 2020, Vol. 41
2020, Vol. 41



2. 城市生活污水资源化利用技术国家地方联合工程实验室, 苏州 215009;
3. 江苏省水处理技术与材料协同创新中心, 苏州 215009
2. National and Local Joint Engineering Laboratory of Municipal Sewage Resource Utilization Technology, Suzhou 215009, China;
3. Jiangsu Collaborative Innovation Center of Technology and Material of Water Treatment, Suzhou 215009, China
短程反硝化(partial denitrification, PD)是指异养反硝化菌以有机物为电子供体, 还原NO3--N NO2--N的不完整生物反硝化过程[1].近年来, PD由于可减少60.1%外碳源, 50%氧气需求, 84%废污泥产量等优点[2~5], 被认为是与厌氧氨氧化(anaerobic ammonium oxidation, ANAMMOX)耦合最具前景的NO2--N供给技术, 成为新一轮污水生物脱氮研究的热点课题之一[6].
PD功能的实现很大程度上依赖以NO2--N为产物的反硝化菌属的富集[3, 7], 而不同泥源富集NO2--N积累菌属实现PD工艺启动及其活性表现仍有差异.鉴于工程应用所需大量的PD污泥实际较难获取, 通过驯化常见废污泥为PD污泥, 达到实现污泥资源化和避免对环境二次污染的目的.Cao等[1]的研究发现C/NO3--N=4时缺/好氧污泥短暂的NO2--N积累特征, NO2--N积累量仅为发酵液碳源型反硝化污泥的1/5, 而后者在108 d培养中获得NO3--N NO2--N转化率(nitrate-to-nitrite transformation ratio, NTR)达80%的高效稳定PD效果.毕春雪等[8]在21 d培养中实现了污水厂二沉池污泥和硝化反硝化除磷系统污泥向PD活性污泥的转变, 认为硝化反硝化除磷系统污泥反应器的PD性能最优. Shi等[9]接种城市污水厂废活性污泥启动PD, 运行100 d左右实现了NTR达84%的良好NO2--N积累效果.而这些研究普遍存在启动周期较长缺陷, 且基于常见废泥源实现PD工艺快速启动的研究鲜见报道, 另外, 温度作为影响微生物功能酶代谢活性的关键因素, 尚未见研究人员开展温度变化下对不同泥源驯化PD污泥的影响.
虽然乙酸营养型短程反硝化菌属已被广泛报道[10, 11], 但研究人员并未提供PD污泥驯化成熟初期微生物分布信息[1, 8].此外, 笔者对不同废泥源富集驯化的乙酸型PD污泥微生物种群差异仍缺乏认知, 而理解上述PD污泥菌群分布将对PD实际应用时的稳定高效运行有重要意义.
因此, 本文探究了温度变化下短程反硝化工艺启动及稳定运行过程PD活性和NTR性能, 并基于高通量测序技术分析了不同废污泥源驯化成熟的PD污泥微生物多样性, 旨在为PD耦合ANAMMOX工艺的工程推广作铺垫.
1 材料与方法 1.1 实验装置与运行本实验装置采用3个完全相同的序批式反应器(SBR), 上部敞口安装搅拌器, 反应器材质为有机玻璃, 内直径12 cm, 高30 cm, 有效容积2 L.反应器壁自底部垂直平均布置5个取样口, 通过电磁阀控制排水.反应器在缺氧和室温(15~22℃)条件下运行, 无剩余污泥排放.
本实验共分为2个阶段, 阶段Ⅰ为PD启动阶段(活性上升期), 阶段Ⅱ为负荷提升阶段(活性稳定期), 3个SBR每天周期运行参数一致, 如表 1所示.
|
|
表 1 SBR运行参数表 Table 1 Operational parameters of the SBR |
1.2 进水水质与接种污泥
反应器进水为人工配水, 主要成分是以硝酸钠和无水乙酸钠配制的60 mg ·L-1NO3--N及180~210 mg ·L-1 COD, 配水其他组成如下:0.10 mg ·L-1 MgSO4 ·7H2 O, 0.40 mg ·L-1 CaCl2, 0.05 mg ·L-1 KH2PO3, 1.25 mL ·L-1微量元素A及B溶液, 微量元素配方如文献[12]的描述.此外, 采用1 mol ·L-1 NaOH溶液调节进水pH为9.0±0.1.
