 2021, Vol. 42
2021, Vol. 42



2. 中国科学院大学, 北京 100049;
3. 江苏省太仓市璜泾镇人民政府, 太仓 215427;
4. 南京万德斯环保科技股份有限公司, 南京 211100
2. University of Chinese Academy of Science, Beijing 100049, China;
3. Huangjing Town People's Government, Taicang 215427, China;
4. Nanjing Wondux Environmental Protection Technology Co., Ltd., Nanjing 211100, China
营养盐的过度输入引起浮游植物的异常繁殖并引起藻华暴发.太湖的富营养化始自20世纪80年代, 蓝藻水华覆盖面积从开始时的梅梁湾逐步扩大到整个西北太湖, 种类以微囊藻为主[1, 2].浮游植物在生长、死亡过程会释放溶解性有机质(DOM), 根据其存在形式可以分为胞外溶解性有机质(E-DOM)和胞内溶解性有机质(I-DOM).E-DOM指的是藻类在生长繁殖过程中释放到水体中的代谢产物; 与E-DOM对应, 细胞衰退阶段, 藻细胞裂解会释放大量的I-DOM, 大幅提高溶液的DOC浓度[3].DOM在水体中会发生光化学降解或生物降解等转化过程, 影响有机质的归趋[4~8].在自然水体中, 这两种转化可能存在耦合作用或竞争作用[9~11].耦合作用指的是较难生物降解后的DOM经光转化后可以提高其生物降解性, 或者生物降解后的产物能进行光降解; 竞争作用是两种作用均对DOM有较好的转化.
DOM的光化学反应、生物降解性与其性质、组成密切相关[10, 12~16].三维荧光光谱近年来广泛应用于DOM组成和来源的研究, 具有灵敏度高和信息量丰富等优点[17].Xiao等[18]研究了4种微囊藻的胞外有机质特征, 其荧光组分包括1种类蛋白组分和3种类腐植酸组分.笔者以往的研究表明太湖蓝藻I-DOM中含有类色氨酸和类腐植酸两种物质:类色氨酸物质在紫外和可见光条件下均能快速降解; 类腐植酸物质在紫外光下可以降解, 在可见光照条件下降解很慢[19].有研究考察了太湖蓝藻水华期湖水中溶解性有机质的生物降解[20], 但其对象为蓝藻水华暴发期的湖水DOM, 包含各种来源有机质.到目前为止, 对于太湖蓝藻胞内有机质的微生物降解的研究仍有限.探明太湖蓝藻I-DOM的生物降解过程对明确水华消亡产生DOM的归趋具有重要的作用; 通过结合I-DOM光降解过程, 可推断I-DOM中各物质的最终归趋.
1 材料与方法 1.1 I-DOM的提取于2016年6月14日在中国科学院太湖湖泊生态系统试验站(120°12.57′E, 31°25.42′N)采集含蓝藻水样10 L, 再通过浮游植物网过滤得到含蓝藻浓度较高的水样; 对过滤后的藻水进行离心(9 000 r·min-1, 15 min), 分离出藻浆; 然后将藻浆放入超纯水中混合清洗, 再离心出藻浆, 重复3遍; 最后用液氮将蓝藻快速冷冻, 再解冻; 重复3遍, 将蓝藻细胞打破.处理后的溶液经孔径为0.22 μm的滤膜(Millipore)过滤得到I-DOM溶液[21, 22], 冷藏.经测定, 储备液浓度为550 mg·L-1.
1.2 I-DOM的生物降解I-DOM的生物降解反应是在温控培养箱中避光进行.本实验一共设置不同温度(20、25和30℃)和不同I-DOM初始浓度(5、10和20 mg·L-1)系列实验, 共5个实验组, 每个实验组设置3个平行, 实验周期14 d.反应前将I-DOM储备液配制成相应浓度的I-DOM溶液(5、10和20 mg·L-1)800 mL, 调节溶液pH为8.0; 接种0.8 mL菌种水(取自采样点, 经0.7 μm滤膜过滤); 为了减少接种湖水中DOM对本实验的干扰, 同时考虑到I-DOM生物可利用性强, 所以接种比为1∶1 000; 为了减少外界污染, 瓶口采用医用棉覆盖.在0、1、2、4、7、10和14 d取样.样品经0.22 μm的滤膜过滤后, 冷藏, 在两周内进行DOC浓度、紫外-可见光全波扫描(UV-Vis)和三维荧光光谱(EEM)分析.
