 2020, Vol. 41
2020, Vol. 41

 , CHEN Sheng-nan
, CHEN Sheng-nan , HUANG Ting-lin , JIA Jing-yu , LIU Kai-wen , MIAO Yu-tian , ZONG Rong-rong
, HUANG Ting-lin , JIA Jing-yu , LIU Kai-wen , MIAO Yu-tian , ZONG Rong-rong
随着地下水污染问题的加剧及城市地下水开采的严格限制, 水库已逐渐成为我国大部分城市的主要供水水源, 而水库作为重要的生态系统, 不仅能够为人类提供饮用水资源, 还具有农业灌溉、防洪和发电等多种重要的生态功能[1, 2].然而, 由于水库特殊的生态学特征, 如水体流动性差、水力停留时间长、季节性热分层明显等, 近年来, 水库水质污染问题日益严重, 尤其是水体富养化导致的藻类周期性大量繁殖.近年来, 国内如山东周村水库[1]、三峡水库[3]和厦门溪东水库[4]等, 以及国外如美国Barberton水库[5]、南非Malilangwe水库[6]和韩国Daechung水库[7]等均出现藻类暴发现象.在藻类大量繁殖的过程中, 不仅会大大降低水体透明度、消耗水中溶解氧以及破坏水生生态系统平衡等[1], 而且, 由于大部分藻类尤其是蓝藻, 能够释放毒性较强的藻毒素, 这些藻毒素不仅能够直接危及水生生物的健康安全, 且其通过这些水生生物在食物链不断向上传递富集后, 造成人体多种器官组织的破坏, 对人类的健康安全造成巨大威胁[8].因此, 有关水源水库生态系统健康演替规律研究迫在眉睫.
伴随藻华生消机制的不断深入研究, 人们认识到, 藻华的形成与水体中的微生物, 如细菌、真菌和浮游动物等具有紧密联系[9].而这些水体中的微生物作为初级生产力的主要贡献者, 在水生食物网、营养物质循环、全球生物化学循环和氧气释放等方面发挥着关键作用[10, 11].近年来, 大量研究结果表明, 微生物与形成藻华的物种之间的相互作用是调节微生物与藻类种群动态的重要因素[1, 12~24].浮游藻类能够为细菌等水体中的微生物提供氧气和生长所需的有机质, 而微生物则可以分解藻类难以直接利用的有机物, 为藻类生长提供生长必须的氮磷营养盐和必须的生长因子, 循环利用营养物质, 从而维持生态系统的平衡[12, 25, 26].除了这种互利共生关系外, 藻类和微生物之间还存在相互抑制、相互竞争和寄生等多种隐秘的生态学相互作用[20], 都会影响微生物群落结构组成.近年来, 大量研究都聚焦于研究细菌与浮游藻类的相互关系[1, 4, 5, 10~16, 18, 21~23, 26, 27].例如, Zhang等[1]利用高通量DNA测序技术, 在研究周村水库在藻类暴发和衰亡期间, 水体中细菌和真菌群落多样性的动态变化特征发现, 藻类暴发显著改变了水库中细菌和真菌的群落结构.Su等[12]对太湖水体中细菌群落组成的动态变化以及细菌群落组成如何受到蓝藻藻华和环境因素的影响进行了研究, 结果表明, α生物多样性指数与微囊藻的丰度以及藻毒素浓度呈显著负相关, 说明微囊藻及其藻毒素可能通过降低其生物多样性来影响细菌群落结构.Tada等[28]利用氨基酸组成和激发发射矩阵(EEM)分析方法, 研究了威氏海链藻(Thalassiosira weissflogi)和Heterocapsa triquetra胞内释放的溶解性有机物对浮游细菌群落结构及多样性的影响, 研究发现, 硅藻和甲藻体内释放的溶解性有机物中的特定氨基酸可能影响细菌群落结构和多样性的变化.然而, 分层型水源水库藻类演替的水质和水体细菌代谢和种群演替驱动机制尚不明确.