本研究S1反应器接种泥源来源于本实验室稳定运行超过180 d的城市污水反硝化除磷系统排泥, 该泥样在-20℃下存储5个月; S2接种自苏州市某污水处理厂污泥浓缩池的混合剩余污泥; S3接种自苏州某高校常年水质轻度富营养化的河涌底泥.本研究所取泥样均经过140目筛网过滤去除体积较大杂质, 再使用去离子水冲洗3遍后分别接种至3个SBR反应器, 接种污泥初始参数如表 2所示.3个PD-SBR反应器运行第14、19和28 d的污泥浓度变化见表 3.
|
|
表 2 初始接种污泥特性 Table 2 Characteristics of the initial inoculation sludge |
|
|
表 3 PD-SBR反应器运行期间污泥浓度变化 Table 3 Sludge concentration changes during the operation of the PD-SBR reactor |
1.3 分析方法
整个研究过程每天取进、出水样监测, 水样经0.45 μm滤纸过滤后, 采用N-(1-萘基)-乙二胺分光光度法测定NO2--N; 麝香草酚分光光度法测定NO3--N; 采用DRB 200型COD快速消解仪测定COD, 本COD数据为校正值, 因为NO2--N对COD测定贡献为1.14 g ·g-1(以COD/NO2--N[13]计); 采用重量法测定混合液悬浮固体浓度(MLSS)和混合液挥发性悬浮固体浓度(MLVSS); 采用30 min沉降法测定SV30及SVI30; 采用雷磁PHSJ-4A型测定pH; 采用哈希便携式溶解氧测定仪(HQ30d)测定温度和溶解氧(dissolved oxygen, DO).
1.4 指标测定方法 1.4.1 SBR长期运行NTR计算方法见式(1):

|
(1) |
式中, NO2--Neff和NO2--Ninf分别为SBR反应器出水和进水NO2--N浓度, mg ·L-1; NO3--Neff和NO3--Ninf分别为SBR反应器出水和进水NO3--N浓度, mg ·L-1.
1.4.2 SBR长期运行出水NO3--N占NOx--N比NP计算方法见式(2):
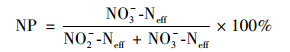
|
(2) |
式中, NO2--Neff和NO3--Neff分别为SBR反应器出水NO2--N和出水NO3--N浓度, mg ·L-1.
1.4.3 SBR典型周期NTR计算方法见式(3):

|
(3) |
式中, NO2--Nt和NO2--Ninitial分别为SBR反应器在t时刻和初始NO2--N浓度, mg ·L-1; NO3--Nt和NO3--Ninitial分别为SBR反应器在t时刻和初始NO3--N浓度, mg ·L-1.
1.4.4 SBR原位批次活性实验比硝态氮还原速率μ(NO3--N)和比亚硝态氮积累速率μ(NO2--N)μ(NO3--N)和μ(NO2--N)是通过拟合曲线计算得到, 即每隔0.083~0.167 h取样测定NO3--N和NO2--N浓度(mg ·L-1)除以污泥MLVSS(g ·L-1)获得, mg ·(h ·g)-1; 后依据μ(NO3--N)和μ(NO2--N)计算批次NTRB(%), 计算方法见式(4)、(5)和(6):

|
(4) |

|
(5) |

|
(6) |
式中, ρ(NO3--N)和ρ(NO2--N)分别为SBR反应器在t时刻NO3--N和NO2--N浓度, mg ·L-1.
1.5 微生物高通量测序分析采集S1、S2和S3的PD-SBR反应器内运行第30 d的污泥样品, 样品对应编号分别为A1、B1和C1.样品由上海美吉生物公司提供测序技术支持, 测序方法见文献[12].
2 结果与分析 2.1 不同废泥源短程反硝化系统快速启动及运行特性图 1 (a)~1(c)分别为32 d内S1、S2和S3反应器短程反硝化启动运行特性.从中可知, 在阶段Ⅰ(1~15 d), 3个反应器均在短时间内实现了PD反应器的启动.S1反应器运行第6 d, 出水NO3--N和NO2--N浓度分别为6.51 mg ·L-1和37.57 mg ·L-1, NTR值稳定维持70%左右, NP值低至14.77%, COD去除率高达100%, 表明S1已具备稳定高效的PD性能.S2启动运行期(1~7 d), 出水NO2--N浓度由19.97mg ·L-1缓慢增加至38.63mg ·L-1, 而剩余NO3--N浓度则减少约20mg ·L-1, 平均出水COD为20.24mg ·L-1, PD活性处于一个逐步上升阶段, 第7 d系统NTR和NP值分别达到69.23%和12.22%, COD去除率接近100%, 表明此时S2具备良好的PD活性和脱硝除碳效果.S3运行第9 d系统NTR和NP分别达到69.98%和13.68%, 此时出水NO2--N和NO3--N浓度分别为37.56 mg ·L-1和5.95 mg ·L-1, 与S1反应器PD活性接近并能够稳定维持, 认为此时S3已基本实现PD反应器的启动.在各个反应器启动期(最长9 d), 均可观察到PD活性的逐步提升, 体现在NO2--N积累量不断增加和剩余NO3--N浓度持续减少, 明显优于文献[8]的报道.这主要是因为反应器运行过程部分反硝化菌属活性受进水条件影响一直被抑制, 系统全程反硝化性能始终处于不良状态, 而以NO2--N为产物的反硝化菌属增殖未受干扰, 因而高效的PD性能得以快速实现.在阶段Ⅱ(16~32 d), 缺氧反应时间由180 min降至60 min, S1、S2和S3反应器NO2--N积累和有机物去除量并未因反应时间的减少发生明显变化, 这意味着PD反应所需时间较为短暂, 而本研究阶段Ⅰ控制的180 min反应时间是为了前期促进反硝化种群的增殖从而缩短反硝化属富集时间, 阶段Ⅱ下调反应器搅拌时间是为了阻断反硝化的连续进行, 进一步提升PD性能.实际上, 本研究中无论180 min还是60 min的反应时间都不会使系统生成的NO2--N被继续还原, 这是因为反硝化菌可利用有机物几乎被全部消耗, 而系统剩余NO3--N的存在又使得NO2--N不会被继续还原为其他氮类物质[14, 15], 这也解释了S1、S2和S3高效的NTR性能、优越的NO2--N积累量和COD去除率.