1.3 I-DOM分析方法溶解性有机质(DOC)浓度的测定:采用TOC仪(TOC-VCPN)测定.
紫外-可见光全波扫描光谱(UV-Vis Spectra)测定:使用Lambda 35(Perkin Elmer)紫外可见分光光度计, 测试范围为200~800 nm, 参比为Milli-Q水.根据式(1)和(2), 通过一定波长下的吸光度计算得到该波长下的吸收系数[23].
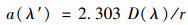
|
(1) |
式中, a(λ′)为波长λ下未校正的吸收系数(m-1); D(λ)为吸光度; r为光程路径(0.01 m).由于过滤清液可能残留细小颗粒会引起散射, 为此作如下散射效应订正[24]:
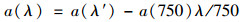
|
(2) |
式中, a(λ)为波长λ的吸收系数(m-1).
三维荧光光谱的测定(EEM):采用日立F-7000荧光光谱仪测定(Hitachi High-Technologies).激发波长(Ex)为200~450 nm, 间隔5 nm; 发射波长(Em)为250~550 nm, 间隔1 nm.EEM-PARAFAC模型的建立:首先对三维荧光光谱进行校正:①利用紫外-可见吸收光谱数据进行内滤效应修正[25]; ②用Milli-Q水做空白扣除; ③利用Milli-Q水的拉曼单位归一化(R.U.)来校准日常的荧光强度的变化.再利用DOMFluor工具箱在MATLAB R2008a软件中得到4组分模型(C1~C4), 通过折半分析(split-half analysis)和随机初始化(random initialization)方法来验证模拟结果的可靠性.
光学指标计算:E2∶E3为250 nm与365 nm吸光系数之比, 比值越大(超过3.5), 分子量越小, 腐植酸比例越小, 富里酸比例越大[26].Sr:波长275~295 nm和350~400 nm的光谱斜率的比值; 与分子量的大小成反比[27].HIX为腐殖化指数, 是激发波长为254 nm, 发射波长435~480 nm范围荧光强度的平均值与300~345 nm范围荧光强度平均值的比值[28].
1.4 统计分析不同温度、不同I-DOM初始浓度对生物降解的影响采用均值检验分析, P<0.05表示显著差异, P>0.05表示无显著差异.
2 结果与分析 2.1 I-DOM的EEM-PARAFAC组分根据不同降解条件实验获得的EEM数据, 建立了4个组分(C1~C4)模型(图 1).C1组分在275 nm存在激发波长最大值, 发射波长最大值出现于332 nm, 表征类色氨酸物质[29].C2组分有两个最大的激发波长, 分别在270和365 nm处, 最大发射波长为448 nm, 与Cawley等[30]研究的C3组分以及Chen等[31]研究的C3峰位置相近, 是一种广泛存在的类腐殖质成分.C3组分在280 nm和340 nm处分别存在激发最大值, 发射波长最大值存在于408 nm处, 认为是微生物作用产生的类腐殖质[22].C4组分的最大激发和发射波长分别为310 nm和392 nm, 也被认为是一种微生物活动产生的类腐殖质[32, 33].提取的I-DOM初始样品中, C1~C4组分的相对丰度分别为80.0%、16.0%、3.7%和0.3%.

|
图 1 I-DOM的EEM-PARAFAC组分及激发和发射波长特征 Fig. 1 EEM-PARAFAC components of I-DOM and characteristics of excitation and emission wavelengths |
图 2(a)显示了I-DOM在生物降解过程中(初始浓度DOC=10 mg·L-1, pH=8.0, T=25℃)的UV-Vis光谱变化, 可以看出I-DOM在紫外区有两个吸收峰, 分别为260 nm和330 nm, 随着I-DOM生物降解过程的进行, I-DOM的吸光度不断下降, 在250~350 nm处尤为明显.以260 nm处吸收系数(a260)的变化率(1-[a260]/[a260]0)表示I-DOM的降解率, 经过14 d的生物降解, I-DOM生物降解率达到74.6%; 前4 d的降解速率最大, 达到74.2%, 而后趋于稳定[图 2(b)]; DOC由初始的9.68 mg·L-1下降到3.80 mg·L-1, 去除率达到61.2% (表 1).

