 2020, Vol. 41
2020, Vol. 41

2. 上海市环境科学研究院, 上海 200233
 , CHEN Xiao-hua2 , SHEN Gen-xiang2
, CHEN Xiao-hua2 , SHEN Gen-xiang2 , ZHU Ying2 , QIAN Xiao-yong2 , ZHANG Xin-liang2 , ZHANG Wei1 , HU Shuang-qing2 , BAI Yu-jie2
, ZHU Ying2 , QIAN Xiao-yong2 , ZHANG Xin-liang2 , ZHANG Wei1 , HU Shuang-qing2 , BAI Yu-jie2
2. Shanghai Academy of Environmental Sciences, Shanghai 200233, China
浮游植物在水生生态系统物质循环和能量流动的过程中起到基础性作用, 是水生生态系统中的初级生产者[1].浮游植物群落与河流、湖泊的水环境因子之间会相互影响.由于浮游植物体积小, 结构简单, 对生境变化较为敏感, 群落结构的变化可以在一定程度上反映环境因素的变化[2, 3], 具有重要的指示意义[4].浮游植物的暴发会给河湖生态系统带来巨大压力, 使整个生态系统的稳定性和多样性遭到破坏.尤其是蓝藻的暴发, 不仅会影响水体的景观, 还会使水体变臭.浮游植物(如微囊藻等有毒藻类)大量聚集于水源地取水口附近, 可能会污染水源, 从而危及供水安全[5]. 2007年太湖的蓝藻暴发引发无锡全城自来水污染更是为人们敲响了警钟.因此, 藻华问题一直持续广受关注[6~11].位于郊区的黄浦江金泽水库是上海城市供水水源地之一, 为典型的河流型水库, 其水源水质受当地河道水体影响较大, 藻类监测可进一步有效保障该区域的饮用水水质安全.
上海水体浮游植物研究主要集中在某一具体湖泊或者是水库, 如淀山湖、滴水湖和青草沙水库等区域水体[12~14], 而对于某一区域内所有河流的浮游植物这种相对宏观的问题缺少探讨.本文面向上海市全市, 选取了包括中心城区、新城镇和农村地区总计44个样点开展调查.中心城区河道主要是指周边区域为建成区的河道; 新城镇河道主要是指周边区域为规划建设区的河道, 河道周边可能既有居民区, 也有农田; 农村河道指的是周边区域为农业生产用地或农村建设用地.在测定水质理化数据的基础上, 研究浮游植物与环境因子的关系及不同区域浮游植物的时空分布特征, 从生物角度对上海市不同区域的富营养化问题进行解读, 并探讨造成浮游植物区域分布差异的影响因素, 以期为后续上海中小河道富营养化问题的研究提供新的角度和思路.
1 材料与方法 1.1 研究区域与样点设置上海市地处长江三角洲平原地带, 辖内河湖众多, 水网密布, 是典型的平原河网地区.上海河网水体流动性复杂, 采取分片控制的治水方针, 利用泵闸等水利片控制工程进行排涝、引水和水资源调度等[15].本研究在上海市选取了14个区41条河道, 总计布设了44个样点, 其中A1~A15位于中心城区河道, B1~B16位于新城镇河道, C1~C13位于农村河道, 具体样点空间分布如图 1所示.采样分两批进行, 一批集中于2018年9~10月(秋季), 一批集中于2019年7~8月(夏季).

|
图 1 采样点位分布示意 Fig. 1 Distribution of sampling sites |
定性样品的采集及分析:使用25号浮游生物网, 在水面呈“∞”状巡回拖动浮游生物网, 然后打开浮游生物网的旋塞, 让网内浓缩的水样流入圆形广口瓶内, 重复操作数次, 取样50 mL, 用1% ~1.5%样品体积的鲁哥氏液摇匀固定.定性样品均使用光学显微镜鉴定拍照, 浮游植物鉴定方法参考文献[16].
