活性污泥样品(1.2节)于4℃下10 000 r·min-1离心5 min, 弃上清液后置于-20℃下密封保存.DNA提取步骤及纯度鉴定方法见文献[10].采用qPCR技术对进水和活性污泥中AOB(目标基因为amoA)进行定量分析, 正向引物为amoA-1F(GGGGTTTCTACTGGTGGT), 反向引物为amoA-2R(CCCCTCTGCAAAGCCTT CTTC), 具体qPCR扩增体系构建及扩增条件见文献[10].根据Harms等[11]的研究, 拷贝数与细胞数的比例为2∶1, 将qPCR所得拷贝数按比例转换为细胞数.
1.3.2 Illumina MiSeq高通量测序从2%琼脂糖凝胶中提取PCR产物, 用AxyPrepDNA凝胶回收试剂盒(AXYGEN公司, 美国)进行纯化, 将PCR产物用QuantiFlourMT-ST蓝色荧光定量系统(Promega公司, 美国)进行检测定量, 之后按照每个样本的测序量要求, 进行相应比例的混合.构建MiSeq文库, 通过Illumina MiSeq PE300平台进行测序.对得到的原始数据进行质量控制, 并按照barcode标签序列识别并区分样品得到有效数据, 获得最终用于分析的序列.应用Qiime软件根据序列相似度获得操作分类单元(OTU), 采用Usearch软件设置97%的相似性, 进行分类学分析, 在各个分类水平上统计样品的群落组成.
1.4 相关性分析采用SPSS 23.0软件对实验数据进行统计学分析, 用r表示皮尔逊相关性系数, 当0<r<1代表两变量正相关, -1<r<0代表两变量负相关.其中, |r|≥0.8时, 代表两变量高度相关; 0.5≤|r|≤0.8时, 代表两变量中度相关; 0.3≤|r|≤0.5时, 代表两变量低度相关; |r|<0.3时, 代表两变量相关性弱.当显著性水平P<0.05, 代表两变量具有显著相关性.
2 结果与讨论 2.1 进水与活性污泥硝化性能相关性分析图 1为进水和活性污泥比氨氧化速率(SAUR)及温度的历时变化.如图 1(a)所示, 进水和活性污泥的SAUR分别为0.48~3.02 mg·(g·h)-1和0.68~2.25 mg·(g·h)-1, 从1~12月总体呈先下降后上升的趋势, 相关性分析结果表明, 进水与次月活性污泥SAUR高度相关(r=0.862, P<0.05), 即进水AOB活性会对次月活性污泥的硝化性能有显著影响.

|
图 1 进水与活性污泥的SAUR及温度历时变化 Fig. 1 SAUR and temperature of raw sewage and activated sludge |
由图 1(b)可得, 冬季进水SAUR[(2.19±0.53)mg·(g·h)-1]明显高于夏季[(0.77±0.25)mg·(g·h)-1].AOB生长及活性受环境因素影响, 如温度、溶解氧和pH等.污水管道中氨氮充足, pH在7~8之间, 溶解氧约为0.3 mg·L-1[12], 由此看出溶解氧是污水管网中AOB生长及活性的主要限制因素, 而污水管网中的溶解氧恢复主要依靠自然复氧, 温度与水中自然复氧速率呈负相关[13].如图 1(b)所示, 冬季进水温度为(15.4±0.71)℃, 低于夏季进水温度(22℃), 从而导致冬季自然复氧速率高于夏季, 因此冬季进水SAUR较高.此外, 秋、冬季进水SAUR分别为(1.14±0.45) mg·(g·h)-1和(2.19±0.53) mg·(g·h)-1, 高于同期活性污泥, 这可能是因为污水管道常年埋在冻土层下方, 在秋冬季节, 管道中的水温高于曝气池水温, 从而导致秋冬季进水SAUR高于同期活性污泥.
