 2020, Vol. 41
2020, Vol. 41

2. 稻米品种安全控制湖南省工程实验室, 长沙 410004
 , CAI Ya-qin1 , ZHOU Hang1,2
, CAI Ya-qin1 , ZHOU Hang1,2 , LIU Ya1 , YANG Wen-jun1 , GU Jiao-feng1,2 , LIAO Bo-han1,2
, LIU Ya1 , YANG Wen-jun1 , GU Jiao-feng1,2 , LIAO Bo-han1,2
2. Hunan Engineering Laboratory for Control of Rice Quality and Safety, Changsha 410004, China
砷(As)是土壤污染的主要元素之一, 2014年全国土壤污染状况调查公报显示, 我国土壤As的点位超标率为2.7%[1].土壤中As污染的成因主要是自然地质和人为活动[2], 开采矿物、煤炭消费、喷施农药化肥和污水灌溉[3, 4]等是造成土壤As污染的主要来源.As是毒性很强的元素之一, 具有致畸性, 可引起肺癌、膀胱癌、肝癌和皮肤癌[5], 增加冠心病发病率[6].无机As及其化合物被国际癌症研究机构(IARC)列为人类Ⅰ类致癌物[7], 慢性接触无机As会损害中枢神经系统[8].三价As可以抑制人体酶的活性, 造成人体代谢紊乱, 引发神经衰弱等问题, 五价As毒性稍弱, 但也可引起脊髓炎和贫血等症状[9].水稻(Oryza sativa L.)是我国主要粮食作物[10], 对As具有较强的累积能力, 能通过食物链威胁人体健康[11].
研究水稻As吸收累积的过程与机制, 是解决水稻As污染问题的关键途径.水稻各部位在不同生育期对As的吸收累积能力有明显差异.段明宇[12]的研究发现水稻在生长前期根系吸收As能力更强, 而程艳等[13]的研究认为水稻在孕穗期对As累积能力更强.水稻根系主要通过磷酸盐转运蛋白和根内的水通道ondulin26蛋白OsNIP2:1(Lsi 1), 分别吸收As(Ⅴ)和As(Ⅲ)[14, 15].水稻吸收的As主要富集在根部, 其中部分As随水稻的生长发育迁移转运至茎、叶、穗、谷壳和糙米中[16], 而水稻糙米中As的累积受水稻不同生育期各部位中As吸收与转运影响.由于水稻各个生育时期生长过程与营养需求状况的差异, 不同生育期对As吸收累积必然存在一定的差异.因此, 本研究通过在不同时期添加外源As来探究水稻不同生育时期吸收累积As的差异, 明确不同生育期As胁迫对水稻成熟期糙米As累积的影响, 确定水稻As累积的关键生育期, 以期为阻控水稻对As的吸收累积及As污染稻田安全利用提供理论支撑.
1 材料与方法 1.1 供试材料本试验采用水稻品种为湘晚籼13号, 为湖南省水稻研究所和金健米业股份有限公司合作选育的常规晚籼品种.水稻培育营养液为木村B营养液[17].
1.2 试验设计本试验地点为长沙市中南林业科技大学水稻盆栽试验基地, 基地光、气、热均为自然状态, 为亚热带季风性气候, 降水丰沛, 气候温和, 雨热同期.用30% H2O2对水稻种子消毒15 min, 用去离子水洗净, 再用湿润的纱布包裹, 置于人工培养箱(温度60℃, 湿度50%)中培养发芽, 芽长为1 cm左右时播种于未受As污染的土壤中. 2018年7月16日, 用自来水冲洗水稻幼苗根系, 去除根系附着的黏土, 用海绵包裹、泡沫浮板固定水稻幼苗, 放入装有18 L营养液的无盖长方体容器(长50 cm, 宽25 cm, 高15 cm)中培养, 保证水稻幼苗生长满足后续试验需要.2018年7月23日, 选择长势均匀一致的水稻幼苗, 五叶一心, 1穴2株, 移入装有3 L营养液的圆柱形塑料桶(高18 cm, 内径17 cm)中进行水培试验.