|
DOC=10 mg·L-1, pH=8.0, T=25℃ 图 2 I-DOM生物降解过程UV-Vis光谱、I-DOM的生物降解率和C1-C4组分变化 Fig. 2 Changes in the UV-Vis spectra, biodegradation ratio, and C1-C4 during the biodegradation of I-DOM |
|
|
表 1 不同生物降解条件下DOC的变化 Table 1 Changes in the DOC during the biodegradation of I-DOM under various conditions |
图 2(c)显示了I-DOM在生物降解过程中C1~C4组分的变化.可以看到, 类蛋白质组分C1在14 d的降解过程中持续下降; C1组分初始浓度为17.61 R.U., 在降解的第2 d浓度下降为8.38 R.U., 降解率达到52.43%, 随后降解变慢, 反应结束时(14 d)浓度为2.87 R.U., 降解率达到83.71%.类腐殖质组分C2~C4变化规律不相一致, C2组分在前2 d时降解较快, 由初始的3.51 R.U.下降到1.63 R.U., 降解率为53.38%; 但在2~7 d时浓度上升, 而后又缓慢下降, 14 d时浓度为1.76 R.U., 降解率为49.89%.C3和C4组分在降解过程中均呈现先上升后下降的变化, C3组分荧光强度的最大值出现在第4 d(2.61 R.U.), 而C4组分的最大值则出现在第10 d(1.21 R.U.).
图 3为I-DOM在生物降解过程中E2∶E3、Sr和HIX的变化.可以得到, 随着生物降解过程的进行, E2∶E3呈下降趋势, 特别是前2 d, 由7.78下降到4.25; 降解结束后, E2∶E3值为4.57; Sr值在第1 d时下降幅度最大, 由初始的2.41下降到1.03, 但在2 d时又迅速上升至2.02, 而后缓慢下降, 14 d时为1.41; HIX则不断上升, 由初始的0.48增加为1.43.

|
DOC=10 mg·L-1, pH=8.0, T=25℃ 图 3 生物降解过程中E2∶E3、Sr和HIX的变化 Fig. 3 Changes in the E2∶E3, Sr, and HIX during the biodegradation of I-DOM |
本实验设置了3个实验组(20、25和30℃)探究温度变化对I-DOM微生物降解的影响(图 4).20、25和30℃条件下, 经14 d微生物降解后DOC的去除率分别为56.5%, 61.2%和58.7%(表 1); 三者比较, 25℃为最佳的I-DOM微生物降解温度.在前4 d, 降解率逐渐上升, 25℃和30℃条件下a260降解率要显著高于20℃条件下(P<0.05), 4 d后3个温度实验组中a260的降解率逐渐趋于稳定.当14 d降解结束后, 20℃时C1组分的荧光强度要显著大于25℃和30℃实验组(P<0.05).C2组分的荧光强度在3个温度实验组中呈现出一致的趋势, 在反应前两天, C2组分减少; 后出现增加至峰值后又减少.3个温度下C2组分峰的大小和出现时间有所差异.20℃和30℃, C2组分的峰值均出现在第7 d, 分别为3.22 R.U.和3.74 R.U.; 25℃条件下, 峰值出现在反应的第4 d, 为2.65 R.U., 显著低于20℃和30℃的峰值(P<0.05).C3和C4组分14 d降解结束后荧光强度(第14 d)没有显著差异(P>0.05), 但C3的累积峰值在25℃和30℃时较20℃左移; C4的累积峰值在30℃时左移并显著高于25℃和20℃的峰值(P<0.05).另外, 14 d降解结束时, 25℃和30℃的HIX值要显著高于20℃(P<0.05), 表明适当升温可以促进I-DOM的腐殖化[图 4(f)].