|
图 1 不同废泥源短程反硝化反应器启动运行特性 Fig. 1 Start-up and operational characteristics of partial denitrification reactors with different waste sludge sources |
特别的是, S1和S2反应器运行期间PD性能受环境条件影响有较为明显的变化, 而S3却能够维持较为稳定的PD活性.伴随室温变化(22.7~14.8℃), S1和S2反应器NO2--N积累量、NP值与NTR性能呈现波动趋势, S3反应器运行则较为平稳.如在20~23 d, 实测3个反应器运行水温由18.2℃降至16℃, S1、S2和S3反应器NO2--N积累浓度分别减少了4.25、5.98和1.96 mg ·L-1, 说明S1与S2反应器响应温度变化较为强烈, S3则对温度变化敏感程度较低.推测这与反应器内接种泥源差异导致的PD污泥不同的温度弹性相关.反应器运行后期(28~32 d), S1和S2开始呈现不同程度的PD活性恶化趋势, S1反应器出水NO3--N浓度达到14.46 mg ·L-1, 平均出水COD升至34.46 mg ·L-1, 出水NO2--N却稳定维持43 mg ·L-1左右, PD性能渐趋下降. S2出水急剧恶化:系统NO2--N浓度下降近40%, 剩余NO3--N浓度由8.98 mg ·L-1增至27.53 mg ·L-1, 甚至超过接种第1 d出水NO3--N浓度25.06 mg ·L-1, S2系统PD性能近乎崩溃.有趣的是, 在第28~32 d, S3反应器仍能够维持平均出水NO2--N和NO3--N浓度约为40.61 mg ·L-1和7.86 mg ·L-1, COD去除率80%以上效果, 表明较S1和S2反应器内PD污泥, S3内接种自河涌底泥的PD污泥抗低温能力较强, 认为此类污泥可能是应用于低温环境下PD启动和运行的优选泥源.当然, 长期低温条件下S3反应器PD活性的变化仍有待进一步观察和研究.
2.2 SBR典型周期不同废泥源短程反硝化系统性能为详细说明不同废泥源PD系统活性和NTR性能, 对各反应器运行第14 d和第28 d典型周期内进行原位批次PD活性实验.
2.2.1 典型周期内氮素转化和COD去除特性图 2所示为S1、S2和S3反应器运行第14 d[图 2 (a)~2(c)]和28 d[图 2 (d)~2(f)]典型周期内氮素转化和COD去除变化曲线, 用以说明PD-SBR运行周期内NO3--N NO2--N转化情况和有机物去除特性.由图可得, 各反应器运行初始阶段, NO2--N迅速积累, 系统NTR值接近整个周期峰值, 有机物和NO3--N短时间内被迅速消耗和转化.Cao等[16]研究原位批次PD活性的实验发现, 搅拌时间为1~2 min PD系统NTR值已达到反应终止时(5~10 min)峰值.与本实验在短时间内获得较优NO2--N积累量结果相似.由图 2(a)~2(c)可知, 第14 d时典型周期内S1、S2和S3平均NTR分别为79.66%、81.52%和64.20%, 最高积累NO2--N浓度分别为42.19、41.79和38.73 mg ·L-1, 反应终止时COD浓度分别为46.24、60.28和47.38 mg ·L-1, 可以看出S1和S2反应器PD性能优于S3.反应器运行至第28 d时, S1、S2和S3平均NTR分别增加了5.11%、10.63%和31.95%, 最高NO2--N浓度略微增加, 表明随着反应器运行, PD活性和系统NTR性能也不断提升, S3反应器PD性能已达到较优运行状态.对图 2进行比较可知, 第28 d时S1和S2反应器运行终点COD浓度明显高于第14 d, 分别增加了42.67 mg ·L-1和91.05 mg ·L-1, 而S3终点COD浓度仅增加约13 mg ·L-1, COD浓度的增加与微生物裂解释放胞内有机物相关, 与2.1节叙述的3个PD-SBR反应器的性能变化相对应.