值得一提的是, 冬季活性污泥SAUR为(2.11±0.73) mg·(g·h)-1, 明显高于春季[(1.93±0.18) mg·(g·h)-1]和夏季[(1.13±0.33) mg·(g·h)-1].而多数研究表明城市污水处理厂冬季AOB活性通常较低, 容易导致出水氨氮上升[14~16], 造成这一矛盾可能有两方面原因:其一, 正常运行的污水处理系统中, 曝气池中硝化菌增殖速率的主要限制因素是氮浓度, 且冬季与夏季氮负荷差异不大, 硝化菌产率系数一致, 但是夏季硝化菌衰减系数更大; 其二, 由于西安市第二污水处理厂未设置初沉池, 使得绝大多数的进水硝化菌得以迁移至活性污泥系统中, 起到强化硝化的作用.Duan等[5]的研究同样指出, 污水处理厂中初沉池的设置与否很大程度上会影响进水硝化菌的迁移效率.迁移效率的高低表示进水硝化菌进入曝气池浓度的大小.Yu等[6]研究进水硝化菌的迁移对活性污泥系统的影响, 分别在2个序批式反应器(SBR)中以无菌污水(对照组)及含菌污水(实验组)为进水, 结果表明实验组中的氨氧化活性是对照组的1.15倍.这表明硝化菌的迁移能强化活性污泥系统硝化性能, 进一步说明不设置初沉池的活性污泥系统硝化性能更强.由图 1(b)可知进水SAUR在冬季时最高, 因此经进水硝化菌迁移后的活性污泥系统在冬季时的硝化性能也强于夏季.
综上可以推断, 不设置初沉池可能更有利于强化污水处理厂冬季硝化活性, 进水硝化菌的迁移能减缓季节性温度变化对活性污泥系统硝化性能的不利影响, 但具有一定滞后性.
2.2 进水氨氧化菌对活性污泥的接种强度接种强度是衡量进水硝化菌对活性污泥系统影响的重要指标之一, 采用式(1)计算进水AOB对活性污泥系统的接种强度, 结果如图 2所示.AOB接种强度为0.21~0.92 g·(g·d)-1, 平均接种强度为0.475 g·(g·d)-1, 研究结果与蒙特利尔LaPrairie污水处理厂中进水硝化菌的接种强度类似[0.3 g·(g·d)-1][1].通常认为温度对接种强度有明显地影响[9], 分别对进水温度、曝气池温度与接种强度进行相关性分析, 结果显示进水及曝气池温度与次月接种强度呈中度相关(r=0.722, P<0.05; r=0.667, P<0.05).这可能存在两方面原因, 其一, 进水温度较低时, 污水管网中溶解氧浓度较高, 进水硝化菌活性较大, 经迁移后强化次月活性污泥硝化性能; 其二, 活性污泥中氨氮浓度受限, 硝化菌产率系数一致, 但在低温下硝化菌衰减系数较小, 同样能在次月表现出较大的活性.

|
图 2 进水AOB对活性污泥系统连续接种强度的历时变化 Fig. 2 Continuous seeding intensity of influent AOB to activated sludge |
值得一提的是, 接种强度会影响污水处理厂最低污泥龄的设计[17], 以夏季[(0.43±0.05) g·(g·d)-1]与冬季[(0.35±0.11) g·(g·d)-1]的接种强度为例, 根据Jauffur等[1]的研究计算得出夏季最低污泥龄在理论上可以减少0.39d, 冬季最低污泥龄在理论上能减少0.87 d, 而最低污泥龄是生物脱氮系统曝气池容积设计的关键性因素[18].由此可见, 在活性污泥模型及污水厂工艺的设计优化中, 有必要考虑到进水硝化菌的迁移对活性污泥系统的影响, 特别是初沉池的设计会影响进水硝化菌的迁移效率.