定量样品的采集及分析:使用采水器在表层0.5 m处采集1 L水样, 加入10 mL鲁哥氏液摇匀固定, 回到实验室后静置沉淀48 h, 用虹吸管小心缓慢地抽掉上清液, 最后浓缩到50 mL, 加入1 mL福尔马林液密封保存.定量样品计数前先摇匀, 然后镜检计数.
1.3 水环境理化因子的测定与分析现场采用便携式溶解氧仪(YSI PRO 2010)测定河水pH值、水温(WT).采用赛氏盘现场测定水体透明度(SD).水样的浊度(Turb)依据GB 13200-91的分光光度法测定, 总氮(TN)依据GB 11894-8989的碱性过硫酸钾消解紫外分光光度法测定, 总磷(TP)依据GB 11893-89的钼酸铵分光光度法测定, 氨氮(NH4+-N)依据GB 7479-87的纳氏试剂比色法测定, 化学需氧量(COD)依据GB 11914-89的重铬酸钾法测定.
1.4 数据处理本文通过Origin 2015和Excel 2013软件对数据进行统计、作图和分析; 使用ArcGIS软件对上海市典型河道采样点位分布图进行绘制.对典型河道浮游植物物种进行筛选(该物种在至少一个采样点位处的相对密度≥1%, 且该物种在各点位出现的频度≥30%)[17], 将筛选物种数据都进行对数lg(x+1)处理后[18], 使用Canoco 4.5对筛选物种数据和环境因子数据进行冗余分析(redundancy analysis, RDA).
物种优势度的计算公式[19]:
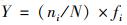
|
(1) |
式中, ni为采样点位中i种的个数; N为采样点位中浮游植物总个数; fi为i种在各采样点位中出现的频率.
2 结果与分析 2.1 浮游植物群落的季节变化秋季、夏季全市浮游植物分别有8门149种和8门185种, 夏季浮游植物种数高出秋季24%. 3个地区的种类进行单因素方差分析.结果显示:两个季度的种类数差异不显著(P>0.05), 即季节的变化引起的各门类种数的变化并不明显.绿藻门的种数在两个季度中始终占最高比例[图 2(a)], 分别为34%和35%, 其次为蓝藻和硅藻.裸藻门种数所占比例夏季比秋季略微下降, 硅藻门略微上升, 但是变化幅度不大.

|
图 2 秋季、夏季浮游植物种类比例和密度分布 Fig. 2 Phytoplankton species proportion and density distribution during the autumn and summer |
除新城镇外, 中心城区和农村地区的浮游植物密度季节差异显著(P < 0.05).秋季全市浮游植物平均密度为361.07×104 cells·L-1, 夏季则为1 359.44×104 cells·L-1, 夏季比秋季高出2.77倍.蓝藻门密度在秋季和夏季占比始终最大[图 2(b)], 夏季全市蓝藻门平均密度为883.26×104 cells·L-1, 秋季全市蓝藻门平均密度为268.59×104 cells·L-1, 高出秋季2.29倍.
秋季全市微囊藻优势度最高(表 1), 为0.16, 高出其它优势种1倍以上.夏季优势种优势度差异不大, 变异直链藻优势度最大, 为0.05.
|
|
表 1 中心城区、新城镇和农村地区秋季、夏季浮游植物优势种及优势度 Table 1 Dominant species and degree of phytoplankton during the autumn and summer in the central urban area, new town, and rural area |
2.2 浮游植物群落的空间变化
中心城区浮游植物秋季共有6门77种, 夏季共有8门116种, 高出秋季50.6%;新城镇地区浮游植物秋季共有8门93种, 夏季共有8门117种, 高出秋季25.8%;农村地区浮游植物秋季共有7门92种, 夏季共有8门108种, 高出秋季17.4%(图 3).

