 2020, Vol. 41
2020, Vol. 41



烹饪油烟是指食物烹饪和食品生产加工过程中挥发的油脂、有机质及热氧化和热裂解产生的混合物[1~3], 具有显著的人体健康危害.然而, 大部分烹饪油烟由于缺乏实用高效的油烟气净化技术, 未经处理直排至室外, 成为引起城市大气污染的重要排放源之一[4].烹饪油烟中含有多种挥发性有机物, 如烃类、醛类和酮类等[5, 6].其中, 醛类污染物是最常见的污染物之一[7], 在煎炸过程中其贡献的排放量大于60%[8].而醛类污染物的排放又以甲醛和乙醛为主[9~13].甲醛在我国优先控制致癌污染物名单中列居第二位[14], 短期接触甲醛会导致人的嗅觉、肺功能异常, 而长期接触可引起各种慢性呼吸道疾病、中枢神经系统紊乱、记忆力和智力下降等[15].因此, 研究烹饪油烟中甲醛的净化技术具有重要意义.
甲醛去除方法主要有催化法[16]、吸附法[17]和等离子体净化法[18]等.这些去除方法取得了一定的研究进展, 例如:以氧化锰为催化剂对甲醛进行净化, 24 h甲醛的去除率达到92.3%[19].以氮化硼为原料制成吸附剂去除甲醛, 试验运行4次后, 甲醛去除率可达到8.5 mg·g-1[20].然而, 这些方法存在成本高、体积大和去除效率低的问题[21, 22].如催化净化法去除成本较高, 吸附法存在吸附能力随时间减弱和更换吸附剂的问题[23].因此, 开发一种成本低廉和操作简单的甲醛净化技术是目前亟待解决的问题.
微生物法处理污染物已应用在环境治理各个方面.微生物法废气净化技术是利用微生物废气中的有机组分作为其生命活动的能源或养分, 经代谢分解, 将其转化为简单的无机物或细胞组成物质.在生物过滤器中, 增湿后的有机废气与附着在过滤材料表面的微生物接触时作为能源和营养物质被吸收, 在微生物代谢过程中被分解为无害的无机物.因此, 生物法具有成本低、安全性好和无二次污染等优点, 已成为世界工业废气净化研究的热点之一[24, 25].研究微生物法处理醛类VOCs可能为烹饪油烟VOCs净化技术提供一种安全、高效和经济的解决方案.有研究筛选出针对降解油烟中的脂类、油酸、甲苯和己醛的不同菌种, 进行纯种培养后的生物填料塔的降解能力可稳定在85%以上[26, 27].然而目前利用生物法去除烹饪油烟中甲醛的研究鲜见报道.在现有研究中, 优势甲醛降解菌的种类主要包括从活性污泥或土壤等介质驯化分离得到的假单胞菌属和甲基营养菌属等[28, 29].由于这些菌株缺少耐油性能的相关研究, 难以应用于烹饪油烟中的醛类降解.
本研究从油烟冷凝液中筛选分离得到1株耐油的甲醛降解菌.与其他的甲醛降解菌相比, 它具有非常明确的耐油性能;并且通过对其进行胞外代谢产物的分析, 初步推断了菌株降解甲醛的可能的代谢途径, 以期为烹饪油烟的生物净化法提供了一定的试验基础和理论依据.
1 材料与方法 1.1 试验材料 1.1.1 样品采集本试验油烟冷凝液来源于北京工商大学西区食堂.样品采集后, 置于4℃冰箱中待后续试验使用.
1.1.2 培养基基础培养基(g·L-1):蛋白胨10.0、琼脂15.0、NaCl 5.0、KH2PO4 0.45和Na2HPO4 0.47;痕量金属溶液1 mL·L-1;pH 7.00±0.2, 121℃高压灭菌40 min.
痕量金属溶液(g·L-1):FeSO4·7H2O 0.55、ZnSO4·7H2O 0.23、MnSO4·7H2O 0.34、Co(NO3)2·6H2O 0.075、CuSO4·5H2O 0.047和(NH4)6MO7O24·4H2O 0.025.
无碳培养基(g·L-1):氯化铵1.77、氯化钠5、磷酸二氢钾0.45、磷酸氢二钠0.47、氯化钙0.02和硫酸镁0.25;痕量金属溶液1 mL·L-1;原始pH为6.60, 并在121℃下灭菌
耐油菌的筛选培养基:在灭菌后的基本培养基中加入油烟冷凝液.
复筛培养基:在灭菌后的无碳培养基中加入适当浓度甲醛溶液.
1.1.3 主要试剂和仪器NH4Cl、ZnSO4·7 H2O、FeSO4·7H2O、MnSO4·7H2O、Co(NO3)2·6 H2O、CuSO4·5 H2O、KH2PO4、Na2HPO4、MgSO4·7 H2O、NaCl、LB肉汤和蛋白胨等试剂购自北京半夏科技发展有限公司;甲醛、乙酰和丙酮等试剂购自北京伊诺凯有限公司;BSTFA(with 1% TMCS, 体积比)购自美国REGIS Technologies;L-2-氯苯丙氨酸购自上海恒柏生物科技;饱和脂肪酸甲酯购自Dr. Ehrenstorfer;甲氧铵盐购自东京化成工业株式会社.
分析天平, 北京赛多利科学仪器有限公司;生化培养箱, 上海申贤恒温设备厂;紫外可见分光光度计(UV-2550), 日本岛津制作所;高压蒸汽灭菌器, Panasonic公司;超净操作台, 哈尔滨市东联电子技术开发有限公司;低温冷冻离心机, Sigma公司;PCR仪, TECHNE公司;智能冰箱, SAMSUNG公司;气相色谱(7890A), Agilent公司;质谱仪(PEGASUS HT), LECO;超低温冰箱(Forma 900 series), Thermo Fisher Scientific;研磨仪(JXFSTPRP-24), 上海净信科技有限公司.