|
DOC=10 mg·L-1, pH=8.0 图 4 温度变化对I-DOM生物降解过程的影响 Fig. 4 Effects of temperature on biodegradation of I-DOM |
设置3个实验组(初始DOC为5、10和20 mg·L-1)考察I-DOM的生物降解过程(图 5).[DOC]0为5、10和20 mg·L-1这3个不同I-DOM浓度, 经14 d微生物降解后DOC的去除率分别为50.4%, 61.2%和73.6%.在实验范围内, 更高的I-DOM初始浓度会促进其降解率的增加.前2 d, a260的降解率为20 mg·L-1>10 mg·L-1>5 mg·L-1(P<0.05); 4 d之后, 3个实验组a260的降解率逐渐稳定.对于PARAFAC组分, 3个不同初始DOC浓度下, 4个组分均呈现较一致的趋势.C1组分在14 d的微生物降解过程中均下降:反应前两天快速下降; 2~7 d出现停滞; 7 d后, 20 mg·L-1的初始浓度溶液中, C1组分的剩余率再次出现下降, 10 mg·L-1的初始浓度溶液中C1组分的剩余率也出现下降, 而5 mg·L-1的溶液中没有出现此现象.从整体上看, C1组分14 d后的剩余率为20 mg·L-1<10 mg·L-1<5 mg·L-1(P<0.05).3个实验组C2组分均在降解初期荧光强度降低后又增加出现峰值, 10和20 mg·L-1组C2组分的峰值相较于5 mg·L-1提早.C3和C4组分均先增后减, 初始浓度为20 mg·L-1的实验组C3组分峰值出现较其他两组早.

|
T=25℃, pH=8.0 图 5 初始DOC浓度对I-DOM生物降解的影响 Fig. 5 Effects of initial DOC concentrations on biodegradation of I-DOM |
从图 5(f)可以看出, 随着初始DOC浓度的上升, I-DOM在14 d生物降解结束后的HIX也越大(P<0.05), 腐殖化程度更高.
3 讨论 3.1 I-DOM降解过程的组分和性质变化I-DOM的初始荧光组分以类色氨酸C1占绝对优势(80%), 含有少量的类腐植酸C2(广泛存在, 16.0%, peak A+C); 与以往报道的胞内有机质DOM的特征一致, 类色氨酸组分占一些藻类胞内FDOM(荧光溶解性有机质)的75%~93%[22].以初始浓度在10 mg·L-1的I-DOM在25℃下的降解为例, 在反应前两天, C1和C2均出现大幅度的下降, 随后C1组分荧光强度下降速度变缓, 而C2组分由减少转为增加; C3和C4也出现了先增后减.C1组分荧光强度减少速度放缓, C2、C3和C4组分的增加表明了微生物生长过程会释放此类物质.不过后续C1的进一步减少、C2、C3和C4达到峰值后减少都提示这些物质能被微生物进一步消耗.细菌在消耗这些组分的同时, 也产生这些组分, 这也导致了在微生物转化过程中, 各组分的变化是比较复杂的, 观测到的各组分变化是其消耗和产生的净结果.湖泊中细菌对溶解性有机质的同时消耗和产生在文献中也有报道[34, 35].
光学指标E2∶E3和Sr呈现一致的下降趋势, 说明在微生物降解过程, 溶液中产生的物质的分子量增大.腐殖化指数HIX在微生物降解过程中的上升趋势表明, 溶液的腐殖化程度增加, 初始时溶液的HIX仅为0.42, 腐殖化程度很低, 存在的腐植酸仅占20%; 降解结束时, 溶液HIX为1.43, 腐殖化程度提高, 溶液中类腐殖质(C2、C3和C4)的丰度为52%.溶液DOM腐殖化程度的增加, 可能是其生物可利用性降低的原因; 这导致荧光组分在反应后期变化速率减缓.
3.2 温度或I-DOM初始浓度的影响在20~30℃范围内, 各PARAFAC组分的变化趋势一致, 说明在该温度范围内, 微生物的生长并不会受到限制.不过, 相对于25℃和30℃, 20℃实验组表现出较显著的延迟; 这可能是随着温度的降低, 微生物生长较为缓慢, 抑制其对有机质的利用和生长释放有机质过程较慢[36].太湖水华一般在10~11月份衰退, 此时水体温度为15~24℃[37].据本实验结果, 此温度下I-DOM依然能得到较好降解.