|
图 2 PD-SBR反应器典型周期内氮素转化和COD去除变化曲线 Fig. 2 Variation of nitrogen conversion and COD removal in a typical cycle for PD-SBR reactors |
PD-SBR反应器运行过程NO2--N达到峰值后基本保持稳定, 随后反应器剩余COD各自维持在一定范围以内, 且系统还会残留少量NO3--N.实际上, 此时剩余有机物很可能是微生物代谢释放的可溶性微生物产物(SMP), Cao等[17]的研究证明了这一点:在原位PD活性批次实验中, 以乙酸钠作为唯一进水碳源, 通过气相色谱未检测到出水乙酸盐的存在.SMP易在内源呼吸和饥饿状态下产生, 而反应器内培养的乙酸营养型PD微生物难以利用SMP物质[9], 故外碳源的匮乏使得系统NO3--N还原和NO2--N生成速率严重减缓.整个周期内, NO2--N积累量随NO3--N还原而持续增加, 但当外碳源不足时, 氮素转化速率严重放缓.
2.2.2 典型周期批次短程反硝化活性对PD-SBR反应器运行第2、14和28 d活性进行原位测定, 以比反硝化速率计.如图 3所示, 运行后期S1和S2较接种初期的比硝态氮还原速率和比亚硝态氮积累速率有显著提高, 在第28 d时的比反硝化速率甚至达到第2 d的6倍之多, S3则呈先上升后下降趋势, 说明PD-SBR反应器运行期间, S1和S2的PD功能菌属活性得到了极大提升, S3比硝态氮还原速率和比亚硝态氮积累速率则维持在约64.09 mg ·(h ·g)-1和35.42 mg ·(h ·g)-1, 这可能是由于S3较大的污泥浓度(3.08~3.60 g ·L-1, 以VSS计, 见表 3)使得比反硝化速率很难有明显增加而维持在一定范围内.第28 d时, S2比硝态氮还原速率达到了301.48 mg ·(h ·g)-1, 比亚硝态氮积累速率为276.40 mg ·(h ·g)-1, 远远超过Cao等[18]报道的PD原位最大比硝态氮还原速率84.9 mg ·(h ·g)-1的数值, 这是由于此时受持续低温影响污泥逐渐解体引起MLVSS值仅为第14 d时的1/4(见表 3), 导致了PD比反硝化活性的显著提升, 但NO3--N还原量却因生物量的降低而减少.

|
图 3 PD-SBR反应器典型周期原位批次比反硝化活性和NTR性能 Fig. 3 Typical cycle in-situ batch denitrification activity and NTR performance of PD-SBR reactors |
典型周期内S1的NTRB维持在79.81% ~91.25%, S2平均NTRB达到了88.29%, S3的NTRB则整体维持60%左右.表明PD-SBR反应器运行期间能够维持稳定的PD活性和NTR效率, S1和S2批次PD活性与Cao等[18]获得的平均88.3%的NTR效率接近, 但高于之前报道的PD长期运行80%的NTR效果[1].NTRB是基于比亚硝态氮积累速率和比硝态氮还原速率的比值获得, 而S3长期运行过程中(见图 1)NO2--N积累量明显低于S1和S2, 说明S3反应器内以NO2--N为最终产物的反硝化种群活性在3个SBR中最低, 故导致了批次实验中相对较小的比NO2--N积累速率和NTR性能.
2.3 不同废泥源短程反硝化系统微生物群落组成 2.3.1 门水平微生物群落结构图 4为不同泥源PD-SBR反应器运行30 d污泥样品门水平微生物丰度群落(微生物相对丰度>1.0%为主要菌门).从中可知, 不同泥源的污泥样品共检测出7个主要菌门, 分别为变形菌门(Proteobacteria)、拟杆菌门(Bacteroidetes)、绿弯菌门(Chloroflexi)、厚壁菌门(Firmicutes)、Patescibacteria菌门、酸杆菌门(Acidobacteria)和放线菌门(Actinobacteria).按照门水平丰度高低排序, S1反应器主要菌门分别为变形菌门(56.17%)、拟杆菌门(21.08%)、厚壁菌门(12.43%)和绿弯菌门(7.82%); S2依次为拟杆菌门(48.77%)、变形菌门(38.40%)和绿弯菌门(6.08%); S3反应器内变形菌门(78.50%)和拟杆菌门(13.45%)则构成了超过90%的门水平物种.这说明PD-SBR反应器功能菌群落多样性低, 富集程度高[12], 特别是S3反应器.