1.2 试验方法 1.2.1 菌株分离和纯化 1.2.1.1 菌株获取取适量油烟冷凝液于洗净的烧杯中, 滤膜封口后在超声清洗机中超声2 h, 取1%的油烟冷凝液加入到100 mL灭菌后的基础培养基中培养, 从摇瓶中取100 μL菌液涂布至营养琼脂的平板中分离纯化.
1.2.1.2 菌株驯化培养取分离纯化后的菌液按照5%的体积比加到灭菌的LB肉汤中, 摇瓶培养制成种子液, 在基础培养基中加入2.5 mg·L-1的甲醛, 置于170 r·min-1、30℃的摇床中培养24 h.之后, 将5 mL的混合物接种到新的100 mL基础培养基中培养, 并加入5 mg·L-1的甲醛置于相同条件培养24 h.以此类推, 甲醛浓度从2.5 mg·L-1升高至20 mg·L-1进行驯化培养.富集结束后, 使用稀释涂布法和平板划线法筛选出能以甲醛为碳源生存的单菌落, 并分离纯化.
1.2.2 菌株鉴定 1.2.2.1 菌株的形态学观察将分离得到的菌株在平板上涂布后30℃培养2 d, 观察菌落形态.通过电子显微镜和扫描电镜观察细菌的细胞形态及菌体大小.
扫描电镜样品制备:取培养至稳定期的菌液2 mL, 8 000 r·min-1离心5 min, 弃上清, 加入40倍菌体体积的2.5%戊二醛固定液, 置于4℃冰箱中固定2.5 h, 用磷酸盐缓冲液冲洗3遍, 依次用50%、70%、90%和100%乙醇进行梯度脱水, 每次脱水时间为5 min.脱水结束后, 用乙醇-叔丁醇溶液(1:1, 体积比, 下同)置换20 min, 最后用100%叔丁醇置换两次, 每次20 min.置换后将样品进行冷冻干燥处理, 干燥后的样品进行离子溅射仪喷镀后, 扫描电子显微镜观察和拍照.
1.2.2.2 菌株的生理生化试验细菌的生理生化试验根据文献[30]进行革兰氏染色、过氧化氢酶、甲基红、吲哚、V-P反应和淀粉水解等试验测定.
1.2.2.3 16S rDNA鉴定及系统发育分析鉴定菌株划线接种于营养琼脂的平板中, 在30℃霉菌培养箱中培养至对数期, 对该菌株16S rDNA基因序列分析.利用DNA提取试剂盒提取总DNA作为PCR模板, 选用16S菌保守序列进行PCR扩增.16S rDNA扩增的引物为27F: AGAG TTTGATCMTGGCTCAG和1492R: TACGGYTACCTT GTTACGACTT.PCR程序为:95℃预变性5 min, 95℃变性30 s, 55℃退火30 s, 72℃延伸1 min, 循环35次;72℃延伸10 min, 12℃保存.得到750 bp的片段, 测序得到菌株XF-1的16S rDNA全序列, 并将序列上传到NCBI基因库进行BLAST比对.利用MEGA 5.05软件构造系统发育进化树.
1.2.3 菌株XF-1的降解性能 1.2.3.1 不同培养条件对菌株降解能力的影响及降解过程的研究取活化后的XF-1菌悬液分别按照不同接种量(8%、10%、12%、15%、18%和20%)接种至甲醛初始浓度为20 mg·L-1的培养基中.每个梯度设置3个平行, 以空白培养基为对照.将摇瓶置于转速为170 r·min-1、温度为30℃恒温摇床中培养, 定时取样测定培养基中甲醛浓度、D600[此处的D600代表溶液在波长为600 nm处的吸光值(optical density)]和pH.配制含300 mg·L-1甲醛的培养基, 并进行灭菌.冷却后, 取对数期的菌悬液以最佳接种量加入其中, 设置不同培养温度(25、30、35和40℃)和pH(5.0、6.0、7.0和8.0), 将摇瓶置于转速为170 r·min-1进行培养.每组做3个平行, 以空白培养基为对照.培养至一定时间取样并测定培养基中的甲醛浓度.
1.2.3.2 菌株对不同浓度的甲醛降解为了研究菌株在不同甲醛浓度下的降解能力, 将甲醛浓度梯度分别设置为100、200、300、500和800 mg·L-1.每个浓度梯度做3个平行, 以空白培养基为对照.培养基中加入的微生物量为10%(体积分数).对照组培养基和不同浓度甲醛的培养基同时置于30℃、170 r·min-1的摇床中培养.定时取样检测甲醛浓度, 所有取样均在无菌操作台里操作.
1.2.3.3 菌株对甲醛的最大耐受浓度按照10%的接种量分别将种子液接种于甲醛浓度为100、300、800、1 000、1 500、2 000和3 000 mg·L-1的培养基中, 每个浓度梯度设置2个平行, 以空白不加菌的培养基为对照.相同条件培养一段时间后对样品中的菌液稀释涂布进行活菌计数.
1.2.4 菌株XF-1的特异性 1.2.4.1 菌株XF-1的耐油特性将待测定的菌株活化后的种子液以10%的接种量接入至每100 mL含食用油(已灭菌)分别为1、4、8、10、20、30、50、60、80和90 g的无碳培养基中, 另将已冷却的接种环伸入菌液中蘸取一环菌液加到含有100 g食用油(已灭菌)的摇瓶中.每个浓度梯度样品设置2个平行, 以空白不加菌的培养基为对照.培养基同时置于30℃、170 r·min-1的摇床中培养, 培养一段时间后检测各摇瓶的细菌生长情况.食用油质量浓度小于50%的样品采用重悬法检测D600;食用油质量浓度大于50%的样品, 为避免干扰D600检测结果, 故采用稀释涂布法进行活菌计数检测细菌生长情况.