10 mg·L-1和20 mg·L-1的初始DOC浓度I-DOM的生物降解率要略高于5 mg·L-1的.较高浓度的I-DOM, 可能支撑更多的微生物生长, 微生物对I-DOM的利用导致较高的降解率.这一点与光降解相反, 光降解过程中, 由于光照(光强)一定, 提供的光子数量固定, 随着I-DOM浓度的增大, 单位DOM能利用光子的数量减少, 因此光降解率随I-DOM初始浓度的增大而减少.
3.3 与光降解结合推断I-DOM的可能归趋太湖水华消退阶段, 藻类释放的I-DOM在湖泊中能发生微生物降解和光降解.笔者前期对I-DOM的光降解研究[19]表明, 类色氨酸物质(C1)在紫外和可见光条件下均能快速降解; 而对于广泛存在的类腐殖质(C2)和微生物来源类腐殖质(C4), 可见光并不能使其降解; 同样地, UV-A区的紫外光(320~400 nm)不能降解C2和C4, 而UV-B区(280~400 nm)的紫外光能使40%~50%的C2和C4降解, 但不能完全降解.由于能进入到达地面的紫外光仅占5%左右, 且紫外光在水体中衰减很快, 仅能穿透几cm; 可以推断在实际水体中类腐殖质不能完全被光降解
而对于微生物降解, 在本研究结束时(14 d), C2组分还处于快速下降阶段(图 2); 虽然本研究未明确C2在长时间内能否完全降解, Zhang等[38]对微囊藻降解过程的研究, 与C2荧光特征相似的组分在反应14 d后, 荧光强度变化较小, 到21 d后还有略微上升, 说明微生物降解不能使C2完全降解.对于其他两种类腐殖质(C3和C4), 本研究结果显示, 随着反应的进行, 微生物来源类腐殖质(C3和C4)减少速率降低(图 2).因此, 光降解和微生物降解研究表明, I-DOM中的类腐殖质组分难以完全光降解或生物降解.除了光降解和微生物降解, 在颗粒性物质/金属矿物上的吸附或共沉淀是DOM的另一归趋[21, 39, 40].类腐殖质其具有芳香性, 较易发生吸附或共沉淀作用[14, 41].结合本实验结果(I-DOM的微生物降解)和光降解结果[19], 推测I-DOM在自然水体中的可能归趋.蓝藻水华消亡产生的I-DOM组分, 其归趋可能是:类色氨酸既能被光降解, 又能被生物降解, 而广泛存在的类腐殖质(C2)也能被光降解和生物降解, 生物降解可能更彻底地去除C2.微生物来源类腐殖质, 部分能被光降解或生物降解.在光降解或生物降解中, 类腐殖质难以彻底被去除; 由于其芳香性, 其可能发生沉降而从水体中去除, 埋藏在沉积物中.微生物作用过程中产生的这些难降解的物质的产生是水体碳埋藏的重要途径[42].不过在同时存在光降解和生物降解体系中, 类腐殖质组分是否可能被彻底去除还需要进一步研究.
4 结论(1) I-DOM的腐殖性小, 荧光组分以类色氨酸组分C1为主(80%), 存在少量的类腐殖质组分C2和微生物来源类腐殖质组分(C3、C4).微生物作用过程中, C2、C3和C4组分既被消耗, 同时又能产生.
(2) 在较低温度(类似于太湖水华消亡阶段的水体温度), I-DOM的降解在前两天有所延迟, 随后能有效被微生物降解; 5~20 mg·L-1的范围内的I-DOM能被微生物有效降解; 降解过程以20 mg·L-1的I-DOM最快.
(3) 结合各荧光组分的光降解结果, 推断太湖水华消亡产生的I-DOM可能有以下的归趋:类色氨酸能被光降解和生物降解; 而广泛存在的类腐殖质(C2)和微生物来源类腐殖质(C3、C4)能部分被光降解和生物降解, 但难以完全降解, 这些类腐殖质极可能与金属物质发生作用而从水体中去除, 埋藏在沉积物中.