2.3 进水与活性污泥中硝化菌的丰度采用qPCR技术对不同月份中进水和活性污泥的AOB丰度进行检测.由图 3可知, 进水AOB丰度为1.32×108~2.36×109 cells·g-1, 活性污泥AOB丰度为1.12×1010~1.19×1010 cells·g-1.进水AOB丰度较高是因为污水管道中是微氧环境[12], 污水在管道中停留时间较长且温度相对稳定, 这为硝化菌的大量增殖创造了有利条件.有关城市污水厂硝化菌丰度的研究中, 处理水量与本研究中取样水厂相似, 但工艺流程中设有初沉池, 其活性污泥系统中AOB丰度仅为108~109 cells·g-1[19, 20].由此可推测, 初沉池的设计会对进水硝化菌的迁移产生影响, 并且进水硝化菌会对活性污泥系统硝化菌丰度做出贡献.值得一提的是, 进水及活性污泥中AOB丰度并不与其SAUR相对应, 这可能是因为在污水管网及活性污泥中, AOB生长及活性都受到一定抑制, 从而导致其活性与丰度不能相互对应[21].

|
图 3 进水与活性污泥AOB丰度的历时变化 Fig. 3 Abundance of AOB in raw sewage and activated sludge |
此外, 活性污泥在夏季时的AOB丰度略高于秋冬季节.理论上, 硝化菌属于自养菌, 增殖速率较慢, 且易受温度的影响而产生数量波动[22], 尤其是在秋冬季节活性污泥中硝化菌丰度可能会因温度降低而大幅下降.在本研究中, 秋冬季活性污泥AOB丰度虽有下降, 但仍然保持在同一数量级上(1010 cells·g-1).由此推断, 进水硝化菌的迁移能缓解温度降低对活性污泥中硝化菌丰度的负面影响.
2.4 进水AOB对活性污泥系统群落结构的影响 2.4.1 OTUs从进水及活性污泥样品中OTU数来分析两者微生物多样性, 具体结果如表 1所示.6个进水样品及6个活性污泥样品中AOB有效OTUs分别在126~280和104~116范围内, 这说明进水与活性污泥中AOB的生物多样性存在差异.值得注意的是, 各月份活性污泥样品中OTUs都低于进水样品, 这是因为经过污水处理厂工艺条件的筛选, 进水AOB中一些不适应曝气池环境的菌群被淘汰, 从而导致活性污泥系统中生物多样性较低.
|
|
表 1 进水AOB和活性污泥AOB的多样性1) Table 1 Diversity of nitrifiers in raw sewage and activated sludge |
此外, 每个月份中进水与活性污泥AOB中均具有一定的共有OTUs(45~75), 且共有OTUs在活性污泥中占比的范围为40%~50%, 在进水中占比的范围为15%~30%, 二者相关性分析结果表明, 共有OTUs在进水中的占比与在活性污泥中的占比高度相关(r=0.988, P<0.05), 而每一类OTUs代表一类菌种[23], 因此, 进水与活性污泥有相同的共有优势菌, 且进水AOB的群落结构会显著影响活性污泥群落结构的形成.Frigon等[4]在研究污水厂进水硝化菌迁移理论中指出, 硝化菌在污水处理厂中生物多样性较低, 且竞争能力较弱, 经迁移后的进水硝化菌能与活性污泥硝化菌进行竞争.由此进一步推断, 活性污泥群落结构的构建符合中性(迁移)理论, 中性(迁移)理论遵循随机漂移的过程, 微生物从一个群落迁移到另一个群落, 认为具有相同生态功能的物种具有中性竞争性, 并考虑了出生、死亡和通过迁移的有限扩散的随机过程, 以解释群落的组成和结构[24~26].Ofiteru等[27]研究Palo Alto污水厂细菌种群动态, 结果表明AOB种群动态和丰度分布符合中性理论, 并指出AOB群落结构可能是由中性(迁移)理论及生态位理论共同作用下构建的.
2.4.2 群落结构组成分析在属和种水平上对测序结果进行归类, 对比分析所取的6个进水与6个活性污泥样品在不同分类水平上的菌群组成及相对丰度, 图 4为进水和活性污泥AOB属水平群落结构组成.其中, Nitrosomonas菌属在进水和活性污泥中的相对丰度最高, 为两者的共同优势菌属.然而, Nitrosomonas菌属在进水AOB中相对丰度为38.3%~69.4%, 而在活性污泥中相对丰度为52.8%~89.7%.刘文如等[28]在研究不同溶解氧浓度下硝化菌群落结构变化中发现, Nitrosomonas菌属在溶解氧较高的环境中相对丰度较高.因此, 污水管道与曝气池中环境因子的不同(如溶解氧和温度等环境因素)是造成进水和活性污泥中硝化菌相对丰度差异的重要原因.