1.2.4.2 菌株降解甲醛的特异性为了验证从油烟冷凝液中驯化得到的解淀粉芽孢杆菌XF-1具有可降解甲醛的特异性, 与其他解淀粉芽孢杆菌进行对比试验.从中国微生物菌种保藏管理委员会普通微生物中心(CGMCC)购入两株解淀粉芽孢杆菌, 编号分别为1.871 6和1.871 9进行甲醛降解试验, 研究相同菌种的菌株对甲醛的降解能力.甲醛浓度设置为100 mg·L-1, 每种菌株设置3个平行, 以空白培养基为对照.对照组与试验组同时置于30℃、170 r·min-1的摇床中培养.定时取样检测微生物的生长情况和甲醛浓度的变化, 所有取样均在无菌操作台里操作.
1.2.5 分析方法 1.2.5.1 甲醛浓度的测定参照GB/T 13197-1991, 取适量待测样品加入10 mL具塞刻度试管, 用蒸馏水定容至10 mL刻度线后, 吸取2 mL乙酰丙酮溶液加入该试管, 混匀, 60℃水浴15 min, 室温冷却1 h后, 以蒸馏水为参比检测在波长413 nm处的吸光度.并建立如图 1所示的甲醛标准曲线, 线性范围为0.2~3.5 mg·L-1, 线性方程为y=0.220 73x+0.000 53, 相关度R2=0.999 97.

|
图 1 甲醛标准曲线 Fig. 1 Formaldehyde standard curve |
取44支盛有5 mL LB肉汤的试管, 灭菌后备用.其中33支接种0.175 mL已培养12 h的菌液(使初始D600达到0.01).另11支作为对照.将44支试管置于恒温摇床中振荡培养, 温度设置为30℃、转速170 r·min-1.每隔一段时间取样检测其在600 nm处的吸光度值, 得到微生物生长曲线.
根据菌株在不同时间的生长数据, 选择修正SGompertz模型[31, 32]拟合降解菌的生长曲线.从而通过模型参数计算方程[公式(1)]推导得到最大比生长速率U[公式(2)].最终采用R2对建立模型进行评价.
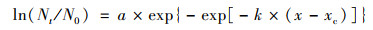
|
(1) |
由此可以得到U的计算公式:
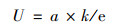
|
(2) |
式中, t为时间, Nt和N0分别表示在时间t时和初始时间时的微生物数量, xc为达到最大生长速率的时间, e=2.718 2.
1.2.5.3 重悬法检测D600为防止食用油干扰细胞生长量的测定, 采用以下方法排除干扰.取4 mL样品至离心管中, 12 000 r·min-1、4℃离心5 min倾斜倒去上清液, 收集湿菌体并用无菌水洗涤两次, 最后将湿菌体转入4 mL无菌水中涡旋形成均匀稳定的悬浮液, 以蒸馏水为参比, 在分光光度计上于600 nm检测吸光度.
1.2.5.4 胞外代谢产物测定代谢物提取:取样本300 μL于1.5 mL EP管中, 加入300 μL预冷甲醇提取液, 再加入10 μL L-2-氯苯丙氨酸, 涡旋30 s, 然后将样品置于冰水浴中超声10 min, 4℃离心, 12 000 r·min-1离心15 min, 小心移取100 μL上清液于1.5 mL EP管中, 每个样本各取50 μL混合成QC样本;在真空浓缩器中干燥提取物并将干燥后的代谢物加入60 μL甲氧胺盐试剂(甲氧胺盐酸盐, 溶于吡啶20 mg·mL-1), 轻轻混匀后, 放入烘箱中80℃孵育30 min, 再向每个样品中加入80 μL BSTFA (含有1% TMCS, 体积比), 70℃孵育1.5 h, 冷却至室温后向混合的样本中加入5 μL FAMEs(溶于氯仿), 随机顺序上机检测.
分析方法:使用Agilent 7890气相色谱仪-飞行时间质谱仪进行GC-TOF-MS分析, 色谱柱:DB-5MS毛细管柱(30 m× 250 μm×0.25 μm, J & W Scientific, Folsom, CA, USA).以氦气为载体气体, 前段进气吹扫流量为3 mL·min-1, 进样量为1 μL, 柱流速为1 mL·min-1.升温程序:初始温度50℃保持1 min, 以10℃·min-1升至310℃保持8 min.前进样口、传输线、离子源温度分别为280、280和250℃, 电子能量:70 eV, 质谱数据在全扫描模式下获得, m/z范围为50~500, 溶剂延迟为6.30 min.
数据处理:使用ChromaTOF软件(V 4.3x, LECO)对质谱数据进行了峰提取、基线矫正、解卷积、峰积分和峰对齐等分析.对物质定性工作中, 使用了LECO-Fiehn Rtx5数据库, 包括质谱匹配及保留时间指数匹配.
2 结果与讨论 2.1 菌种的鉴定 2.1.1 菌株的形态特征观察筛选分离后, 对得到的可降解甲醛的菌株XF-1进行形态特征观察.如图 2所示, 可以观察到菌株XF-1呈清晰的单菌落, 菌株大小2 mm, 圆形, 边缘规则, 凸起, 不透明, 乳白色, 表面光滑.

|
(a)平板; (b)电子显微镜; (c)扫描电镜 图 2 菌株在平板、电子显微镜下观察和扫描电镜下观察的形态 Fig. 2 Plate, electron microscope, and transmission electron microscope photos of the XF-1 strain |
通过电子显微镜和扫描电镜进行观察, 该菌株呈短杆状, 有芽孢产生, 为革兰氏染色阳性菌.