本试验共设计7个外源As[GSB 04-1714-2004, As标准溶液, As(Ⅲ)]添加处理, 以全生育期不添加外源As为空白对照, 每个处理重复3次, 具体试验设计见表 1.按照试验设计在各生育时期添加外源As, 使营养液中的As浓度为20 μg·L-1, 控制营养液pH为5.5(用NaOH和HCl调节), 每3 d更换一次营养液直至水稻成熟收获.水培试验期间根据实际情况喷洒农药.
|
|
表 1 试验设计与编号 Table 1 Experiment design and numbering |
1.3 样品分析测定方法
2018年11月4日, 培养104 d后收获水稻植株, 用自来水洗净, 然后用纯水、超纯水各2次冲洗, 自然风干后放入烘箱, 于105℃杀青30 min, 70℃烘干至恒重, 分别记录根、茎、叶、穗、谷壳和糙米的干重, 所有样品粉碎并用密封袋保存待测.
水稻样品中的As采用干灰法进行消解(GB/T 5009-2003), 样品溶液中的As含量使用原子荧光光度计(海光, LS-AFS 9530)测定.水稻样品分析过程中以标准物质湖南大米GBW 10045(GSB-23)进行质量控制, As的回收率为97.6%~101.5%, 同时做空白试验.
1.4 数据统计与分析水稻各部位(根、茎、叶、穗、谷壳和糙米)As累积量(μg·株-1)用BAi表示:
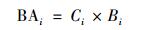
|

|
式中, Ci是水稻各部位(根、茎、叶、穗、谷壳和糙米)As含量(mg·kg-1), Bi是水稻各部位生物量(干重, g·株-1).
水稻不同生育期As胁迫下水稻植株和糙米As累积速率用V表示:
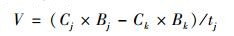
|
水稻不同生育期As累积对水稻成熟期植株、地上部位和糙米As相对贡献率用P表示:
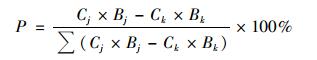
|
式中, Cj是不同生育期As胁迫(分蘖期、拔节期、孕穗期、灌浆期、蜡熟期和成熟期)下各部位As含量(mg·kg-1), Bj是不同生育期As胁迫下各部位生物量(干重, g·株-1); Ck是CK处理各部位As含量(mg·kg-1), Bk是CK处理各部位生物量(干重, g·株-1); 水稻植株和地上部位As累积速率中tj是不同生育期As胁迫(分蘖期、拔节期、孕穗期、灌浆期、蜡熟期和成熟期)的时间, 其中TS、JS、BS、FS、DS和MS处理的tj分别为30、16、13、17、15和13 d; 糙米As累积速率中tj是糙米开始受到As胁迫到成熟收获的时间, 其中TS、JS、BS、FS、DS和MS处理的tj分别为45、45、45、45、28和13 d.
本研究数据统计与分析使用SPSS 23.0软件进行, 采用Duncan多重比较法对各处理数据结果进行差异性分析(P<0.05), 使用Origin 2018绘制图形.
2 结果与分析 2.1 不同生育期As胁迫对水稻生物量的影响水稻植株各部位生物量见表 2, 水稻不同生育期As胁迫对不同部位生物量及总生物量有明显影响.与对照CK相比, 6个单一时期As胁迫处理中仅TS处理使水稻生物量增加, 为121.1 g·株-1, 而其他5个处理均使水稻生物量降低, 降幅为6.9%~24.4%, 其中BS处理最低, 仅为86.1 g·株-1.所有单一时期As胁迫处理糙米和谷壳的生物量均低于对照CK(21.7 g·株-1), 其中BS、DS和MS处理最低, 糙米生物量分别比对照CK降低了15.2%、15.2%和13.8%, 谷壳生物量分别比对照降低了22.7%、21.1%和18.6%.各单一时期As胁迫处理根、茎、叶和穗的生物量均为TS处理最高, 且均显著高于对照CK, 除MS处理茎的生物量以外, 其它处理根、茎、叶和穗的生物量均低于对照CK.各单一时期As胁迫处理中BS处理的根、茎和叶生物量最低, 与对照相比降低了33.8%、35.9%和14.0%.表中还可以看出, 全生育时期As胁迫处理(CG)中水稻各部位生物量和总生物量均低于对照CK, 其中糙米生物量仅为17.6 g·株-1, 显著低于对照CK处理.此外, CG处理糙米、谷壳、穗、叶和茎的生物量均低于各单一时期As胁迫处理.