|
RS为进水AOB, AS为活性污泥AOB, 下同 图 4 进水及活性污泥中AOB属水平群落结构 Fig. 4 Community structures of AOB in raw sewage and activated sludge by genus |
进水与活性污泥AOB种水平群落结构组成如图 5所示, 进水和活性污泥AOB具有多种共同菌种, 其中Nitrosomonas sp. Nm58、Nitrosomonas sp. JL21和bacterium CYCU-0253为两者共同的优势菌种.Nitrosomonas sp. Nm58通常被认为广泛存在于下水道及低溶解氧的生物膜中[29, 30], 因此城市污水处理厂进水中的Nitrosomonas sp. Nm58相对丰度较高.然而如Nitrosococcus sp. LT-3存在于进水中却没有在活性污泥中被检测到, Ye等[31]对中国香港沙田污水处理厂进水和活性污泥等区域进行群落结构分析后同样发现, 进水中的一些菌群不会在活性污泥中增殖.这表明进水硝化菌迁移至活性污泥系统后, 受环境因子(如温度和溶解氧等)的影响而导致群落结构发生变化.

|
图 5 进水及活性污泥中AOB种水平群落结构组成 Fig. 5 Community structures of AOB in raw sewage and activated sludge by species |
先前研究通常认为活性污泥群落结构的形成仅仅受环境因子(如温度和溶解氧等)的影响, 并认为硝化菌的增殖仅来源于生物脱氮系统内部(如曝气池中), 即通过生态位理论来解释活性污泥群落结构的形成并指导污水处理厂的设计.本研究结果表明, 在未设置初沉池的污水处理厂中, 进水硝化菌与活性污泥系统在硝化性能与群落结构方面有显著相关性, 且由于活性污泥中硝化菌生物多样性较低, 进水硝化菌能与活性污泥硝化菌进行中性竞争.因此, 活性污泥群落结构的形成是中性(迁移)机制和生态机制共同作用的结果.
3 结论(1) 在未设置初沉池的污水处理厂中, 进水硝化菌与活性污泥系统存在高度相关性, 进水硝化菌的迁移能减缓秋冬季温度降低对活性污泥系统硝化性能不利的影响, 但具有一定的滞后性.
(2) 进水硝化菌的迁移能弥补季节性温度变化而引起的活性污泥硝化菌丰度降低, 并起到接种作用, 在活性污泥模型及污水处理厂设计中, 应当考虑到进水硝化菌对活性污泥系统的影响.
(3) 活性污泥硝化菌群落结构与进水硝化菌群落结构具有高度相关性, 活性污泥群落结构的形成是中性(迁移)机制和生态机制共同作用的结果.
| [1] | Jauffur S, Isazadeh S, Frigon D. Should activated sludge models consider influent seeding of nitrifiers? Field characterization of nitrifying bacteria[J]. Water Science & Technology, 2014, 70(9): 1526-1532. |
| [2] | Chen G H, Leung D H W, Hung J C. Biofilm in the sediment phase of a sanitary gravity sewer[J]. Water Research, 2003, 37(11): 2784-2788. DOI:10.1016/S0043-1354(03)00083-6 |
| [3] |
金鹏康, 王斌. 城市污水管网对水质的生化改善特性[J]. 环境工程学报, 2016, 10(7): 3401-3408. Jin P K, Wang B. Improvement in biodegradability of wastewater quality in urban sewer system[J]. Chinese Journal of Environmental Engineering, 2016, 10(7): 3401-3408. |
| [4] | Frigon D, Wells G. Microbial immigration in wastewater treatment systems: analytical considerations and process implications[J]. Current Opinion in Biotechnology, 2019, 57: 151-159. DOI:10.1016/j.copbio.2019.02.021 |
| [5] | Duan H R, Ye L, Wang Q L, et al. Nitrite oxidizing bacteria (NOB) contained in influent deteriorate mainstream NOB suppression by sidestream inactivation[J]. Water Research, 2019, 162: 331-338. DOI:10.1016/j.watres.2019.07.002 |
| [6] | Yu L F, Li R, Delatolla R, et al. Natural continuous influent nitrifier immigration effects on nitrification and the microbial community of activated sludge systems[J]. Journal of Environmental Sciences, 2018, 74: 159-167. DOI:10.1016/j.jes.2018.02.021 |
| [7] | Saunders A M, Albertsen M, Vollertsen J, et al. The activated sludge ecosystem contains a core community of abundant organisms[J]. The ISME Journal, 2016, 10(1): 11-20. DOI:10.1038/ismej.2015.117 |
| [8] |