| [1] |
刘俊杰, 陆隽, 朱广伟, 等. 2009-2017年太湖湖泛发生特征及其影响因素[J]. 湖泊科学, 2018, 30(5): 1196-1205. Liu J J, Lu J, Zhu G W, et al. Occurrence characteristics of black patch events and their influencing factors in Lake Taihu during 2009 and 2017[J]. Journal of Lake Sciences, 2018, 30(5): 1196-1205. |
| [2] |
辛华荣, 朱广伟, 王雪松, 等. 2009~2018年太湖湖泛强度变化及其影响因素[J]. 环境科学, 2020, 41(11): 4914-4923. Xin H R, Zhu G W, Wang X S, et al. Variation and driving factors of black water event intensity in Lake Taihu during 2009 to 2018[J]. Environmental Science, 2020, 41(11): 4914-4923. |
| [3] |
叶琳琳, 史小丽, 吴晓东, 等. 西太湖秋季蓝藻水华过后细胞裂解对溶解性有机碳影响[J]. 中国环境科学, 2011, 31(1): 131-136. Ye L L, Shi X L, Wu X D, et al. The effect of cyanobacteria on dissolved organic carbon post the bloom in autumn in Western Lake Taihu[J]. China Environmental Science, 2011, 31(1): 131-136. |
| [4] | Du Y X, Zhang Y Y, Chen F Z, et al. Photochemical reactivities of Dissolved Organic Matter (DOM) in a sub-alpine lake revealed by EEM-PARAFAC: An insight into the fate of allochthonous DOM in alpine lakes affected by climate change[J]. Science of the Total Environment, 2016, 568: 216-225. DOI:10.1016/j.scitotenv.2016.06.036 |
| [5] | Groeneveld M, Tranvik L, Natchimuthu S, et al. Photochemical mineralisation in a boreal brown water lake: considerable temporal variability and minor contribution to carbon dioxide production[J]. Biogeosciences, 2016, 13(13): 3931-3943. DOI:10.5194/bg-13-3931-2016 |
| [6] | Kellerman A M, Guillemette F, Podgorski D C, et al. Unifying concepts linking dissolved organic matter composition to persistence in aquatic ecosystems[J]. Environmental Science & Technology, 2018, 52(5): 2538-2548. |
| [7] |
刘堰杨, 孙辉, 刘琛, 等. 川西高原河流水体CDOM的光化学降解特性[J]. 环境科学, 2019, 40(12): 5318-5329. Liu Y Y, Sun H, Liu C, et al. Characteristics of chromophoric dissolved organic matter (CDOM) in natural rivers of Western Sichuan Plateau[J]. Environmental Science, 2019, 40(12): 5318-5329. |
| [8] |
祁闯, 方家琪, 张利民, 等. 太湖藻型湖区沉积物中生物易降解物质组成及分布规律[J]. 湖泊科学, 2019, 31(4): 941-949. Qi C, Fang J Q, Zhang L M, et al. Composition and distribution of biodegradable compounds in phytoplankton-dominated zone of Lake Taihu[J]. Journal of Lake Sciences, 2019, 31(4): 941-949. |
| [9] | Bittar T B, Vieira A A H, Stubbins A, et al. Competition between photochemical and biological degradation of dissolved organic matter from the cyanobacteria Microcystis aeruginosa[J]. Limnology and Oceanography, 2015, 60(4): 1172-1194. DOI:10.1002/lno.10090 |
| [10] | Bowen J C, Kaplan L A, Cory R M. Photodegradation disproportionately impacts biodegradation of semi-labile DOM in streams[J]. Limnology and Oceanography, 2020, 65(1): 13-26. DOI:10.1002/lno.11244 |
| [11] | Cory R M, Kling G W. Interactions between sunlight and microorganisms influence dissolved organic matter degradation along the aquatic continuum[J]. Limnology and Oceanography Letters, 2018, 3(3): 102-116. DOI:10.1002/lol2.10060 |
| [12] |
劳心宇, 原杰, 刘瑜, 等. 思林水库荧光溶解性有机质的特征、来源及其转化动力学[J]. 环境科学, 2019, 40(3): 1209-1216. Lao X Y, Yuan J, Liu Y, et al. Sources, characteristics and transformation dynamics of fluorescent dissolved organic matter in the Silin reservoir[J]. Environmental Science, 2019, 40(3): 1209-1216. |
| [13] | Sankar M S, Dash P, Singh S, et al. Effect of photo-biodegradation and biodegradation on the biogeochemical cycling of dissolved organic matter across diverse surface water bodies[J]. Journal of Environmental Science, 2019, 77: 130-147. DOI:10.1016/j.jes.2018.06.021 |