|
图 3 中心城区、新城镇和农村地区秋季、夏季浮游植物种类比例 Fig. 3 Ratio of phytoplankton species during the autumn and summer in the central urban area, new town, and rural area |
方差齐性检验结果显示:中心城区、新城镇和农村这3个地区浮游植物密度的方差在P=0.05前提下是相等的(其中秋季Sig.=0.571>0.05, 夏季Sig.=0.121>0.05), 可进行方差分析.所有浮游植物平均密度和蓝藻门平均密度均呈现农村>新城镇>中心城区(图 4), 蓝藻门密度在各地区均为最高, 占比61% ~80%.秋季各地区浮游植物平均密度的差异均不显著(P>0.05), 而夏季农村地区浮游植物平均密度显著高于中心城区(P < 0.05).夏季农村地区浮游植物平均密度是各地区中最高的, 为2 045.12×104cells·L-1, 高出浮游植物密度最低的中心城区1.82倍.夏季农村地区蓝藻门平均密度是各地区中最高的, 为1 302.66×104 cells·L-1, 高出蓝藻门密度最低的中心城区1.93倍.

|
图中相同字母表示浮游植物密度平均值差异显著(P < 0.05), 不同字母则表示平均值差异不显著(P>0.05) 图 4 中心城区、新城镇和农村地区秋季、夏季浮游植物密度 Fig. 4 Phytoplankton density in the central urban area, new town, and rural area during the autumn and summer |
各地区优势种无明显规律(表 1), 仍然是秋季各地区的微囊藻优势度最高, 而夏季优势种的优势度差异不显著, 农村地区水华微囊藻的优势度比较高, 为0.08, 其它优势种见表 1.
2.3 浮游植物分布与环境因子间的关系分别对全市河道秋季和夏季的浮游植物密度数据进行去趋势对应分析(DCA), 以选择合适的排序分析方法.DCA分析结果显示全市河道秋季、夏季DCA排序轴梯度最大值均小于3, 因此选用RDA分析, 并进行蒙特卡洛置换检验, 逐步筛选出对浮游植物群落特征具有显著性影响的环境因子(表 2)进行分析, 秋季全市河道筛选出SD、TP、TN和Turb等4个主要环境因子, 夏季筛选出TN、Turb、SD和pH等4个主要环境因子.物种数据与环境因子均呈显著强相关, 且各因子对物种数据的累计解释量都较高(表 3), 秋季、夏季筛选出的环境因子与排序轴1及所有排序轴分别为极显著相关(P < 0.01)和显著相关(P < 0.05), 秋季和夏季物种排序轴1与轴2的相关系数分别为-0.122和0.077, 相关性很低, 环境排序轴1与轴2相关系数均为0, 说明结果是可靠的[17].
|
|
表 2 各地区主要理化因子值 Table 2 Averages of the main physicochemical factors in each region |
|
|
表 3 主要浮游植物RDA分析统计信息 Table 3 Summary statistics for the axes of the RDA performed for major phytoplankton |
RDA分析结果(表 4)显示秋季SD与排序轴1呈最大正相关, TP与排序轴1呈最大负相关, Turb与排序轴2呈最大负相关; 夏季Turb与排序轴1呈最大正相关, SD与排序轴1呈最大负相关, TN与排序轴2呈最大正相关, pH与排序轴2呈最大负相关.秋季SD、Turb和TP是影响浮游植物群落的重要因子, 其次是TN; 夏季TN与Turb是影响浮游植物群落的重要因子, 其次是pH、SD.
|
|
表 4 RDA前2轴与环境因子之间相关系数 Table 4 Correlation coefficients between various environmental factors and the first two axis of the RDA |
从图 5(a)可以看出, 秋季新城镇样方聚集于第二、三象限, 与TN、TP靠近, 而农村地区样方主要聚集于第一、四象限, 与SD靠近, 中心城区样方分布较为分散, 影响因素复杂, 受人为干扰影响极大; 见图 5(b), 夏季新城镇和农村样方聚集于第一、四象限, 与TN、Turb靠近, 中心城区样方仍然较为分散, 影响因素复杂.