张潇, 赵博玮, 岳秀萍. 取消初沉池对AAO工艺活性污泥微生物结构的影响[J]. 工业水处理, 2019, 39(6): 30-34. Zhang X, Zhao B W, Yue X P. Influences of canceling primary settling tanks on the microbial structure of activated sludge in AAO process[J]. Industrial Water Treatment, 2019, 39(6): 30-34. |
| [9] |
于莉芳, 杜倩倩, 傅学焘, 等. 城市污水中硝化菌群落结构与性能分析[J]. 环境科学, 2016, 37(11): 4366-4371. Yu L F, Du Q Q, Fu X T, et al. Community structure and activity analysis of the nitrifiers in raw sewage of wastewater treatment plants[J]. Environmental Science, 2016, 37(11): 4366-4371. |
| [10] |
李韧, 于莉芳, 张兴秀, 等. 硝化生物膜系统对低温的适应特性: MBBR和IFAS[J]. 环境科学, 2020, 41(8): 3691-3698. Li R, Yu L F, Zhang X X, et al. Adaptability of nitrifying biofilm systems to low temperature: MBBR and IFAS[J]. Environmental Science, 2020, 41(8): 3691-3698. |
| [11] | Harms G, Layton A C, Dionisi H M, et al. Real-time PCR quantification of nitrifying bacteria in a municipal wastewater treatment plant[J]. Environmental Science & Technology, 2003, 37(2): 343-351. |
| [12] | Jin P K, Wang B, Jiao D, et al. Characterization of microflora and transformation of organic matters in urban sewer system[J]. Water Research, 2015, 84: 112-119. DOI:10.1016/j.watres.2015.07.008 |
| [13] | 张自杰. 排水工程-下册[M]. (第五版). 北京: 建筑工业出版社, 2015: 113-135. |
| [14] |
陈翰, 马放, 李昂, 等. 低温条件下污水生物脱氮处理研究进展[J]. 中国给水排水, 2016, 32(8): 37-43. Chen H, Ma F, Li A, et al. Research progress in biological nitrogen removal from wastewater under low temperature condition[J]. China Water & Wastewater, 2016, 32(8): 37-43. |
| [15] |
韩文杰, 吴迪, 周家中, 等. 长三角地区IFAS污水厂低温季节微生物多样性分析[J]. 环境科学, 2020, 41(11): 5037-5049. Han W J, Wu D, Zhou J Z, et al. Microbial diversity analysis of WWTPs based on IFAS in low temperature season in Yangtze River delta[J]. Environmental Science, 2020, 41(11): 5037-5049. |
| [16] | Delatolla R, Young B, Stintzi A. Application of the MBBR technology to achieve nitrification below 1℃: biofilm and microbiome analysis[A]. Frontiers in Wastewater Treatment and Modelling[C]. Cham: Springer, 2017.513-517. |
| [17] |
于莉芳, 滑思思, 莫鹏程, 等. 原生硝化菌对活性污泥系统影响研究进展[J]. 工业水处理, 2019, 39(11): 12-16. Yu L F, Hua S S, Mo P C, et al. Research progress on the effect of natural nitrifier immigration on the activated sludge system[J]. Industrial Water Treatment, 2019, 39(11): 12-16. |
| [18] |
李卫平, 韩剑宏, 延克军. 曝气池容积计算方法分析[J]. 环境工程, 2004, 22(4): 24-25. Li W P, Han J H, Yan K J. Analysis of the calculative ways of aeration tanks' volumes[J]. Environmental Engineering, 2004, 22(4): 24-25. |
| [19] |
曾薇, 张丽敏, 王安其, 等. 污水处理系统中硝化菌的菌群结构和动态变化[J]. 中国环境科学, 2015, 35(11): 3257-3265. Zeng W, Zhang L M, Wang A Q, et al. Community structures and population dynamics of nitrifying bacteria in activated sludges of wastewater treatment plants[J]. China Environmental Science, 2015, 35(11): 3257-3265. |