|
图 4 不同泥源PD-SBR反应器门水平微生物群落柱状图 Fig. 4 Bar chart showing the microbial community at the phylum level for PD-SBR reactors with different sludge sources |
图 5为不同泥源PD-SBR反应器运行30 d污泥样品属水平微生物丰度群落图(属水平相对丰度>1.0%为主要分析对象).明显地, Acinetobacter属在S1中丰度高达40.60%, 是该反应器绝对优势菌属, 其次分别是Flavobacterium(8.46%)、Exiguobacterium(5.42%)和Thauera(4.71%)等, 以上4个属分别对应变形菌门、拟杆菌门、厚壁菌门和变形菌门.S2中Flavobacterium(27.79%)相对丰度占比最高, 后面依次为norank_f_Saprospiraceae(11.08%)、Ferruginibacter(4.25%)、Defluviimonas(3.99%)、norank_f_Rhodanobacteraceae(3.81%)和Thauera(3.60%)等.S3反应器内部优势菌属为Thauera(25.09%)、Dechloromonas(18.98%)、norank_f_Steroidobacteraceae(7.94%)及norank_f_Rhodanobacteraceae(4.68%)等.显然, Thauera属主导了S3反应器菌属群落.

|
图 5 不同泥源PD-SBR反应器属水平微生物群落柱状图 Fig. 5 Bar chart showing the microbial community at the genus level for PD-SBR reactors with different sludge sources |
3个反应器在较短时间内均实现了PD的快速启动.原因是本研究始终控制反应条件:进水pH为8.90~9.10、COD/NO3--N为2.75~3.76. pH被证明是选择富集NO2--N积累菌属的有效条件[9, 13, 19], 反硝化过程中NO3--N还原为N2是连续的4步酶促反应, 反应的连续进行与电子接收、传递和消耗有关[3], 高pH则会降低反硝化过程碳氧化速率, 进而加剧反硝化酶之间的电子竞争[20], NO2--N还原酶活性明显受到高碱度抑制, 而NO3--N还原酶活性基本不受影响, 这导致了NO2--N暂时的积累[13, 21].Qian等[13]的研究表明, 控制进水pH=9.0更利于富集NO3--N还原酶基因数高的反硝化物种; Shi等[9]在进水pH=9.0条件下, 提升废污泥NTR值达84%的良好PD活性.因此, 本研究控制的进水pH为9.0±0.1条件是实现3个反应器快速启动PD和有效NO2--N积累的关键因素, 能够快速筛选NO2--N积累菌群和抑制NO2--N还原菌属增殖.应当指出的是, 虽然本研究3个PD-SBR反应器获得平均出水NTR最高为75.92%, 低于研究人员认为平均NTR=80%是PD反应启动成功的标志[1], 但80%的NTR效率是基于PD反应过程中取样点时刻NO3--N还原量和NO2--N积累量而得, 而本研究S1、S2和S3在运行期间能够保持70%左右出水NTR效率, 因而认为本实验中反应器出水NTR=70%是PD启动的一个标志.另外, 限制低COD/NO3--N比可以促进PD功能菌的增殖, 因为反硝化物种会优先储存易被利用碳源而非直接利用[21, 22].COD/NO3--N比很大程度上决定了反硝化进程, 控制硝酸盐还原速率[23].相关研究表明COD/NO3--N为2.0~3.5是PD高效运行较优的控制条件[3, 4, 24], 本研究实际运行中同样发现在COD/NO3--N为3.0~3.5范围内PD活性较优, 两者还可能存在正相关性.启动期间反硝化物种通过种间选择, 在乙酸盐有机物、高碱度和适宜COD/NO3--N比的反应条件下, NO2--N积累菌迅速占据优势地位[24], NO2--N还原菌的增殖则被严重抑制, 因而3个PD-SBR反应器在短期内快速启动同时保持运行的高效性.
值得注意的是, 本研究S1、S2和S3在32 d的运行中分别获得平均NTR为75.92%、73.36%和69.90%效果, 均明显优于Qian等[13]接种反硝化污泥在进水pH=9.0条件下PD反应器平均出水NTR=57%, 这可能是因为后者控制COD/NO3--N比为3.0, 电子供体不足引起了NO3--N还原速率下降.