| [14] | Du Y X, Ramirez C E, Jaffé R. Fractionation of dissolved organic matter by co-precipitation with iron: effects of composition[J]. Environmental Processes, 2018, 5(1): 5-21. DOI:10.1007/s40710-017-0281-4 |
| [15] | Ward C P, Cory R M. Assessing the prevalence, products, and pathways of dissolved organic matter partial photo-oxidation in arctic surface waters[J]. Environmental Science: Processes & Impacts, 2020, 22(5): 1214-1223. |
| [16] | Zhou L, Zhou Y Q, Hu Y, et al. Microbial production and consumption of dissolved organic matter in glacial ecosystems on the Tibetan Plateau[J]. Water Research, 2019, 160: 18-28. DOI:10.1016/j.watres.2019.05.048 |
| [17] | Jaffé R, Cawley K M, Yamashita Y. Advances in the physicochemical characterization of dissolved organic matter: Impact on natural and engineered systems[M]. Washington: American Chemical Society, 2014: 27-73. |
| [18] | Xiao M, Li M, Duan P F, et al. Insights into the relationship between colony formation and extracellular polymeric substances (EPS) composition of the cyanobacterium Microcystis spp.[J]. Harmful Algae, 2019, 83: 34-41. DOI:10.1016/j.hal.2019.02.006 |
| [19] |
孙伟, 巩小丽, 陈煜, 等. 太湖藻源溶解性有机质光化学降解研究[J]. 湖泊科学, 2018, 30(1): 91-101. Sun W, Gong X L, Chen Y, et al. Photochemical degradation of the algae-derived dissolved organic matter in Lake Taihu[J]. Journal of Lake Sciences, 2018, 30(1): 91-101. |
| [20] |
许明, 刘伟京, 白永刚, 等. 太湖蓝藻水华期可溶有机物的生物降解[J]. 中国环境科学, 2018, 38(9): 3494-3501. Xu M, Liu W J, Bai Y G, et al. Biodegradation of dissolved organic matter in Lake Taihu during cyanobacterial blooms[J]. China Environmental Science, 2018, 38(9): 3494-3501. |
| [21] | Lalonde K, Mucci A, Ouellet A, et al. Preservation of organic matter in sediments promoted by iron[J]. Nature, 2012, 483(7388): 198-200. DOI:10.1038/nature10855 |
| [22] | Lee D, Kwon M, Ahn Y, et al. Characteristics of intracellular algogenic organic matter and its reactivity with hydroxyl radicals[J]. Water Research, 2018, 144: 13-25. DOI:10.1016/j.watres.2018.06.069 |
| [23] | Vecchio R D, Blough N V. Photobleaching of chromophoric dissolved organic matter in natural waters: kinetics and modeling[J]. Marine Chemistry, 2002, 78(4): 231-253. DOI:10.1016/S0304-4203(02)00036-1 |
| [24] | Keith D J, Yoder J A, Freeman S A. Spatial and temporal distribution of coloured dissolved organic matter (CDOM) in Narragansett Bay, Rhode Island: implications for phytoplankton in coastal waters[J]. Estuarine, Coastal and Shelf Science, 2002, 55(5): 705-717. DOI:10.1006/ecss.2001.0922 |
| [25] | McKnight D M, Boyer E W, Westerhoff P K, et al. Spectrofluorometric characterization of dissolved organic matter for indication of precursor organic material and aromaticity[J]. Limnology and Oceanography, 2001, 46(1): 38-48. DOI:10.4319/lo.2001.46.1.0038 |
| [26] | Minero C, Lauri V, Falletti G, et al. Spectrophotometric characterisation of surface lakewater samples: implications for the quantification of nitrate and the properties of dissolved organic matter[J]. Annali Di Chimica, 2007, 97(10): 1107-1116. DOI:10.1002/adic.200790094 |
| [27] | Helms J R, Stubbins A, Ritchie J D, et al. Absorption spectral slopes and slope ratios as indicators of molecular weight, source, and photobleaching of chromophoric dissolved organic matter[J]. Limnology and Oceanography, 2008, 53(3): 955-969. DOI:10.4319/lo.2008.53.3.0955 |
| [28] | Zsolnay A, Baigar E, Jimenez M, et al. Differentiating with fluorescence spectroscopy the sources of dissolved organic matter in soils subjected to drying[J]. Chemosphere, 1999, 38(1): 45-50. DOI:10.1016/S0045-6535(98)00166-0 |