|
|
表 2 不同生育期As胁迫对水稻生物量的影响1)/g·株-1 Table 2 Effects of As stress at different growth stages on rice biomass/g·plant-1 |
2.2 不同生育期As胁迫对水稻各部位As含量的影响
表 3为不同处理水稻成熟期各部位中As含量.从中可知, 水稻不同生育期As胁迫各部位中As含量均高于对照CK处理.6个单一时期As胁迫处理糙米中As含量均高于对照CK糙米中As含量, 范围为0.08~0.24 mg·kg-1, 大小顺序为BS>FS>JS>MS>TS>DS.各单一As胁迫处理中TS处理根系和茎中As含量最高, 分别为25.75 mg·kg-1和1.17 mg·kg-1, 其次为FS处理, 分别为21.89 mg·kg-1和1.11 mg·kg-1; DS处理根系和茎中As含量最低, 仅为12.22 mg·kg-1和0.43 mg·kg-1, 显著低于其余各单一As胁迫处理.所有单一时期As胁迫处理谷壳和穗中As含量均为BS处理下最高, 分别为0.25 mg·kg-1和2.54 mg·kg-1, 显著高于其他处理, 其次为FS处理, 而DS和MS处理中谷壳和穗中As含量较低.表 3中还可以看出, 全生育时期As胁迫处理(CG)中各部位As含量均显著高于对照CK和各单一As胁迫处理, 其糙米As含量达到0.37 mg·kg-1, 为BS处理的1.5倍.
|
|
表 3 不同生育期As胁迫对水稻各部位As含量的影响/mg·kg-1 Table 3 Effects of As stress at different growth stages on As contents in different rice tissues/mg·kg-1 |
2.3 不同生育期As胁迫对水稻各部位As累积量的影响
图 1为不同处理水稻成熟期各部位中As的累积量.从中可知, 6个单一时期As胁迫处理水稻植株As的累积量均显著高于对照CK, 累积量变化范围为98.9~306.2 μg·株-1, 各部位累积量大小顺序为根系>茎>叶>谷壳>糙米>穗, 其中根系中As的累积量占比最大, 达到65.3%~75.1%, 糙米中As的累积量较低, 仅为0.63%~1.82%.各单一时期As胁迫处理根系中As累积量范围为71.8~228.7 μg·株-1, 其中TS处理最高(228.7 μg·株-1), 其次为FS处理(153.6 μg·株-1), 分别为地上部位As累积量的2.95和1.89倍, 而MS、JS和BS处理根系中As的累积量相当, DS处理根系中As的累积量最低.各处理地上部位As累积变化趋势与根系As累积一致, 各单一时期As胁迫处理地上部位As累积量变化范围为27.2~81.5 μg·株-1, 其中FS处理最高(81.5 μg·株-1), 其次为TS处理(77.5 μg·株-1), 而MS、JS和BS处理地上部位中As的累积量相当, DS处理地上部位中As的累积量最低.全生育时期As胁迫处理(CG)中水稻植株累积量为442.0 μg·株-1, 显著高于对照CK和各单一As胁迫处理, 其中糙米、谷壳、穗、叶、茎和根中As累积量分别为5.7、10.0、2.2、66.7、117.9和239.4 μg·株-1.