3.2 不同废泥源短程反硝化系统性能变化分析碳源的选取对于PD反应速率有很大影响, Le等[23]比较了多种碳源对短程反硝化短期影响, 发现乙酸和甘油显示出高效的PD选择, 并推测与电子传递途径相关. Li等[25]同样发现相比乙酸物质, 葡萄糖反硝化反应会上调乙酸产出量和下调乙酸利用率导致细胞内NADH/NAD+比下降, 进而降低脱硝速率, 因而认为乙酸盐是NO2--N积累的优选碳源.广泛的研究同样表明, 乙酸作为简单小分子物质极易被微生物利用, 因而PD反应选取乙酸作为电子供体能够促进乙酸营养型PD种群快速增殖, 加快PD反应进程[3, 18, 26]. 3个PD-SBR反应器均以乙酸钠作为唯一碳源, 导致了剧烈的PD反应速率.批次实验表明, 反应器在阶段Ⅰ运行第14 d, 60 min时系统NO2--N积累量近乎峰值, 随着运行时间推移, 阶段Ⅱ运行第28 d时S1、S2和S3反应器30 min时刻NO2--N浓度分别达44.31、42.43和41.09 mg ·L-1, 高于或接近反应终点浓度, 表明本研究可进一步减少缺氧搅拌时间以获得更高的NO2--N产量.
近期, Meng等[27]的研究发现硝化菌和反硝化菌能够承受较高范围的温度变化扰动, 但持续低温下传统反硝化菌相对丰度仍会逐渐下降[28].S1和S2反应器种泥由于长期处于中温(~25℃)环境, 耐冷菌丰度不足, 因而温度降低反硝化活性也会随之发生明显减弱, 长期低温抑制下S2污泥甚至失活死亡(观察到明显的污泥白化, 出水透明度较低).S3取自河道底泥的表层反硝化菌可能适应了自然界中比生物反应器更低的温度[29], 因此低温影响下PD活性并未受明显影响.
3.3 不同废泥源短程反硝化系统微生物种群分析接种泥源为城市污水处理系统反硝化除磷单元的S1反应器, 变形菌门、拟杆菌门、厚壁菌门和绿弯菌门为主导菌门, 而在城市污水培养的PD污泥也检测出类似菌门[9].变形菌门在良好性能的活性污泥系统含量最高, 包含了氮循环功能相关的氨氧化菌、亚硝酸盐氧化菌和反硝化菌等[30], 先前的PD系统中也发现变形菌门通常占比极高(44.1% ~82.3%)[31], 与本研究类似.因此高丰度的变形菌门是PD系统高效NTR的保证, PD-SBR变形菌门丰度高低依次为S3>S1>S2(78.50%>56.17%>38.40%), 对应反应器运行至30 d时PD性能S3最优, S1渐趋恶化, S2近乎崩溃.拟杆菌门和绿弯菌门通常具备降解易溶有机物(如短链脂肪酸)、胞外聚合物和衰亡菌体等功能[32], 拟杆菌门还与反硝化作用和颗粒形成紧密相关[12], S2中两类菌门占比高达54.85%, 远高于S1和S3[28.9%和13.45%(仅拟杆菌门)], 进一步说明S2污泥严重衰亡引起了拟杆菌门和绿弯菌门的大量增殖.
Thauera属为广泛报道的PD反应器中实现NO2--N积累的功能菌属[9, 12, 13], 其在微生物群落中丰度分布对PD活性影响极大.反应器运行至30 d时, S3中Thauera属相对丰度高达25.09%, 而S1和S2中仅有4.71%和3.60%, 这解释了S3反应器在长期低温影响下仍能保持良好PD活性和NO2--N积累效果的特性.有研究表明, 随温度下降, 提升功能菌属生物量丰度能够维持微生物氮降解活性[33].长期运行S1和S2反应器平均NTR分别为75.92%和73.36%, 均高于S3的69.90%, 这可能是因为在适宜温度下, S1和S2内Thauera属活性高于S3, 而长期低温S1和S2内Thauera属活性和丰度下降, S3内Thauera属则维持高丰度.S1中属于变形菌门的Acinetobacter属是其优势菌属, 在污水处理系统发挥高效的自养反硝化作用, 因而有利于维持S1系统的反硝化性能[34]. S2菌属中Flavobacterium、norank_f_Saprospiraceae和Ferruginibacter比例达43.12%, 均属于可利用死亡细菌残体物质的拟杆菌门, 解释了该系统污泥衰亡恶化、性能崩溃的现象.
4 结论(1) 在22~23℃, pH=9.0±0.1和COD/NO3--N为3.0~3.5条件下, S1、S2和S3分别在第6、7和9 d成功启动PD, 系统长期运行平均NTR为S1>S2>S3(75.92%>73.36%>69.90%).
(2) 短程反硝化活性批次实验表明, 乙酸钠为碳源和适宜COD/NO3--N比的进水水质, PD-SBR反应器获得良好的比反硝化速率和NTR性能, 第28 d典型周期内S1、S2和S3平均NTR分别为84.77%、92.15%和96.15%, 最大比NO2--N积累速率为116.95、276.40和36.73 mg ·(h ·g)-1.