| [29] | Kida M, Kojima T, Tanabe Y, et al. Origin, distributions, and environmental significance of ubiquitous humic-like fluorophores in Antarctic lakes and streams[J]. Water Research, 2019, 163. DOI:10.1016/j.watres.2019.114901 |
| [30] | Cawley K M, Ding Y, Fourqurean J, et al. Characterising the sources and fate of dissolved organic matter in Shark Bay, Australia: a preliminary study using optical properties and stable carbon isotopes[J]. Marine and Freshwater Research, 2012, 63(11): 1098-1107. DOI:10.1071/MF12028 |
| [31] | Chen M L, Kim J H, Nam S I, et al. Production of fluorescent dissolved organic matter in Arctic Ocean sediments[J]. Scientific Reports, 2016, 6(1). DOI:10.1038/srep39213 |
| [32] | Lapierre J F, Del Giorgio P A. Partial coupling and differential regulation of biologically and photochemically labile dissolved organic carbon across boreal aquatic networks[J]. Biogeosciences, 2014, 11(20): 5969-5985. DOI:10.5194/bg-11-5969-2014 |
| [33] | Wünsch U J, Murphy K R, Stedmon C A. The one-sample PARAFAC approach reveals molecular size distributions of fluorescent components in dissolved organic matter[J]. Environmental Science & Technology, 2017, 51(20): 11900-11908. |
| [34] | Fox B G, Thorn R M S, Anesio A M, et al. The in situ bacterial production of fluorescent organic matter; an investigation at a species level[J]. Water Research, 2017, 125: 350-359. DOI:10.1016/j.watres.2017.08.040 |
| [35] | Guillemette F, Del Giorgio P A. Simultaneous consumption and production of fluorescent dissolved organic matter by lake bacterioplankton[J]. Environmental Microbiology, 2012, 14(6): 1432-1443. DOI:10.1111/j.1462-2920.2012.02728.x |
| [36] | Moinet G Y K, Hunt J E, Kirschbaum M U F, et al. The temperature sensitivity of soil organic matter decomposition is constrained by microbial access to substrates[J]. Soil Biology and Biochemistry, 2018, 116: 333-339. DOI:10.1016/j.soilbio.2017.10.031 |
| [37] |
曾野, 朱金格, 王艳平, 等. 东太湖水温变化与水-沉积物界面热通量初探[J]. 湖泊科学, 2018, 30(6): 1599-1609. Zeng Y, Zhu J G, Wang Y P, et al. Changes of water temperature and heat flux at water-sediment interface, East Lake Taihu[J]. Journal of Lake Sciences, 2018, 30(6): 1599-1609. |
| [38] | Zhang Y L, Van Dijk M A, Liu M L, et al. The contribution of phytoplankton degradation to chromophoric dissolved organic matter (CDOM) in eutrophic shallow lakes: field and experimental evidence[J]. Water Research, 2009, 43(18): 4685-4697. DOI:10.1016/j.watres.2009.07.024 |
| [39] | Du Y X, Zhang Q Y, Liu Z W, et al. Composition of dissolved organic matter controls interactions with La and Al ions: implications for phosphorus immobilization in eutrophic lakes[J]. Environmental Pollution, 2019, 248: 36-47. DOI:10.1016/j.envpol.2019.02.002 |
| [40] | Sowers T D, Adhikari D, Wang J, et al. Spatial associations and chemical composition of organic carbon sequestered in Fe, Ca, and organic carbon ternary systems[J]. Environmental Science & Technology, 2018, 52(12): 6936-6944. |
| [41] | Coward E K, Ohno T, Sparks D L. Direct evidence for temporal molecular fractionation of dissolved organic matter at the iron oxyhydroxide interface[J]. Environmental Science & Technology, 2019, 53(2): 642-650. |
| [42] | Jiao N Z, Herndl G J, Hansell D A, et al. Microbial production of recalcitrant dissolved organic matter: long-term carbon storage in the global ocean[J]. Nature Reviews Microbiology, 2010, 8(8): 593-599. DOI:10.1038/nrmicro2386 |