|
图 1 不同生育期As胁迫下水稻各部位As累积量 Fig. 1 As accumulation in different rice tissues under As stress at different growth stages |
图 2为不同生育期As胁迫下水稻植株和糙米中As的累积速率.从中可知, 水稻植株As累积速率FS处理最高, 为9.804 μg·d-1, 其次为TS处理(7.926 μg·d-1), 而DS处理最低, 仅为2.032 μg·d-1.各处理糙米中As的累积来源于各生育期As胁迫处理下水稻植株累积的As向糙米中转运.各处理糙米As累积速率范围为0.001~0.069 μg·d-1, 其大小排序为:BS>MS>FS>JS>TS>DS.图 2中在水稻营养生长阶段(分蘖期、拔节期和孕穗期)对水稻进行As胁迫处理, 糙米As的积累速率由TS处理到BS处理呈现逐渐上升趋势, 在BS处理下达到最大累积速率0.069 μg·d-1, 而在水稻生殖生长阶段(灌浆期、蜡熟期和成熟期)对水稻进行As胁迫处理, 糙米As的累积速率MS处理下为0.054 μg·d-1, 而DS处理下仅为0.001 μg·d-1.

|
图 2 不同生育期As胁迫下水稻成熟期各部位As累积速率 Fig. 2 As accumulating rates in different rice tissues at maturity stage under As stress at different growth stages |
表 4为水稻不同生育期As胁迫对各部位As累积的相对贡献率.从中可知, 水稻孕穗期(BS)累积的As对水稻成熟期糙米中As累积量的相对贡献率最大, 为40.3%, 其次为灌浆期(FS)与拔节期(JS), 其相对贡献率分别为26.0%和17.1%, 而成熟期(MS)、分蘖期(TS)和蜡熟期(DS)对糙米中As累积相对贡献率较低, 其相对贡献率分别为9.0%、7.2%和0.4%.对水稻植株和地上部位As累积的相对贡献率而言, 各处理相对贡献率趋势基本一致, 其中分蘖期(TS)和灌浆期(FS)的As累积相对贡献率较高, 而蜡熟期(DS)的相对贡献率最低.
|
|
表 4 水稻不同生育期As胁迫对成熟期各部位As累积的相对贡献率/% Table 4 Relative contributions of As stress at different growth stages on As accumulation in different rice tissues at maturity stage/% |
3 讨论
水稻不同生育期As胁迫下成熟期各部位As含量(表 3)和累积量(图 1)均有着明显的差异.分蘖期As胁迫处理(TS)下, 水稻根系、茎及植株中As累积量最高, 分别达到228.7、44.9和306.2 μg·株-1, 其次是灌浆期As胁迫处理(FS).其原因在于, 一方面分蘖期是水稻发育中的主要营养生长期, 时间跨度最长(30 d), 在这一时期水稻根系快速生长发育, 茎叶大量分蘖并快速生长[18], 根系在吸收营养物质快速生长的同时, 吸收累积了大量的As.另一方面, 水稻分蘖期大量新生长的根系表面还没有覆盖根表铁膜, 而已经形成根表铁膜的部分根段其铁膜厚度还比较薄, 根表铁膜没有起到阻控根系吸收As的屏障作用.有研究表明, 根表铁膜对水稻吸收As具有阻隔或富集作用[19], 在不同生育期根表铁膜发挥作用不同, 在生育初期, 根表铁膜主要起富集作用[20]; 郭伟等[21]在研究分蘖期根表铁膜对水稻As吸收的影响中发现, 根表铁膜促进了水稻根部对As的吸收.随着水稻生长发育, 根表铁膜生长完全, 阻隔了根系对As的吸收, 薛培英等的研究发现生育后期根表铁膜可使水稻根系中As含量比生育初期降低50%~90%[22].