(3) 运行期间, S1和S2反应器PD活性响应温度变化较为明显, 而S3反应器PD活性对温度敏感程度较低, 推测河涌底泥可能是低温实现短程反硝化快速启动和稳定运行的优选种泥.
(4) 变形菌门和拟杆菌门是PD反应器主导微生物菌门, PD活性和高NO2--N积累量取决于属于变形菌门的Thauera属丰度.S3反应器Thauera属丰度高达25.09%, 是其维持低温PD活性的重要原因.
| [1] | Cao S B, Wang S Y, Peng Y Z, et al. Achieving partial denitrification with sludge fermentation liquid as carbon source:the effect of seeding sludge[J]. Bioresource Technology, 2013, 149: 570-574. |
| [2] | Cao S B, Du R, Peng Y Z, et al. Novel two stage partial denitrification (PD)-Anammox process for tertiary nitrogen removal from low carbon/nitrogen (C/N) municipal sewage[J]. Chemical Engineering Journal, 2019, 362: 107-115. |
| [3] | Du R, Peng Y Z, Ji J T, et al. Partial denitrification providing nitrite:Opportunities of extending application for anammox[J]. Environment International, 2019, 131. DOI:10.1016/j.envint.2019.105001 |
| [4] | Zhang M, Wang S Y, Ji B, et al. Towards mainstream deammonification of municipal wastewater:partial nitrification-anammox versus partial denitrification-anammox[J]. Science of the Total Environment, 2019, 692: 393-401. |
| [5] | Cao S B, Oehmen A, Zhou Y. Denitrifiers in mainstream anammox processes:Competitors or supporters?[J]. Environmental Science & Technology, 2019, 53(19): 11063-11065. |
| [6] | Ma B, Wang S Y, Cao S B, et al. Biological nitrogen removal from sewage via anammox:recent advances[J]. Bioresource Technology, 2016, 200: 981-990. |
| [7] | Ma B, Xu X X, Wei Y, et al. Recent advances in controlling denitritation for achieving denitratation/anammox in mainstream wastewater treatment plants[J]. Bioresource Technology, 2019, 299: 299. DOI:10.1016/j.biortech.2019.122697 |
| [8] |
毕春雪, 于德爽, 杜世明, 等. 乙酸钠作为碳源不同污泥源短程反硝化过程亚硝酸盐积累特性[J]. 环境科学, 2019, 40(2): 783-790. Bi C X, Yu D S, Du S M, et al. Nitrite accumulation characteristics of partial denitrification in different sludge sources using sodium acetate as carbon source[J]. Environmental Science, 2019, 40(2): 783-790. |
| [9] | Shi L L, Du R, Peng Y Z. Achieving partial denitrification using carbon sources in domestic wastewater with waste-activated sludge as inoculum[J]. Bioresource Technology, 2019, 283: 18-27. |
| [10] | Du R, Cao S B, Li B K, et al. Step-feeding organic carbon enhances high-strength nitrate and ammonia removal via DEAMOX process[J]. Chemical Engineering Journal, 2019, 360: 501-510. |
| [11] | Du R, Cao S B, Li B K, et al. Performance and microbial community analysis of a novel DEAMOX based on partial-denitrification and anammox treating ammonia and nitrate wastewaters[J]. Water Research, 2017, 108: 46-56. |
| [12] | Du R, Cao S B, Zhang H Y, et al. Formation of partial-denitrification (PD) granular sludge from low-strength nitrate wastewater:the influence of loading rates[J]. Journal of Hazardous Materials, 2020, 384. DOI:10.1016/j.jhazmat.2019.121273 |
| [13] | Qian W T, Ma B, Li X Y, et al. Long-term effect of pH on denitrification:High pH benefits achieving partial-denitrification[J]. Bioresource Technology, 2019, 278: 444-449. |
| [14] | Le T, Peng B, Su C Y, et al. Nitrate residual as a key parameter to efficiently control partial denitrification coupling with anammox[J]. Water Environment Research, 2019, 91(11): 1455-1465. |
| [15] | Du R, Peng Y Z, Cao S B, et al. Mechanisms and microbial structure of partial denitrification with high nitrite accumulation[J]. Applied Microbiology and Biotechnology, 2016, 100(4): 2011-2021. |
| [16] | Cao S B, Du R, Niu M, et al. Integrated anaerobic ammonium oxidization with partial denitrification process for advanced nitrogen removal from high-strength wastewater[J]. Bioresource Technology, 2016, 221: 37-46. |
| [17] | Cao S B, Peng Y Z, Du R, et al. Characterization of partial-denitrification (PD) granular sludge producing nitrite:effect of loading rates and particle size[J]. Science of the Total Environment, 2019, 671: 510-518. |