|
图中空白圆圈表示中心城区河道样方, 实心黑色正方形表示新城镇河道样方, 空白星形表示农村河道样方 图 5 浮游植物样方与环境因子RDA排序 Fig. 5 RDA biplot of phytoplankton sampling and environmental variables |
中心城区、新城镇和农村河道浮游植物的种类均是夏季种类数显著高于秋季, 而且3个地区河道的种类以绿藻最多, 其次为蓝藻, 这与李为星的研究结果一致[20].而夏季各地区浮游植物密度也高于秋季, 尤其是蓝藻门, 这可能与河道内浮游植物的群落结构、气候及周围环境的变化有关.调查河道秋季平均水温为(20.83±3.32)℃, 夏季平均水温为(29.94±1.95)℃, 夏季水温明显高于秋季, 而蓝藻均适宜在高温下大量繁殖, 一般蓝藻适宜生长的温度范围为25~35℃, 为耐高温藻类[21], 因而夏季水温的升高为蓝藻生长提供了良好的生长环境, 进而引起蓝藻暴发, 因此蓝藻种类数及密度均为夏季高于秋季.由于细胞分裂速度与光合作用产物C的供应有关, 所以细胞分裂速度取决于光强度[22].而夏季光线强度一般比秋季强, 进入水体真光层的深度也比秋季大, 可以促进浮游植物的光合作用, 有利于浮游植物的生长.N、P营养盐是浮游植物生长所必需的营养物质, 水体中N、P营养盐的浓度及比例会很大程度影响浮游植物的分布[23].夏季降雨量比较大, 陆域的营养盐随着雨水的冲刷进入河道内, 使营养盐浓度升高, 也为浮游植物的生长提供了良好环境.本研究结果显示夏季TN浓度高于秋季, 而TP的差异不大, 在营养盐方面, 上海市河道浮游植物的结构总体上是受TN限制的.这些因素随着季节的变化, 对浮游植物的群落结构有很大的影响.
3.2 浮游植物群落结构空间变化的影响因素中心城区人口密集, 且管理严格, 影响因素比较复杂, 多数河道采用半封闭式河道池塘生态修复技术, 不同的点位实际上有不同的影响因素, 而浮游植物群落对人为影响非常敏感[24], 要具体点位具体分析, 本文不着重讨论.而新城镇河道浮游植物群落主要受Turb、TN和TP的影响.新城镇河道因为周边既有居民区又有农田, 可能部分点位会接纳到生活污水和农业面源污染双重污染负荷, 尤其是农业面源的污染, 再加上夏季降雨频繁, 降雨量大, 陆域的污染负荷随着雨水一同进入新城镇河道内, 使TN和TP浓度产生了空间上的异质性, 成为限制新城镇河道浮游植物的主要影响因子, 其中TN浓度波动很大, 造成浮游植物分布的差异性. Turb一般与SD的关系呈负相关, RDA分析结果也同样呈现负相关关系, 这与Lee等[25]的报道一致.秋季SD与浮游植物密度呈显著正相关, 而在夏季则呈显著负相关.在浑浊的水体环境中, 悬浮颗粒物会改变光的水下传递, 导致水下光照不足从而影响浮游植物生长[26]. RDA分析结果表明, 农村河道浮游植物的群落结构主要与SD或Turb关系比较大.农村河道多为自然岸坡, 由于地表径流冲刷等因素的扰动, 使河道周边土壤或泥沙进入河道, 增加了河道悬浮物含量, 从而影响其Turb和SD的大小, 进而影响浮游植物群落结构.
本研究结果显示, 可以看到农村河道的TN、TP浓度均值在3个地区中虽然不是最高, 但是浮游植物密度却是最高的, 这是因为高浓度的浮游植物易形成藻华, 对营养盐反而有去除效果[27].而藻华是水体对过量营养物质的自我维持应急反应的表现[28], 可以看出农村河道富营养化问题十分严重, 上海需要重点关注农村河道的浮游植物分布情况.水动力是影响浮游植物生长的一个重要环境条件, 水工建筑物能显著地影响河流系统中浮游植物的生物量[29].水力停留时间长、流速慢即水动力差的水体, 可以为浮游植物提供稳定的生长环境, 浮游植物可以充分生长[30].农村地区河道与外界河流的交换是通过泵闸等设施, 交换频率较低, 导致农村地区河道水动力不足, 水体长期处于几乎静止的状态, 为浮游植物营造了一个相对舒适的环境.由于水体交换较少, 营养盐也会慢慢累积, 尤其是在夏季温度较高的条件下, 蓝藻容易暴发, 形成水华.后续可以针对水动力等水温因素开展深入研究.