2.1.2 菌株的生理生化结果及分子生物学鉴定菌株XF-1的5个生理生化试验结果显示(表 1), 为革兰氏阳性菌, 吲哚试验阴性, V-P试验阳性, 淀粉水解试验阳性, 过氧化氢酶试验阳性.对菌株的DNA进行PCR扩增及16S rDNA基因鉴定后, 根据BLAST结果, 表明该菌株与Bacillus sp.亲源关系最近, 同源性达99%以上.在NCBI上匹配与菌株XF-1所测序列较高相似性的序列, 在MEGA 5.05软件中绘制系统发育树图(图 3).结合菌株的形态观察结果及生理生化试验结果, 确定该菌株鉴定为解淀粉芽孢杆菌.
|
|
表 1 生理生化特性鉴定结果1) Table 1 Identification results of the physiological and biochemical characteristics |

|
参与比对序列的GenBank登录号列于括号中, 标尺表示序列差异的分支长度 图 3 菌株XF-1的系统发育进化树 Fig. 3 Phylogenetic tree derived from 16S rDNA sequences of the XF-1 strain |
菌株XF-1的生长曲线如图 4所示.从中可知, 菌株在6~10 h达到最大生长率速度, 菌株的D600值最高达1.25 ± 0.036.运用Origin软件, 选用SGompertz模型拟合菌株的生长曲线(R2=0.985), 如图 4所示.根据1.2.5.2节中提到的公式(1)计算得到, 菌株达到最大生长速率所需时间xc为6.04 h;接着由[公式(2)]计算可得到菌株的最大比增长速率:

|
图 4 SGompertz模型拟合的Bacillus sp. XF-1的生长曲线 Fig. 4 Growth curve of the XF-1 strain and SGompertz fit of the XF-1 strain |
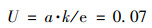
|
为了确定XF-1菌株对甲醛的最佳降解效果, 分别考察不同接种量、温度和pH对菌株降解甲醛性能的影响, 以获得最佳降解条件参数.
2.2.1 接种量分别以质量分数为8%、10%、12%、15%、18%和20%的菌液接种量接种活化后的XF-1菌悬液至含20 mg·L-1甲醛的培养基中, 结果如图 5所示, 甲醛在培养基中的浓度随着时间延长而不断降低.其中, 10%接种量的甲醛浓度曲线为实线, 其余接种量为虚线.从图 5中可知, 随着接种量的增大, 甲醛降解速率随之增加, 接种量几乎与甲醛降解成正相关.在6 h以后, 除8%外, 其余接种量条件下的甲醛降解率没有显著差异.即在接种量达10%以后, 降解率不再随接种量的增大而显著增加.

|
图 5 不同接种量下, 菌株XF-1降解甲醛时甲醛浓度随时间的变化 Fig. 5 Change in the formaldehyde concentration over time when XF-1 degrades formaldehyde under different inoculation conditions |
由图 6(a)可得:当接种量为18%和20%时, 虽然降解速率明显在4 h高于其他接种量, 但由于细菌数较大使得彼此之间的竞争激烈, 其衰亡时间也较其他接种量提前, 而10%、12%和15%的接种量所对应的细菌生长量变化较为一致, 8%接种量的降解效果比10%~ 20%接种量的差, 因而可认为10%为最佳接种量.图 6(b)为XF-1菌降解甲醛时pH随时间的变化, 从图 5和图 6可知, 随着时间的变化, pH不断增大, 其甲醛浓度逐渐减小, 细菌生长量呈先升高后降低趋势, 但各试验组间的pH变化相差不大.综合不同接种量条件下菌株XF-1降解甲醛的甲醛浓度变化、D600和pH检测结果分析, 可判定10%为最佳接种量.

|
图 6 不同接种量下, 菌株XF-1降解甲醛时D600与pH随时间的变化 Fig. 6 Change of D600 and pH over time when the bacteria XF-1 degraded formaldehyde at different inoculation quantities |
选取4个pH梯度, 通过1 mol·L-1的HCl和20%的NaOH将初始pH分别调为5.0、6.0、7.0和8.0.以300 mg·L-1为甲醛初始浓度研究pH对菌株XF-1降解甲醛的影响, 结果如图 7所示.尽管菌株在pH为5~8时都可以生长, 但pH等于6时的降解效果最好, 降解率可达94.61%, 当初始pH大于6时, 降解能力减弱.微生物在降解污染物的过程中, 微生物活性是影响降解率的关键因素.本研究中, 菌株XF-1生长的最佳pH为6, 在微生物降解VOCs的过程中, 酸碱环境会改变底物和酶的带电状态, 进而影响酶和底物的特异性结合.当微生物处于最适pH范围内会增大活性, 达到最佳的状态[33].

|
图 7 甲醛初始浓度为300 mg·L-1时, 菌株XF-1在不同pH下对甲醛降解率的变化 Fig. 7 Change in the formaldehyde degradation rate of the XF-1 strain at different pH levels when the initial concentration of formaldehyde was 300 mg·L-1 |
以300 mg·L-1为甲醛初始浓度, 分别在不同温度25、30、35和40℃培养时, 温度对菌株XF-1降解甲醛的影响如图 8所示.在144 h时, 温度为30℃, 菌株降解甲醛能力最佳, 降解率为76.5%, 而在40℃时, 甲醛降解率仅为49.62%.菌株XF-1最佳生长温度为30℃左右, 这与张娟等[34]的研究发现解淀粉芽孢杆菌的培养温度一致.

|
图 8 甲醛初始浓度为300 mg·L-1时, 菌株XF-1在不同温度下对甲醛降解率的变化 Fig. 8 Change in the formaldehyde degradation rate of the XF-1 strain at different temperature when the initial concentration of formaldehydes was 300 mg·L-1 |
菌株XF-1对不同浓度甲醛的降解能力如图 9所示, 当甲醛浓度分别为100、200、300、500和800 mg·L-1时, 菌株XF-1在48 h对100 mg·L-1甲醛的降解效率可达99.63%;在96 h, 菌株XF-1对300 mg·L-1甲醛的降解效率可达89.46%, 对800 mg·L-1甲醛的降解效率可达到73.01%.菌株XF-1对甲醛有良好的降解能力.