综上所述, 水源水库藻华暴发可能与某些特定的细菌群落结构存在着一一对应的关系.但是, 关于西安李家河水库藻类和细菌之间的相互作用及其垂向演替并未见相关报道.因此, 本实验以李家河水库为研究对象, 采用高通量DNA测序和Biolog技术分析了在李家河水库微囊藻暴发期间, 水质参数、浮游藻类及细菌群落结构在垂向上的演变特征, 并且通过主成分分析方法探究了细菌群落组成垂向演替的调控因子.主要目的为:①探明在微囊藻暴发期间, 水库中的主要水质参数的垂向演变规律; ②研究在藻类暴发期间, 浮游藻类群落结构的垂向变化特征; ③通过高通量测序技术研究细菌群落结构的垂向演替, 并通过主成分分析方法探究细菌群落结构同藻类和水质参数间的偶联关系.本文对水源水库藻类暴发期藻类与细菌群落的偶联机制进行探究, 以期为分层型水库水体藻华消涨的分子微生态驱动机制研究提供科学依据.
1 材料与方法 1.1 研究区域概况李家河水库(33°51′~34°08′N; 109°17′~109°36′E)位于陕西省西安市蓝田县境内, 是西安市浐河以东地区的重要水源工程.其建成于2015年, 属峡谷分层型水源水库, 总库容5.69×107 m3, 平均水深56 m, 最大水深80 m.伴随水库供水服役时间的延长, 水库生态系统呈现逐年富营养化趋势[29].
1.2 样品采集及水质参数测定本研究于2017年8月2日, 在李家河水库分别选取位于主库区的S1(33°59′28″N; 109°23′46″E)、S2(33°59′34″N; 109°23′55″E)和S3(33°59′17″N; 109°24′00″E)为采样点, 进行样品采集.采用直立式有机玻璃取样器, 分别在每个采样点的深度0、3和6 m处各采集1.5 L水样, 并在24 h内带回实验室进行各项水质指标的分析.其中, 水温(T)、pH、溶解氧(DO)、叶绿素a(Chla)、电导率(conductivity, Cond)、浊度(turbidity, Tur)使用美国HACH Hydro-Lab DS-5型多参数水质分析仪进行原位在线测定.总氮(TN)、硝氮(NO3--N)、氨氮(NH4+-N)、总磷(TP)、溶解性有机碳(DOC)、铁(Fe)和锰(Mn)等化学指标均参照文献[30]进行测定, 其中硝氮(NO3--N)和氨氮(NH4+-N)分别采用紫外分光光度法和纳氏试剂分光光度法(DR5000, 美国HACH)进行测定, 总氮(TN)和总磷(TP)于高压锅消解后(121℃, 30 min), 也根据紫外分光光度法进行测定, 溶解性有机碳(DOC)采用TOC分析仪(TOC-L CPN, 日本岛津)进行测定, 而铁(Fe)和锰(Mn)的浓度基于火焰原子吸收光谱法采用原子吸收光谱仪(ICE-3000, 美国赛默飞)进行测定.所有水质参数分析在48 h内完成(n=3).
1.3 浮游藻类计数及鉴定取500 mL混匀水样, 加1%酸性Lugol溶液保存, 沉淀48 h后, 使用真空抽滤装置将水样浓缩至10 mL.对浮游藻类计数前, 先充分混匀藻液, 然后用移液枪吸取100 μL到浮游生物计数框中, 盖上盖玻片, 待藻细胞沉降至计数框底部后, 在10×40倍显微镜下(CX31显微镜, 日本奥林巴斯公司)计数100个视野, 藻密度以104cells·L-1为单位来计, 每个样本计数两次(若2次计数的结果与平均值相差超过15%, 则需对其进行第3次计数), 然后取平均值为最终计数结果[30].藻类计算公式为:
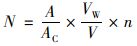
|
(1) |
式中, N为每升水体中浮游藻类个数(×104cells·L-1); A为计数框面积(mm2), 即400 mm2; AC为计数面积(mm2), 即视野面积×视野数; VW为水样经沉淀浓缩后的样品体积(mL), 即10 mL; V为计数框体积, 即0.1 mL; n为每片计数所得浮游藻类的细胞个数.
1.4 水体细菌群落代谢活性测定将20 mL原水倒入已灭菌的V型加液槽中, 采用8排电子移液枪(Bio-Rad, 美国)将液体加入ECO微平板(ECO Microplate, 美国Matrix Technologies公司)中, 每个微孔加入150 μL原水, 实验过程在超净工作台完成.随后将ECO板置于聚乙烯盒中, 放置在30℃的恒温培养箱中连续培养240 h, 每隔24 h测定微平板在590 nm处的吸光度值并绘制细菌群落代谢活性曲线, 即AWCD曲线[31].除此之外, 选取第120 h的吸光度值进行六大类碳源利用差异性分析、细菌群落多样性指数和多元统计分析.