| [20] | Kayee P, Sonthiphand P, Rongsayamanont C, et al. Archaeal amoA genes outnumber bacterial amoA genes in municipal wastewater treatment plants in Bangkok[J]. Microbial Ecology, 2011, 62(4): 776-788. |
| [21] | Liu W L, Yang Q, Ma B, et al. Rapid achievement of nitritation using aerobic starvation[J]. Environmental Science & Technology, 2017, 51(7): 4001-4008. |
| [22] |
尚越飞, 王申, 宗倪, 等. 污水生物处理工艺低温下微生物种群结构[J]. 环境科学, 2020, 41(10): 4636-4643. Shang Y F, Wang S, Zong N, et al. Microbial community structure for sewage wastewater treatment plants in winter[J]. Environmental Science, 2020, 41(10): 4636-4643. |
| [23] |
曾涛涛, 蒋小梅, 韩科昌, 等. 生活污水处理厂微生物群落结构解析[J]. 安全与环境学报, 2018, 18(2): 697-703. Zeng T T, Jiang X M, Han K C, et al. Analysis of microbial community constituent composition of some sewage treatment and processing plant[J]. Journal of Safety and Environment, 2018, 18(2): 697-703. |
| [24] | Mei R, Liu W T. Quantifying the contribution of microbial immigration in engineered water systems[J]. Microbiome, 2019, 7(1): 144. |
| [25] |
牛克昌, 刘怿宁, 沈泽昊, 等. 群落构建的中性理论和生态位理论[J]. 生物多样性, 2009, 17(6): 579-593. Niu K C, Liu Y N, Shen Z H, et al. Community assembly: the relative importance of neutral theory and niche theory[J]. Biodiversity Science, 2009, 17(6): 579-593. |
| [26] |
周淑荣, 张大勇. 群落生态学的中性理论[J]. 植物生态学报, 2006, 30(5): 868-877. Zhou S R, Zhang D Y. Neutral theory in community ecology[J]. Journal of Plant Ecology, 2006, 30(5): 868-877. |
| [27] | Ofiteru I D, Lunn M, Curtis T P, et al. Combined niche and neutral effects in a microbial wastewater treatment community[J]. Proceedings of the National Academy of Sciences of the United States of America, 2010, 107(35): 15345-15350. |
| [28] |
刘文如, 顾广发, 宋小康, 等. 不同溶解氧浓度下硝化工艺中微生物种群结构对比[J]. 环境科学, 2019, 40(8): 3706-3712. Liu W R, Gu G F, Song X K, et al. Comparison of the microbial community structure in nitrifying processes operating with different dissolved oxygen concentrations[J]. Environmental Science, 2019, 40(8): 3706-3712. |
| [29] | Purkhold U, Pommerening-Roser A, Juretschko S, et al. Phylogeny of all recognized species of ammonia oxidizers based on comparative 16S rRNA and amoA sequence analysis: Implications for molecular diversity surveys[J]. Applied and Environmental Microbiology, 2000, 66(12): 5368-5382. |
| [30] | Park H D, Noguera D R. Characterization of two ammonia-oxidizing bacteria isolated from reactors operated with low dissolved oxygen concentrations[J]. Journal of Applied Microbiology, 2007, 102(5): 1401-1417. |
| [31] | Ye L, Zhang T. Bacterial communities in different sections of a municipal wastewater treatment plant revealed by 16S rDNA 454 pyrosequencing[J]. Applied Microbiology and Biotechnology, 2013, 97(6): 2681-2690. DOI:10.1007/s00253-012-4082-4/figures/5 |