|
图 9 菌株XF-1对不同浓度甲醛的降解能力 Fig. 9 Ability of the XF-1 strain to degrade formaldehyde at different concentrations |
分别对甲醛浓度为100、300、800、1 000、1 500、2 000和3 000 mg·L-1的培养基中的菌液稀释涂布进行活菌计数.在培养一段时间后, 对培养基中的菌液稀释涂布计数, 发现其细菌数呈下降趋势, 表明甲醛对菌株XF-1存在一定的生长抑制.同时, 如图 10所示, 在50 h, 菌株XF-1对甲醛的去除效率开始随着时间变化趋于平缓, 表明菌株对该浓度甲醛的降解能力接近最大, 故对培养至50 h的菌液稀释涂布进行活菌计数进而研究其对甲醛的最大耐受浓度.本试验发现, 当培养至50 h时, 甲醛浓度大于1.5 g·L-1的菌液在平板上没有长出菌落, 而其他甲醛浓度小于1.5 g·L-1的菌液经过稀释涂布在平板上均有菌落长出(图 10), 最后得到菌株XF-1的最大耐受甲醛浓度为1.5 g·L-1.

|
图 10 菌株XF-1在不同甲醛浓度培养时细菌数随时间的变化 Fig. 10 Changes in bacterial number over time in the XF-1 strain culture at different formaldehyde concentrations |
现有研究中甲醛降解菌对不同浓度的甲醛的降解效率和最大耐受浓度存在一定的差异, 如张明等[35]以甲醛为唯一碳源, 在小型试验填料塔内长满生物膜的陶粒上分离纯化出两株甲醛降解菌P1和Q1, 在含甲醛浓度300 mg·L-1的培养基中, 菌株P1在44 h内的甲醛降解率为92.8%, 菌株Q1在28 h内的甲醛降解率为97.9%.钟卫鸿等[36]从污水处理厂收集的土壤中分离到一株能降解甲醛的菌株, 其对甲醛的最大耐受浓度为1.2 g·L-1.结合试验数据与上述结果进行比较, 表明菌株XF-1可以耐受较高浓度的甲醛并且具有良好的甲醛降解能力.
2.4 菌株XF-1的特异性 2.4.1 3株解淀粉芽孢杆菌对甲醛的降解从中国微生物菌种保藏管理委员会普通微生物中心(CGMCC)购入两株编号分别为1.871 6和1.871 9的解淀粉芽孢杆菌, 将其和菌株XF-1以相同条件培养进行甲醛降解试验, 3株解淀粉芽孢杆菌对甲醛的降解效果如图 11所示, 从中可知, 虽然菌株皆为解淀粉芽孢杆菌, 但从油烟冷凝液中筛选、驯化后的菌株XF-1与其他两株菌相比, 对甲醛具有一定的耐受性和更好的降解效果.编号为1.871 6和1.871 9的解淀粉芽孢杆菌在6 h时, D600从最初的0.33下降至0.05, 表示菌株由于甲醛的毒性抑制, 菌体大部分失活死亡.

|
图 11 3株解淀粉芽孢杆菌甲醛浓度和D600随时间的变化 Fig. 11 Changes in formaldehyde concentration and D600 of three Bacillus amyloliquefaciens sp. |
于食用油质量分别为1%、4%、8%、10%、20%、30%和50%的无碳培养基中加入10%的菌液, 培养一段时间后采用重悬法检测D600, 如图 12所示, 含1%~50%质量分数食用油的样品D600均有明显增高.为防止较大食用油含量影响重悬法检测D600的准确性, 当食用油质量分数大于50%时, 利用活菌计数法检测细菌生长情况.通过试验发现, 含60%、80%和90%质量分数食用油的样品经过48 h培养后, 60%和80%的样品细菌数有明显升高的趋势, 90%细菌数几乎没有变化, 但是将菌株接种在含100%质量分数食用油的样品中并培养48 h后, 对样品稀释涂布发现没有菌落长出.试验表明1%~80%质量分数的食用油条件下, 菌株可以良好生长, 最大可以耐受90%质量分数, 即900 g·L-1的食用油.表明菌株XF-1可以耐受高油环境, 可以为生物法降解烹饪油烟中的甲醛提供优良的菌株资源.

|
图 12 不同质量分数食用油下, 菌株D600或细菌数随时间的变化 Fig. 12 Variation of D600 or bacteria number with time at different mass fraction of edible oil |
利用GC-TOF-MS对胞外代谢产物进行检测分析, 共筛选得到66种代谢物.将6 h和24 h样品中测得的代谢产物进行比较.随着时间的变化, 其中有26种代谢物上调, 包括2-羟基丁酸、戊二酸和核糖等;33种代谢物下调, 如对-羟基苯乙酸、苯丙氨酸、丙氨酸(如图 13).另外有2种代谢物只出现在6 h的样品中, 分别是苯乙酸和果糖;5种代谢物单独存在于24 h的样品中, 分别为乙醇胺、甘油、核糖酸、葡萄糖内酯和色醇.

|
列举相对变化分别为前十的代谢物 图 13 在不同生长阶段代谢物浓度的相对变化 Fig. 13 Relative changes in metabolite concentrations at different growth stages |
有研究发现生物法降解甲醛过程中甲醛代谢途径主要分为同化与异化两种途径[37].同化作用主要包括丝氨酸途径(Serine)和核酮糖单磷酸途径(RuMP).丝氨酸循环途径是通过C3化合物酸化生成苹果酸, 苹果酸被再次分解生成甘氨酸.而RuMP途径是存在于许多甲基营养菌中的甲醇、甲酸、甲胺等一碳化合物的同化途径, 磷酸己酮糖合成酶(HPS)和磷酸果糖异构酶(PHI)是RuMP代谢途径的关键酶, 而这两种酶普遍存在于甲基营养菌中[38].甲醛在整个微生物氧化过程中处于一个中心位置, 作为大多数甲基基质氧化的一个普遍的中间体.在微生物细胞内, 游离甲醛首先通过与各种辅因子的结合形成加合物而脱毒[39].