调查发现秋季微囊藻是农村地区的主要优势种, 其优势度都是显著高于其它种类的; 而夏季农村地区水华微囊藻的优势度也是相对最大的.微囊藻是一类会向水体中释放微囊藻毒素的蓝藻[31], 会对饮用水、灌溉和娱乐用水健康造成危害[32].金泽水库位于上海市青浦区的农村地区, 其水源取自太浦河, 太浦河的水质对水库水质会有直接的影响.而本研究得出农村河道浮游植物的密度在3个地区中相对最大, 因此浮游植物尤其是像微囊藻这类蓝藻可能会在取水口聚集, 进而危害水源地源水健康.因此, 政府应加强农村地区尤其是水源地周边河道的浮游植物群落监控, 通过提高泵闸运作频率, 增加农村地区河道水动力从而减少浮游植物富集情况的发生, 以保障全市饮用水安全.
4 结论(1) 调查河道浮游植物种类数和平均密度均呈现夏季>秋季, 农村河道>新城镇河道>中心城区河道的规律.秋季、夏季的浮游植物种类数按从大到小排列都是绿藻、蓝藻和硅藻, 而秋季、夏季的浮游植物密度按从大到小排列都是蓝藻、硅藻和绿藻.
(2) 影响秋季和夏季浮游植物群落结构的主要因子分别为SD、TP、TN、Turb和TN、Turb、SD、pH.秋季, 新城镇河道主要受Turb、总氮和总磷的影响, 农村河道则主要受SD的影响; 夏季, 中心城区和农村河道主要受TN、Turb的影响.两次调查的中心城区浮游植物的影响因素均较为复杂.
(3) 应着重关注农村河道浮游植物水华和富营养化问题, 监测水源地附近河道优势种的变化, 尤其是对水体有害的优势种, 防止其污染水源地水源.
| [1] | Manickam N, Bhavan P S, Santhanam P, et al. Phytoplankton biodiversity in the two perennial lakes of Coimbatore, Tamil Nadu, India[J]. Acta Ecologica Sinica, 2020, 40(1): 81-89. |
| [2] | Pérez J R, Loureiro S, Menezes S, et al. Assessment of water quality in the Alqueva Reservoir (Portugal) using bioassays[J]. Environmental Science and Pollution Research, 2010, 17(3): 688-702. |
| [3] | Cui L, Lu X X, Dong Y L, et al. Relationship between phytoplankton community succession and environmental parameters in Qinhuangdao coastal areas, China:A region with recurrent brown tide outbreaks[J]. Ecotoxicology and Environmental Safety, 2018, 159: 85-93. |
| [4] | Kazamia E, Helliwell K E, Purton S, et al. How mutualisms arise in phytoplankton communities:building eco-evolutionary principles for aquatic microbes[J]. Ecology Letters, 2016, 19(7): 810-822. |
| [5] |
许海, 陈洁, 朱广伟, 等. 水体氮、磷营养盐水平对蓝藻优势形成的影响[J]. 湖泊科学, 2019, 31(5): 1239-1247. Xu H, Chen J, Zhu G W, et al. Effect of concentrations of phosphorus and nitrogen on the dominance of cyanobacteria[J]. Journal of Lake Sciences, 2019, 31(5): 1239-1247. |
| [6] |
胡旻琪, 张玉超, 马荣华, 等. 巢湖2016年蓝藻水华时空分布及环境驱动力分析[J]. 环境科学, 2018, 39(11): 4925-4937. Hu M Q, Zhang Y C, Ma R H, et al. Spatial and temporal dynamics of floating algal blooms in Lake Chaohu in 2016 and their environmental drivers[J]. Environmental Science, 2018, 39(11): 4925-4937. |
| [7] |
余茂蕾, 洪国喜, 朱广伟, 等. 风场对太湖梅梁湾水华及营养盐空间分布的影响[J]. 环境科学, 2019, 40(8): 3519-3529. Yu M L, Hong G X, Zhu G W, et al. Wind field influences on the spatial distribution of cyanobacterial blooms and nutrients in Meiliang Bay of Lake Taihu, China[J]. Environmental Science, 2019, 40(8): 3519-3529. |
| [8] |
罗宜富, 李磊, 李秋华, 等. 阿哈水库叶绿素a时空分布特征及其与藻类、环境因子的关系[J]. 环境科学, 2017, 38(10): 4151-4159. Luo Y F, Li L, Li Q H, et al. Spatial and temporal distribution of chlorophyll a and its relationship to algae and environmental factors in Aha Reservoir[J]. Environmental Science, 2017, 38(10): 4151-4159. |
| [9] |