本研究中孕穗期(BS)和灌浆期(FS)As胁迫下糙米中As的累积量最大, 分别为4.5 μg·株-1和3.4 μg·株-1(图 1), 且孕穗期和灌浆期对糙米中As累积贡献较大, 其相对贡献率分别为40.3%和26.0%(表 4).同时孕穗期As胁迫下糙米As累积速率高于其他生育期, 为0.069 μg·d-1, 其次为成熟期和灌浆期(图 2), 说明水稻孕穗期和灌浆期吸收累积的As为糙米中As的主要来源.有研究表明, 孕穗期水稻根系吸收的非结构碳水化合物在茎鞘中累积[23], 然后在灌浆期转运至糙米中, 但孕穗期累积在茎鞘中的As也会在此过程中被夹带而进入糙米中累积, 同时在灌浆期叶片中的As也会通过茎的韧皮部介导转运累积到糙米中[24].还有研究表明, 灌浆期是水稻生殖生长阶段, 根吸收大量养分的同时吸收了大量As, 这些As能通过茎的木质部转运而进入到糙米中[25], 但水稻茎节[26]对As在木质部中的转运有阻隔作用, 水稻茎节是木质部和韧皮部转换的关键器官[27].本研究灌浆期As胁迫下水稻糙米As累积量(3.4 μg·株-1)小于孕穗期As胁迫As累积量(4.5 μg·株-1), 说明水稻糙米中As主要来源于孕穗期韧皮部的转运作用, 其次来源于灌浆期木质部的转运作用.陈丽娜[28]在不同水分管理模式下水稻吸收累积As的研究中发现, 灌浆期到成熟期糙米中的As含量会随着生育期表现出下降趋势, 本研究中灌浆期、蜡熟期和成熟期的相对贡献率分别为26.0%、0.4%和9.0%, 说明水稻糙米As累积能力从灌浆期开始降低.Honma等[29]在抽穗时改变土壤中Eh、酸碱度和溶解态Fe(Ⅱ)的浓度进而有效降低了水稻糙米中的As累积.本研究中水稻糙米中As累积在营养生长阶段(分蘖期和拔节期)、营养生长和生殖并进阶段(孕穗期)、生殖生长阶段(灌浆期、蜡熟期和成熟期)的相对贡献率分别为24.3%、40.33%和38.9%, 进一步说明水稻成熟期糙米中As主要来源于水稻的营养生长和生殖并进阶段, 说明孕穗期是水稻糙米As累积的关键时期, 其次是灌浆期.水稻糙米中所累积的As究竟是主要来源于灌浆期至成熟期水稻籽粒形成过程中根部吸收的, 还是水稻生育前期植株内吸收累积As的转运累积, 还有待进一步研究.
4 结论(1) 不同生育期As胁迫对水稻植株生物量有显著影响, 单一时期As胁迫处理除分蘖期外, 各时期As胁迫下水稻植株生物量均低于对照CK, 所有单一时期As胁迫处理植株生物量均高于全生育时期As胁迫下植株生物量.
(2) 6个单一时期As胁迫处理糙米中As含量范围为0.08~0.24 mg·kg-1, 其中孕穗期As胁迫处理最高, 其次为灌浆期和拔节期As胁迫处理.
(3) 6个单一时期As胁迫处理水稻植株As累积量为98.9~306.2 μg·株-1, 其中分蘖期As胁迫处理最高, 而糙米中As累积量范围为1.4~4.5 μg·株-1, 其中孕穗期As胁迫处理最高.
(4) 孕穗期和灌浆期是水稻成熟期糙米As累积的关键生育期, 糙米As累积速率分别为0.069 μg·d-1和0.045 μg·d-1, 对糙米As累积相对贡献率分别为40.3%和26.0%.