异化途径主要包括四氢叶酸途径、四氢甲烷蝶呤途径和谷胱甘肽途径[37], 在这3种异化途径过程中, 这些反应产生的甲醛加合物主要被甲醛脱氢酶催化氧化为甲酸, 甲酸继而在甲酸脱氢酶的作用下被微生物利用生成CO2和再生辅因子, 同时提供能量供微生物生长, 其代谢途径产生的甲酸可能会导致pH降低[40], 在研究不同接种量对菌株降解能力的影响试验中, 通过检测pH指标发现, 试验组随着时间变化, pH逐渐上升[图 6(b)], 所以推测XF-1菌株降解甲醛的途径可能不是通过异化途径进行而是其他代谢途径.
通过对不同时间的胞外代谢产物分析发现, 6 h样品中检测得到的代谢物果糖并未在24 h对应的样品中检测到, 且24 h样品主要出现的代谢物包含甘油和核糖.将这些差异代谢物与前人关于甲醛代谢途径的研究进行比对发现, 本试验结果与RuMP途径呈现部分吻合.在RuMP代谢途径中, 6-磷酸果糖(F6P)是在同化作用第一阶段出现的, 在第二阶段会断裂形成3-磷酸甘油醛(GAP)和磷酸二羟丙酮(DHAP), 而5-磷酸核糖(Ri5P)是在第三阶段3-磷酸甘油醛(GAP)通过醛羧酶催化生成的, 这一现象恰与本试验结果相一致.目前芽孢杆菌作为非甲基营养菌, 其同化甲醛的机制还未研究全面, 但是枯草芽孢杆菌中HPS和PHI在非甲基营养生物中的生理作用已得到证实[41], 程琴等[42]利用激光光镊拉曼光谱系统研究不同浓度甲醛胁迫枯草芽孢杆菌萌发后2 h内孢子的响应过程, 发现芽孢杆菌的营养细胞和萌发后孢子对一定浓度的甲醛有耐受能力并且表明营养细胞在甲醛的诱导下, 可以通过RuMP途径代谢甲醛进行脱毒.综上, 初步推测菌株XF-1的甲醛降解途径可能为同化途径——核酮糖单磷酸途径.
3 结论(1) 从油烟冷凝液中分离驯化得到1株可降解甲醛的菌株, 该菌具有良好的耐油性, 可在高浓度的油脂中存活, 最大可耐受900 g·L-1的食用油.通过生理生化以及分子生物学分析, 鉴定该菌株XF-1为解淀粉芽孢杆菌(Bacillus amyloliquefaciens sp.)
(2) 对菌株XF-1降解甲醛的最适条件进行了初步探索, 试验结果表明:菌株XF-1的适宜初始pH为6, 最佳接种量为10%, 适宜温度为30℃;对其生长曲线及生长动力学进行研究, 确定该菌的最佳活性时间(即菌株达到最大生长速率所需时间)为6.04 h, 这为后期菌株的实际应用提供了理论依据.
(3) 考察了不同初始浓度下菌株XF-1对甲醛的降解性能.结果表明:随着甲醛浓度的增大, 菌株完全降解甲醛的时间随之延长.当甲醛浓度为100 mg·L-1时, 菌株XF-1在48 h的降解效率可达99.63%;当甲醛浓度为300 mg·L-1时, 在96 h的降解率可达89.46%;当甲醛浓度为800 mg·L-1时, 菌株XF-1在96 h的降解率达到73.01%.另外, 菌株XF-1对甲醛的最大耐受浓度为1.5 g·L-1.
(4) 通过对菌株的胞外代谢物进行分析, 初步推测该菌降解甲醛的代谢途径可能为同化途径的核酮糖单磷酸途径.
(5) 综上, 菌株XF-1是性能良好的耐油甲醛降解菌, 该研究为烹饪油烟的生物净化法提供了一定的试验基础和理论依据.