余茂蕾, 洪国喜, 许海, 等. 湖泊蓝藻水华对连通河道水质的影响[J]. 环境科学, 2019, 40(2): 603-613. Yu M L, Hong G X, Xu H, et al. Effects of cyanobacterial blooms in eutrophic lakes on water quality of connected rivers[J]. Environmental Science, 2019, 40(2): 603-613. |
| [10] | Sun R, Sun P F, Zhang J H, et al. Microorganisms-based methods for harmful algal blooms control:a review[J]. Bioresource Technology, 2018, 248: 12-20. |
| [11] | Shi K, Zhang Y L, Qin B Q, et al. Remote sensing of cyanobacterial blooms in inland waters:present knowledge and future challenges[J]. Science Bulletin, 2019, 64(20): 1540-1556. |
| [12] |
孙博伟.滴水湖浮游植物优势种群演替及与外围水体的比较研究[D].上海: 上海师范大学, 2018. Sun B W. Succession of dominant species of phytoplankton and its comparison with peripheral water in Dishui Lake[D]. Shanghai: Shanghai Normal University, 2018. http://cdmd.cnki.com.cn/Article/CDMD-10270-1018211365.htm |
| [13] |
杨丽.淀山湖浮游植物功能群的时空演替特征及其与环境因子的关系[D].上海: 上海海洋大学, 2018. Yang L. Spatial and temporal succession characteristics of phytoplankton functional groups and their relationship with environmental factors in Dianshan Lake[D]. Shanghai: Shanghai Ocean University, 2018. https://kns.cnki.net/KCMS/detail/detail.aspx?dbcode=CMFD&filename=1018848906.nh |
| [14] |
严文逸, 王绍祥, 顾静, 等. 青草沙水库浮游藻类群落组成及其与环境因子的关系[J]. 上海海洋大学学报, 2017, 26(1): 75-84. Yan W Y, Wang S X, Gu J, et al. Composition of planktonic algae community and its relationship with environmental factors in Qingcaosha reservoir[J]. Journal of Shanghai Ocean University, 2017, 26(1): 75-84. |
| [15] |
陆倩, 崔冬, 田利勇, 等. 平原河网地区泵闸合建枢纽布置形式[J]. 水利水电科技进展, 2019, 39(3): 62-67. Lu Q, Cui D, Tian L Y, et al. Arrangement types of a combined sluice-pump hub in plain river network region[J]. Advances in Science and Technology of Water Resources, 2019, 39(3): 62-67. |
| [16] |
胡鸿钧, 魏印心. 中国淡水藻类——系统、分类及生态[M]. 北京: 科学出版社, 2006. Hu H J, Wei Y X. The freshwater algae of China:systematics, taxonomy and ecology[M]. Beijing: Science Press, 2006. |
| [17] |
林青, 由文辉, 徐凤洁, 等. 滴水湖浮游动物群落结构及其与环境因子的关系[J]. 生态学报, 2014, 34(23): 6918-6929. Lin Q, You W H, Xu F J, et al. Zooplankton community structure and its relationship with environmental factors in Dishui Lake[J]. Acta Ecologica Sinica, 2014, 34(23): 6918-6929. |
| [18] | Li C C, Feng W Y, Chen H Y, et al. Temporal variation in zooplankton and phytoplankton community species composition and the affecting factors in Lake Taihu-a large freshwater lake in China[J]. Environmental Pollution, 2019, 245: 1050-1057. |
| [19] | Noman M A, Sun J, Gang Q, et al. Factors regulating the phytoplankton and tintinnid microzooplankton communities in the East China Sea[J]. Continental Shelf Research, 2019, 181: 14-24. |
| [20] |