| [1] | 环境保护部.环境保护部和国土资源部发布全国土壤污染状况调查公报[EB/OL].http://www.mee.gov.cn/gkml/sthjbgw/qt/201404/t20140417_270670.htm, 2014-04-17. http://www.cnki.com.cn/Article/CJFDTotal-YQTB201403030.htm |
| [2] | Oremland R S, Stolz J F. The ecology of arsenic[J]. Science, 2003, 300(5621): 939-944. |
| [3] | Li Z J, Yang Q C, Yang Y S, et al. Hydrogeochemical controls on arsenic contamination potential and health threat in an intensive agricultural area, northern China[J]. Environmental Pollution, 2020, 256: 113455. |
| [4] | Liu G, Shi Y, Guo G L, et al. Soil pollution characteristics and systemic environmental risk assessment of a large-scale arsenic slag contaminated site[J]. Journal of Cleaner Production, 2020, 251: 119721. |
| [5] | Meliker J R, Wahl R L, Cameron L L, et al. Arsenic in drinking water and cerebrovascular disease, diabetes mellitus, and kidney disease in Michigan: a standardized mortality ratio analysis[J]. Environmental Health, 2007, 6(1): 4. |
| [6] | Oberoi S, Devleesschauwer B, Gibb H J, et al. Global burden of cancer and coronary heart disease resulting from dietary exposure to arsenic, 2015[J]. Environmental Research, 2019, 171: 185-192. |
| [7] | Tokar E J, Benbrahim-Tallaa L, Ward J M, et al. Cancer in experimental animals exposed to arsenic and arsenic compounds[J]. Critical Reviews in Toxicology, 2010, 40(10): 912-927. |
| [8] | Wang B, Liu J, Liu B, et al. Prenatal exposure to arsenic and neurobehavioral development of newborns in China[J]. Environment International, 2018, 121: 421-427. |
| [9] | Basu A, Mahata J, Gupta S, et al. Genetic toxicology of a paradoxical human carcinogen, arsenic: a review[J]. Mutation Research/Reviews in Mutation Research, 2001, 488(2): 171-194. |
| [10] | 刘笑然, 兰敦臣, 李越. 2014年中国稻米产业研究[J]. 中国粮食经济, 2014(12): 42-47. |
| [11] |
王永杰, 郑祥民, 周立旻. 稻米砷研究进展[J]. 中国水稻科学, 2010, 24(3): 329-334. Wang Y J, Zheng X M, Zhou L M. A review on arsenic in rice[J]. Chinese Journal of Rice Science, 2010, 24(3): 329-334. |
| [12] | 段明宇. AMD污染土壤铁氧化物转化与水稻砷吸收关系研究[D].贵阳: 贵州大学, 2017. http://cdmd.cnki.com.cn/Article/CDMD-10657-1019819025.htm |
| [13] |
程艳, 陈璐, 米艳华, 等. 络合型含铁材料对砷胁迫下水稻生理生化特性及砷累积的影响[J]. 生态学杂志, 2019, 38(8): 2482-2489. Cheng Y, Chen L, Mi Y H, et al. Effects of complexed iron-bearing material on physiological and biochemical characteristics and arsenic accumulation in Oryza sativa L. under arsenic stress[[J]. Chinese Journal of Ecology, 2019, 38(8): 2482-2489. |
| [14] | Zhao F J, Ma J F, Meharg A A, et al. Arsenic uptake and metabolism in plants[J]. New Phytologist, 2009, 181(4): 777-794. |
| [15] | Abedin M J, Feldmann J, Meharg A A. Uptake kinetics of arsenic species in rice plants[J]. Plant Physiology, 2002, 128(3): 1120-1128. |
| [16] |
仲维功, 杨杰, 陈志德, 等. 水稻品种及其器官对土壤重金属元素Pb、Cd、Hg、As积累的差异[J]. 江苏农业学报, 2006, 22(4): 331-338. Zhong W G, Yang J, Chen Z D, et al. Differences in accumulation and distribution of Pb, Cd, Hg and As in rice cultivars and their organs (Oryza sativa L.)[[J]. Jiangsu Journal of Agricultural Sciences, 2006, 22(4): 331-338. |