| [1] |
何万清, 聂磊, 田刚, 等. 基于GC-MS的烹调油烟VOCs的组分研究[J]. 环境科学, 2013, 34(12): 4605-4611. He W Q, Nie L, Tian G, et al. Study on the chemical compositions of VOCs emitted by cooking oils based on GC-MS[J]. Environmental Science, 2013, 34(12): 4605-4611. |
| [2] | Xue Y B, Jiang Y, Jin S, et al. Association between cooking oil fume exposure and lung cancer among Chinese nonsmoking women:a meta-analysis[J]. OncoTargets and Therapy, 2016, 9: 2987-2992. |
| [3] | Yin Z H, Li H, Cui Z G, et al. Polymorphisms in pre-miRNA genes and cooking oil fume exposure as well as their interaction on the risk of lung cancer in a Chinese nonsmoking female population[J]. OncoTargets and Therapy, 2016, 9: 395-401. |
| [4] |
朱春, 李旻雯, 缪盈盈, 等. 城市烹饪油烟颗粒物排放特性分析[J]. 绿色建筑, 2014(5): 57-60, 71. Zhu C, Li M W, Miu Y Y, et al. Analysis of characters of particulate emissions generated from urban cooking fume[J]. Green Building, 2014(5): 57-60, 71. |
| [5] |
高雅琴, 王红丽, 许睿哲, 等. 餐饮源挥发性有机物组成及排放特征[J]. 环境科学, 2019, 40(4): 1627-1633. Gao Y Q, Wang H L, Xu R Z, et al. Characterization of volatile organic compounds from cooking emissions[J]. Environmental Science, 2019, 40(4): 1627-1633. |
| [6] | Yi H H, Huang Y H, Tang X L, et al. Mn-CeOx/MeOx(Ti, Al)/cordierite preparation with ultrasound-assisted for non-methane hydrocarbon removal from cooking oil fumes[J]. Ultrasonics Sonochemistry, 2019, 53: 126-133. DOI:10.1016/j.ultsonch.2018.12.043 |
| [7] | Peng C Y, Lan C H, Lin P C, et al. Effects of cooking method, cooking oil, and food type on aldehyde emissions in cooking oil fumes[J]. Journal of Hazardous Materials, 2017, 324: 160-167. DOI:10.1016/j.jhazmat.2016.10.045 |
| [8] | Klein F, Platt S M, Farren N J, et al. Characterization of gas-phase organics using proton transfer reaction time-of-flight mass spectrometry:Cooking emissions[J]. Environmental Science & Technology, 2016, 50(3): 1243-1250. |
| [9] | Wu M T, Lin P C, Pan C H, et al. Risk assessment of personal exposure to polycyclic aromatic hydrocarbons and aldehydes in three commercial cooking workplaces[J]. Scientific Reports, 2019, 9(1). DOI:10.1038/s41598-018-38082-5 |
| [10] | Cheng J H, Lee Y S, Chen K S. Carbonyl compounds in dining areas, kitchens and exhaust streams in restaurants with varying cooking methods in Kaohsiung, Taiwan[J]. Journal of Environmental Sciences, 2016, 41: 218-226. DOI:10.1016/j.jes.2015.06.006 |
| [11] | Alves C A, Evtyugina M, Cerqueira M, et al. Volatile organic compounds emitted by the stacks of restaurants[J]. Air Quality, Atmosphere & Health, 2015, 8(4): 401-412. |
| [12] |
程婧晨, 崔彤, 何万清, 等. 北京市典型餐饮企业油烟中醛酮类化合物污染特征[J]. 环境科学, 2015, 36(8): 2743-2749. Cheng J C, Cui T, He W Q, et al. Pollution characteristics of aldehydes and ketones compounds in the exhaust of Beijing typical restaurants[J]. Environmental Science, 2015, 36(8): 2743-2749. |
| [13] | Lin J M, Liou S J. Aliphatic aldehydes produced by heating Chinese cooking oils[J]. Bulletin of Environmental Contamination and Toxicology, 2000, 64(6): 817-824. DOI:10.1007/s0012800076 |
| [14] |
徐云, 金晶, 郑重, 等. 高活性高耐受甲醛降解菌株的分离鉴定及降解条件研究[J]. 环境科学, 2010, 31(10): 2481-2486. Xu Y, Jin J, Zheng Z, et al. Isolation, identification and conditions of bacterial strain capable to metabolize high concentrations of formaldehyde[J]. Environmental Science, 2010, 31(10): 2481-2486. |
| [15] |
冯永, 王茹, 陈丽梅. 固定沼泽红假单胞菌构建悬浮式生物反应器对室内甲醛的去除[J]. 环境工程学报, 2018, 12(4): 1071-1082. Feng Y, Wang R, Chen L M. Performance of suspension bioreactor with fixed Rhodopseudomonas palustris for removal of formaldehyde in indoor[J]. Chinese Journal of Environmental Engineering, 2018, 12(4): 1071-1082. |
| [16] | Bai B Y, Qiao Q, Li J H, et al. Progress in research on catalysts for catalytic oxidation of formaldehyde[J]. Chinese Journal of Catalysis, 2016, 37(1): 102-122. DOI:10.1016/S1872-2067(15)61007-5 |
| [17] |
蒋昕楠, 孔振凯, 王际童, 等. 高锰酸钾改性球形中孔炭的甲醛吸附性能[J]. 环境工程学报, 2018, 12(6): 1676-1682. Jiang X N, Kong Z K, Wang J T, et al. Formaldehyde adsorption performance of KMnO4 modified mesoporous carbon spheres[J]. Chinese Journal of Environmental Engineering, 2018, 12(6): 1676-1682. |
| [18] |
高丁丁, 蒙冕武, 谢君梁, 等. 改性活性炭纤维协同等离子体去除甲醛[J]. 环境工程, 2018, 36(12): 125-129. Gao D D, Meng M W, Xie J L, et al. Formaldehyde removal by modified activated carbon fiber combined with plasma[J]. Environmental Engineering, 2018, 36(12): 125-129. |
| [19] | Sekine Y. Oxidative decomposition of formaldehyde by metal oxides at room temperature[J]. Atmospheric Environment, 2002, 36(35): 5543-5547. DOI:10.1016/S1352-2310(02)00670-2 |
| [20] | Ye J W, Zhu X F, Cheng B, et al. Few-layered graphene-like boron nitride:a highly efficient adsorbent for indoor formaldehyde removal[J]. Environmental Science & Technology Letters, 2017, 4(1): 20-25. |
| [21] | Bao M T, Chen Q G, Li Y M, et al. Biodegradation of partially hydrolyzed polyacrylamide by bacteria isolated from production water after polymer flooding in an oil field[J]. Journal of Hazardous Materials, 2010, 184(1-3): 105-110. DOI:10.1016/j.jhazmat.2010.08.011 |