ECO微平板含有96个微孔, 其中每32孔为一组, 共3组平行样, 每组的第一个孔为空白对照.该技术主要依据微生物在新陈代谢中产生的酶与四唑类物质(TTC和TV等)发生颜色反应的浊度差异从而检测微生物代谢特征指纹图谱.其中, 采样孔平均颜色变化率(AWCD)间接反映细菌群落结构的代谢活性, AWCD值越大反映细菌代谢活性越强.采样孔平均颜色变化率(AWCD590 nm)计算公式为[32]:
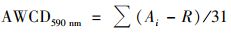
|
(2) |
式中, Ai为ECO微平板每一组中各孔在波长590 nm下的吸光度值, R为对照空白在波长590 nm下的吸光度, 若Ai-R≤0则记为0.群落丰富度指数(S)是指每组中AWCD值>0.2的孔数.香农多样性指数(H′)[32]:
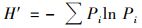
|
(3) |
式中, 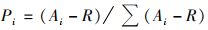
在超净工作台, 取500 mL水样, 用0.22 μm滤膜过滤, 然后对过滤在滤膜上的样品进行DNA的提取.采用CTAB/SDS方法, 利用DNA提取试剂盒(Water DNA Kit, Omega, 美国)来进行细菌总DNA的提取, 最后用2%琼脂糖凝胶电泳检测抽提的基因组DNA.
1.5.2 高通量DNA测序Illumina MiSeq高通量DNA测序在上海美吉生物公司进行, 测序平台为Illumina MiSeq PE300(Illumina, USA)[33].选取16S rRNA基因的高变区V3-V4区域, 运用细菌特异性引物338F-806R进行扩增.其中F端引物序列为ACTCCTACGGGAGGCAGCAG, R端引物序列为GGACTACHVGGGTWTCTAAT[1].所有细菌的DNA样品均使用PCR仪(ABI GeneAmp® 9700, CA, 美国)进行扩增, 且一式3份.PCR反应混合物(20 μL)由10 ng模板DNA、5 μmol·L-1正向和反向引物(0.8 μL)、5×反应缓冲液4 μL、2.5 mmol·L-1 dNTPs(2 μL)、快速Pfu DNA聚合酶(0.4 μL)、0.2 μL BSA和ddH2 O组成.而PCR反应包括98℃下初始变性3 min, 98℃下变性10 s, 55℃退火30 s, 72℃延伸45 s, 72℃延伸10 min, 共27个循环[34].
本研究中的所有高通量测序的16S rRNA基因序列数据均已上传至美国国立生物技术信息中心(NCBI-SRA)数据库中(网址:http://www.ncbi.nlm.nih.gov/), 编号:SRP174567.
1.6 数据处理与分析本研究使用Excel 2010和Origin 8.0对数据进行处理.使用SPSS对数据进行单因素方差分析(One-way ANOVA), 采用Tukey HSD检验, 显著性水平设定为0.01.采用CANOCO 4.5对各项指标进行冗余分析(PCA), 在分析之前, 除了pH值, 其余所有的水质指标均使用蒙特卡罗置换检验(P < 0.05)进行log(x+1)转换, 以使分析数据符合正态性和同方差性[1].使用微生物生态学定量分析(QIIME 2, 版本1.9.1)对Illumina MiSeq原始DNA序列进行处理和检验.经过组装和质量筛选后, 将具有97%及以上相似性的序列划分为同一分类单元(OTU).使用MOTHUR软件计算Chao丰富度指数(Chao 1)、Shannon多样性指数(H′)和Simpson多样性指数(D)[32].选择每个OTU最具代表性的序列, 然后使用RDP数据库为每个代表性序列注释分类信息[1, 27].