| [18] | Cao S B, Du R, Zhang H Y, et al. Understanding the granulation of partial denitrification sludge for nitrite production[J]. Chemosphere, 2019, 236: 124389. DOI:10.1016/j.chemosphere.2019.124389 |
| [19] | Si Z, Peng Y Z, Yang A M, et al. Rapid nitrite production via partial denitrification:pilot-scale operation and microbial community analysis[J]. Environmental Science:Water Research & Technology, 2018, 4(1): 80-86. |
| [20] | Pan Y T, Ye L, Ni B J, et al. Effect of pH on N2O reduction and accumulation during denitrification by methanol utilizing denitrifiers[J]. Water Research, 2012, 46(15): 4832-4840. |
| [21] | Li W, Lin X Y, Chen J J, et al. Enrichment of denitratating bacteria from a methylotrophic denitrifying culture[J]. Applied Microbiology and Biotechnology, 2016, 100(23): 10203-10213. |
| [22] | Ji J T, Peng Y Z, Li X Y, et al. Stable long-term operation and high nitrite accumulation of an endogenous partial-denitrification (EPD) granular sludge system under mainstream conditions at low temperature[J]. Bioresource Technology, 2019, 289. DOI:10.1016/j.biortech.2019.121634 |
| [23] | Le T, Peng B, Su C Y, et al. Impact of carbon source and COD/N on the concurrent operation of partial denitrification and anammox[J]. Water Environment Research, 2019, 91(3): 185-197. |
| [24] | Zhang Z Z, Zhang Y, Chen Y G. Recent advances in partial denitrification in biological nitrogen removal:from enrichment to application[J]. Bioresource Technology, 2020, 298: 298. DOI:10.1016/j.biortech.2019.122444 |
| [25] | Li W, Liu S, Zhang M, et al. Oxidation of organic electron donor by denitratation:Performance, pathway and key microorganism[J]. Chemical Engineering Journal, 2018, 343: 554-560. |
| [26] | Du R, Cao S B, Li B K, et al. Synergy of partial-denitrification and anammox in continuously fed upflow sludge blanket reactor for simultaneous nitrate and ammonia removal at room temperature[J]. Bioresource Technology, 2019, 274: 386-394. |
| [27] | Meng Y B, Zhou Z B, Meng F G. Impacts of diel temperature variations on nitrogen removal and metacommunity of anammox biofilm reactors[J]. Water Research, 2019, 160: 1-9. |
| [28] | Xu J, Pang H L, He J G, et al. Start-up of aerobic granular biofilm at low temperature:performance and microbial community dynamics[J]. Science of the Total Environment, 2020, 698: 698. DOI:10.1016/j.scitotenv.2019.134311 |
| [29] | Zhou S, Borjigin S, Riya S, et al. The relationship between anammox and denitrification in the sediment of an inland river[J]. Science of the Total Environment, 2014, 490: 1029-1036. |
| [30] |
彭永臻, 钱雯婷, 王琦, 等. 基于宏基因组的城市污水处理厂生物脱氮污泥菌群结构分析[J]. 北京工业大学学报, 2019, 45(1): 95-102. Peng Y Z, Qian W T, Wang Q, et al. Unraveling microbial structure of activated sludge in a full-scale nitrogen removal plant using metagenomic sequencing[J]. Journal of Beijing University of Technology, 2019, 45(1): 95-102. |
| [31] | Cao S B, Du R, Li B K, et al. Nitrite production from partial-denitrification process fed with low carbon/nitrogen (C/N) domestic wastewater:performance, kinetics and microbial community[J]. Chemical Engineering Journal, 2017, 326: 1186-1196. |
| [32] |
王晗, 李瀚翔, 陈猷鹏, 等. 盐度条件下ANAMMOX-EGSB反应器颗粒污泥微生物群落[J]. 环境科学, 2019, 40(4): 1906-1913. Wang H, Li H X, Chen Y P, et al. Microbial community of granular sludge in an ANAMMOX-EGSB reactor under saline conditions[J]. Environmental Science, 2019, 40(4): 1906-1913. |
| [33] | Straka L, Summers A, Stahl D A, et al. Kinetic implication of moving warm side-stream Anaerobic ammonium oxidizing bacteria to cold mainstream wastewater[J]. Bioresource Technology, 2019, 288: 288. DOI:10.1016/j.biortech.2019.121534 |
| [34] | Xu L Z J, Xia W J, Yu M J, et al. Merely inoculating anammox sludge to achieve the start-up of anammox and autotrophic desulfurization-denitrification process[J]. Science of the Total Environment, 2019, 682: 374-381. |