| [17] | 薛应龙. 植物生理学实验手册[M]. 上海: 上海科学技术出版社, 1985. |
| [18] | 徐春梅.水稻根际氧浓度对分蘖期根系形态和氮代谢的影响机制[D].南昌: 江西农业大学, 2016. |
| [19] |
胡莹, 黄益宗, 刘云霞. 砷污染土壤中不同基因型水稻根表铁膜的形成及其对砷吸收和转运影响[J]. 生态毒理学报, 2013, 8(6): 923-930. Hu Y, Huang Y Z, Liu Y X. Genotypic differences in iron plaque formation on root surface and their effect on arsenic uptake and translocation in rice (Oryza sativa L.) grown in Arsenic contaminated soil[J]. Asian Journal of Ecotoxicology, 2013, 8(6): 923-930. |
| [20] |
胡莹, 黄益宗, 黄艳超, 等. 根表铁锰膜对不同生育期水稻吸收和转运As的影响[J]. 生态毒理学报, 2013, 8(2): 163-171. Hu Y, Huang Y Z, Huang Y C, et al. Influence of iron and manganese plaque on root surface on arsenic uptake and translocation by rice (Oryza sativa L) at different growth stages[J]. Asian Journal of Ecotoxicology, 2013, 8(2): 163-171. |
| [21] |
郭伟, 林咸永, 程旺大. 不同地区土壤中分蘖期水稻根表铁氧化物的形成及其对砷吸收的影响[J]. 环境科学, 2010, 31(2): 496-502. Guo W, Lin X Y, Cheng W D. Effect of iron plaque formation of root surface on As uptake by rice seedlings grown on different types of soils[J]. Environmental Science, 2010, 31(2): 496-502. |
| [22] |
薛培英, 刘文菊, 刘会玲, 等. 中轻度砷污染土壤-水稻体系中砷迁移行为研究[J]. 土壤学报, 2010, 47(5): 872-879. Xue P Y, Liu W J, Liu H L, et al. Arsenic behaviors in the system of arsenic contaminated soil-rhizosphere-rice plants[J]. Acta Pedologica Sinica, 2010, 47(5): 872-879. |
| [23] |
李绍波, 章志宏, 李绍清, 等. 直播水稻茎鞘非结构碳水化合物积累与转运的遗传剖析[J]. 植物学报, 2010, 45(1): 29-35. Li S B, Zhang Z H, Li S Q, et al. Genetic dissection of nonstructural carbohydrate accumulation and translocation in rice culms and leaf sheaths under direct sowing[J]. Chinese Bulletin of Botany, 2010, 45(1): 29-35. |
| [24] | Zhao F J, Stroud J L, Khan M A, et al. Arsenic translocation in rice investigated using radioactive 73As tracer[J]. Plant and Soil, 2012, 350(1): 413-420. |
| [25] | Kobayashi N I, Tanoi K, Hirose A, et al. Characterization of rapid intervascular transport of cadmium in rice stem by radioisotope imaging[J]. Journal of Experimental Botany, 2013, 64(2): 507-517. |
| [26] | Tisarum R, Chen Y S, Dong X L, et al. Uptake of antimonite and antimonate by arsenic hyperaccumulator Pteris vittata: effects of chemical analogs and transporter inhibitor[J]. Environmental Pollution, 2015, 206: 49-55. |
| [27] | Fujimaki S, Suzui N, Ishioka N S, et al. Tracing cadmium from culture to spikelet: noninvasive imaging and quantitative characterization of absorption, transport, and accumulation of cadmium in an intact rice plant[J]. Plant Physiology, 2010, 152(4): 1796-1806. |
| [28] | 陈丽娜.不同水分管理模式下砷在土壤-水稻体系中的时空动态规律研究[D].保定: 河北农业大学, 2009. http://d.wanfangdata.com.cn/thesis/D144805 |
| [29] | Honma T, Ohba H, Kaneko-Kadokura A, et al. Optimal soil Eh, pH, and water management for simultaneously minimizing arsenic and cadmium concentrations in rice grains[J]. Environmental Science & Technology, 2016, 50(8): 4178-4185. |