2 结果与讨论 2.1 水质分析李家河水库在2017年8月处于热分层期, 水库坝前区域靠近岸边, 含有大量外源有机物如腐殖质等, 且此时正处于夏季, 水温较高, 光照强度较大, 加之水体扰动小, 因此易产生藻华现象[35, 36].由表 1可知, 随着水深增加, 藻密度逐渐降低.其中, 表层0 m藻密度最高(F=82.48, P < 0.001), 主要是由于表层水体光照强度充足, 水温较高且DO浓度大.当水深大于3 m时, 受光照强度、水温及其DO等影响, 藻类生长受到一定抑制导致藻密度急剧下降[37].与此同时, Chla与藻密度呈现相同变化趋势(F=37.60, P < 0.001), 本研究表明Chla与藻密度呈正相关.随着水深不断增加, pH下降趋势较显著(F=112.56, P < 0.001).表层水体呈弱碱性主要是由于大量藻类进行光合作用打破了水体中碳酸盐平衡.此外, DO随深度变化, 呈现降低趋势, 尤其是在深度6 m处, DO降低幅度最大(F=714.50, P < 0.001), 由深度0 m处(6.41±0.22) mg·L-1迅速降至(0.12±0.00) mg·L-1, 这主要是受藻密度大幅度较低所致[2, 35].TN、NH4+-N和NO3--N随水深变化显著, 其中TN和NO3--N浓度随深度变化逐渐增加, 而NH4+-N则逐渐降低(F=26.87, P < 0.001), 该结果可能一方面受到藻密度显著变化的影响, 另一方面可能与此时水库水体处于热分层期存在一定关联[31].TP随水深以及藻密度变化并不显著, 除此之外Fe和Mn含量随深度变化也无明显差异, 这可能是因为此时藻密度相较于藻类暴发高峰期而言比较低, 作为藻类暴发前期, 其藻类仍处于生长期所导致[35].有研究发现, 在藻类暴发高峰期以及衰亡期时, 水体当中Fe的含量会发生剧烈升高, 这主要是因为Fe作为藻类生长所必需的的微量元素, 参与藻类的光合作用和呼吸作用等生命活动[38].并且, Zhang等[1]的研究发现, 在藻华前期, 水库中Fe的浓度一直处于最低值, 而在藻华衰退期时, Fe的浓度则突然显著增加(P < 0.001).Landa等[39]在研究浮游植物大量繁殖过程中, 与碳循环增加有关的细菌群落组成的变化时指出, 较高浓度的铁可以促进海洋环境中藻华的形成.
|
|
表 1 李家河水库不同水深水质参数1) Table 1 Spatial variation of water quality parameters in the Lijiahe Reservoir |
2.2 藻类群落结构
李家河不同水深处藻类群落结构(属水平)如图 1所示.随着水深的增加, 藻类群落结构发生显著变化.其中, 在深度0 m和3 m处, 微囊藻(Microcystis)占绝对优势, 其相对丰度在整个藻类群落结构中达93%以上.而在水深6 m处, 藻类群落结构发生显著改变, 微囊藻的优势地位被小环藻(Cyclotella)所取代, 其中小环藻的相对丰度为57.28%, 其次为微囊藻(23.30%), 除此之外, 针杆藻(Synedra)也作为优势藻之一出现, 相对丰度达7.44%.微囊藻隶属于蓝藻门, 是引起淡水藻华的常见藻种, 某些微囊藻会产生藻毒素, 毒性较强, 能够抑制水生生物的生长[13, 17, 40, 41].微囊藻在高温及较高的光照强度下可快速生长, 而在低温条件下其生长则会受到抑制[8].并且在氮营养盐浓度较低的情况下, 微囊藻仍可大量生长繁殖, 这主要是由于蓝藻自身具有固氮的能力, 就算水体中的氮源不充足, 也能够通过固定空气中的氮气(N2)来满足自身对氮源的需求[42, 43].因此, 在夏季, 由于天气晴朗、温度较高且水面较为平静, 充分满足了微囊藻的生长条件, 故易暴发微囊藻藻华, 而随着全球气候变暖, 藻华的持续时间通常延长至秋季[44, 45].此外, 在水深3 m处, 虽然其藻密度较水深0 m减少了51.31%, 但微囊藻仍为绝对优势藻种, 说明微囊藻在水深0 m至3 m均能较好生长.而在水深6 m处, 硅藻门的小环藻取代微囊藻, 成为该水层相对丰度最大的优势藻种.与微囊藻相比, 小环藻对温度的适应范围更加广泛, 并且在高温下小环藻的光能利用效率高[46, 47].王敏等[47]对嘉陵江重庆出口段长期监测发现, 该地在每年1月下旬至3月上旬期间会发生小环藻暴发, 在暴发期间, 小环藻相对丰度达85% ~94%, 且有研究表明春季水温适宜小环藻生长, 较低的温度和水体扰动以及适宜的光照强度均促进小环藻的暴发.而Yang等[48]在研究亚热带水库浮游藻类群落结构演替的外部和内部因子时发现, 针杆藻(Synedra)的数量在8~9月期间达到最大, 占总藻密度的36%, 这主要是因为针杆藻相比于其他藻种对有效的水下光环境恶化具有耐受性.