李为星.上海市生态建设河道浮游植物群落结构特征及其对水环境指示作用研究[D].上海: 上海海洋大学, 2013. Li W X. Study on the characters of phytoplankton community structure and its indication for aquatic environment of suburb rivers in Shanghai[D]. Shanghai: Shanghai Ocean University, 2013. http://cdmd.cnki.com.cn/Article/CDMD-10264-1014013543.htm |
| [21] |
方丽娟, 刘德富, 杨正健, 等. 水温对浮游植物群落结构的影响实验研究[J]. 环境科学与技术, 2014, 37(S2): 45-50. Fang L J, Liu D F, Yang Z J, et al. Effects of water temperature on the phytoplankton community structure[J]. Environmental Science and Technology, 2014, 37(S2): 45-50. |
| [22] | Thomas M K, Aranguren-Gassis M, Kremer C T, et al. Temperature-nutrient interactions exacerbate sensitivity to warming in phytoplankton[J]. Global Change Biology, 2017, 23(8): 3269-3280. |
| [23] | Huo S L, Zhang H X, Ma C Z, et al. Algae community response to climate change and nutrient loading recorded by sedimentary phytoplankton pigments in the Changtan Reservoir, China[J]. Journal of Hydrology, 2019, 571: 311-321. |
| [24] | Jones H F E, Özkundakci D, McBride C G, et al. Modelling interactive effects of multiple disturbances on a coastal lake ecosystem:implications for management[J]. Journal of Environmental Management, 2018, 207: 444-455. DOI:10.1016/j.jenvman.2017.11.063 |
| [25] | Lee Hestir E, Brando V E, Bresciani M, et al. Measuring freshwater aquatic ecosystems:the need for a hyperspectral global mapping satellite mission[J]. Remote Sensing of Environment, 2015, 167: 181-195. |
| [26] | Dodds W K. Eutrophication and trophic state in rivers and streams[J]. Limnology and Oceanography, 2006, 51(1): 671-680. |
| [27] | Peng Y K, Liu L, Jiang L J, et al. The roles of cyanobacterial bloom in nitrogen removal[J]. Science of the Total Environment, 2017, 609: 297-303. |
| [28] | Zhao W, Zheng Z, Zhang J L, et al. Evaluation of the use of eucalyptus to control algae bloom and improve water quality[J]. Science of the Total Environment, 2019, 667: 412-418. |
| [29] | Rao K, Zhang X, Yi X J, et al. Interactive effects of environmental factors on phytoplankton communities and benthic nutrient interactions in a shallow lake and adjoining rivers in China[J]. Science of the Total Environment, 2018, 619-620: 1661-1672. |
| [30] | Stumpner E B, Bergamaschi B A, Kraus T E C, et al. Spatial variability of phytoplankton in a shallow tidal freshwater system reveals complex controls on abundance and community structure[J]. Science of the Total Environment, 2020, 700: 134392. |
| [31] | Žegura B, Štraser A, Filipi Dč M. Genotoxicity and potential carcinogenicity of cyanobacterial toxins-a review[J]. Mutation Research/Reviews in Mutation Research, 2011, 727(1-2): 16-41. |
| [32] | Carvalho L, McDonald C, de Hoyos C, et al. Sustaining recreational quality of European lakes:minimizing the health risks from algal blooms through phosphorus control[J]. Journal of Applied Ecology, 2013, 50(2): 315-323. |