| [22] | Miao L, Wang J L, Zhang P Y, et al. Review on manganese dioxide for catalytic oxidation of airborne formaldehyde[J]. Applied Surface Science, 2019, 466: 441-453. DOI:10.1016/j.apsusc.2018.10.031 |
| [23] | 姜威. 吸附法油烟处理研究[J]. 现代农业, 2009(5): 104-105. |
| [24] | Dhamodharan K, Varma V S, Veluchamy C, et al. Emission of volatile organic compounds from composting:a review on assessment, treatment and perspectives[J]. Science of the Total Environment, 2019, 695. DOI:10.1016/j.scitotenv.2019.133725 |
| [25] | Rezaei M, Moussavi G, Naddafi K, et al. Enhanced biodegradation of styrene vapors in the biotrickling filter inoculated with biosurfactant-generating bacteria under H2O2 stimulation[J]. Science of the Total Environment, 2020, 704. DOI:10.1016/j.scitotenv.2019.135325 |
| [26] |
刘超, 廖雷, 魏建文, 等. 不同填料-液相油烟微生物降解动力学研究[J]. 环境科学与技术, 2016, 39(11): 95-100. Liu C, Liao L, Wei J W, et al. Kinetic study on the biodegrading pollutants from cooking fume in different bio-packing[J]. Environmental Science & Technology, 2016, 39(11): 95-100. |
| [27] |
徐健全, 茹江华, 郑连英. 固定化混合菌降解油烟废气的研究[J]. 中国粮油学报, 2006, 21(4): 97-100, 105. Xu J Q, Ru J H, Zheng L Y. Degradation of oil smoke with immobilized bacteria[J]. Journal of the Chinese Cereals and Oils Association, 2006, 21(4): 97-100, 105. |
| [28] | Yu D S, Song G, Song L L, et al. Formaldehyde degradation by a newly isolated fungus Aspergillus sp. HUA[J]. International Journal of Environmental Science and Technology, 2015, 12(1): 247-254. DOI:10.1007/s13762-013-0411-0 |
| [29] |
谢文娟, 王洁, 孙佩石, 等. 高效降解甲醛菌株的分离鉴定及其特性[J]. 微生物学通报, 2011, 38(11): 1626-1631. Xie W J, Wang J, Sun P S, et al. Isolation and identification of bacterial strains efficient degrading formaldehyde and their degrading characteristics[J]. Microbiology China, 2011, 38(11): 1626-1631. |
| [30] | 东秀珠, 蔡妙英. 常见细菌系统鉴定手册[M]. 北京: 科学出版社, 2001. |
| [31] | Zwietering M H, Jongenburger I, Rombouts F M, et al. Modeling of the bacterial growth curve[J]. Applied and Environmental Microbiology, 1990, 56(6): 1875-1881. DOI:10.1128/AEM.56.6.1875-1881.1990 |
| [32] | Tjørve K M C, Tjørve E. The use of Gompertz models in growth analyses, and new Gompertz-model approach:an addition to the Unified-Richards family[J]. PLoS One, 2017, 12(6). DOI:10.1371/journal.pone.0178691 |
| [33] | Zhou X, Jin W B, Sun C F, et al. Microbial degradation of N, N-dimethylformamide by Paracoccus sp. strain DMF-3 from activated sludge[J]. Chemical Engineering Journal, 2018, 343: 324-330. DOI:10.1016/j.cej.2018.03.023 |
| [34] |
张娟, 杨彩梅, 曹广添, 等. 解淀粉芽孢杆菌及其作为益生菌的应用[J]. 动物营养学报, 2014, 26(4): 863-867. Zhang J, Yang C M, Cao G T, et al. Bacillus amyloliquefaciens and its application as a probiotic[J]. Chinese Journal of Animal Nutrition, 2014, 26(4): 863-867. |
| [35] |
张明, 王洁, 孙珮石, 等. 甲醛降解功能菌的分离鉴定及降解特性[J]. 环境保护科学, 2018, 44(2): 65-70. Zhang M, Wang J, Sun P S, et al. Isolation and identification of bacteria strains able to degrade formaldehyde and their degrading characteristics[J]. Environmental Protection Science, 2018, 44(2): 65-70. |
| [36] |
钟卫鸿, 吴婉欣, 陈雯雯, 等. 一株甲醛降解菌的分离鉴定及降解条件研究[J]. 浙江工业大学学报, 2013, 41(1): 25-30. Zhong W H, Wu W X, Chen W W, et al. Isolation, identification and degrading conditions of a formaldehyde-degrading bacterium[J]. Journal of Zhejiang University of Technology, 2013, 41(1): 25-30. |
| [37] | Marx C J, van Dien S J, Lidstrom M E. Flux analysis uncovers key role of functional redundancy in formaldehyde metabolism[J]. PLoS Biology, 2005, 3(2). DOI:10.1371/journal.pbio.0030016 |
| [38] |
张韦, 宋中邦, 陈丽梅. 甲基营养微生物的甲醛代谢途径及其在环境生物技术中的应用[J]. 生命科学, 2012, 24(3): 266-273. Zhang W, Song Z B, Chen L M. Formaldehyde metabolic pathways in methylotroph and their applications in environmental biotechnology[J]. Chinese Bulletin of Life Sciences, 2012, 24(3): 266-273. |
| [39] | Baghalian K, Hajirezaei M R, Schreiber F. Plant metabolic modeling:achieving new insight into metabolism and metabolic engineering[J]. The Plant Cell, 2014, 26(10): 3847-3866. DOI:10.1105/tpc.114.130328 |
| [40] | Shao Y H, Li J, Wang Y X, et al. Comparative genomics and transcriptomics insights into the C1 metabolic model of a formaldehyde-degrading strain Methylobacterium sp. XJLW[J]. Molecular Omics, 2019, 2(15): 138-149. |
| [41] | Mitsui R, Kusano Y, Yurimoto H, et al. Formaldehyde fixation contributes to detoxification for Growth of a nonmethylotroph, Burkholderia cepacia TM1, on vanillic acid[J]. Applied and Environmental Microbiology, 2003, 69(10): 6128-6132. DOI:10.1128/AEM.69.10.6128-6132.2003 |
| [42] |
程琴, 董蓉, 黄大林, 等. 枯草芽孢杆菌萌发后孢子响应甲醛胁迫过程的拉曼光谱研究[J]. 中国激光, 2011, 38(10): 230-236. Cheng Q, Dong R, Huang D L, et al. Raman spectrum of germinated spores of Bacillus subtilis in response to formaldehyde stress[J]. Chinese Journal of Lasers, 2011, 38(10): 230-236. |