|
图 1 李家河水库不同水深藻类群落结构 Fig. 1 Phytoplankton community structure at different water depths in the Lijiahe Reservoir |
李家河水库不同水深处细菌群落代谢活性用AWCD590 nm曲线来表征, 如图 2所示.从中可知, 在不同水深处, 细菌代谢活性呈现一定的差异性.在水深6 m处, AWCD590 nm值为最大, 而水深0 m和3 m处的AWCD590 nm无显著区别.由于AWCD590 nm曲线可间接表征细菌的代谢活性, 因此结果表明, 水深6 m处的细菌代谢活性最大, 其次为水深3 m和0 m的细菌群落.这一结果与Yang等[31]的研究结果相似, 他们发现在水体热分层期间, 随着水深增加, 细菌代谢活性也逐渐升高.这可能是由于微囊藻释放藻毒素抑制细菌生长, 并且微囊藻仍处于生长期, 会争夺水体中大部分营养物质, 并且, 聚集在表层的大量微囊藻严重影响了水体透光率, 进而对细菌群落代谢活性也会产生不利影响.但也有研究发现, 随着藻密度增大, 细菌群落整体代谢活性会升高[49], 这可能是由于地理位置以及微生物群落结构的差异所造成.Zhang等[32]在研究不同饮用水水库沉积物中细菌和真菌的群落功能多样性时发现, 细菌和真菌群落的对碳源的利用模式在不同水库之间存在显著差异.

|
图 2 李家河水库不同水深细菌群落AWCD590 nm曲线 Fig. 2 AWCD590 nm curves of bacterial communities at different depths of the Lijiahe Reservoir |
在使用Biolog技术测定细菌代谢活性时, 选取培养第120 h的吸光度进行细菌群落对不同碳源利用的差异性分析、细菌群落丰富度指数和香农多样性指数分析, 如表 2所示.从中可知, 在不同水深处, 细菌群落对氨基酸类、糖类和胺类的利用无显著差异, 但其对酚类碳源的利用却呈现显著差异, 相比于水深0 m和6 m, 细菌群落对酚类碳源利用率在水深3 m处显著降低.就物种丰富度而言, 随着水深增加, 细菌群落丰度逐渐增加, 但细菌群落的香农多样性指数(H′)并无显著差异(表 2).综上所述, 微囊藻的暴发对细菌代谢活性及其丰度产生较大影响, 但不改变细菌多样性的变化, 并且在水深6 m处细菌代谢活性和丰度均达到最大.有研究发现, 处于不同水层处的细菌对糖类、氨基酸类、酯类和醇类的利用呈现出显著差异, 其中位于混合层的细菌对氨基酸类、醇类、胺类和羧酸类的利用率最高, 而变温层的细菌对糖类、氨基酸类、酯类和醇类的利用率最低, 并且与本文研究结果类似, 他们还发现香农多样性指数在不同水层间的变化不明显, 即随深度变化无显著差异[31].
|
|
表 2 基于Biolog微生物群落水平生理图谱对细菌群落第120 h的代谢参数分析1) Table 2 Metabolic parameters of the bacterial community at 120 h based on Biolog microbial community level |
2.4 细菌群落结构多样性分析 2.4.1 多样性指数分析
对李家河水库不同水深处的细菌群落进行α多样性分析, 其中包括ACE、Chao 1、Shannon多样性指数(H′)和Simpson多样性指数(D)分析, 其覆盖率均达到99%以上, 说明数据可靠(表 3).ACE和Chao 1反映样本之间的丰富度, 其值越大, 则表明细菌群落丰富度越高[1].而Shannon多样性指数和Simpson多样性指数均表征微生物群落多样性, 其中Shannon多样性指数越高, 表明细菌群落多样越大, 而Simpson多样性指数刚好与之相反, 当其值越高时, 表明细菌群落多样越小[2].由表 3可知, Chao 1丰富度指数均高于细菌的OTU值, 说明在李家河水库还有很多未知序列的细菌有待进一步研究[31, 50, 51].在水深6 m处, ACE、Chao 1和Shannon多样性指数均达到最大值, 而在水深3 m处, 细菌群落、Chao 1和Shannon多样性指数最低, 并且不同水深处的Simpson多样性指数在0.02~0.03之间波动, 无显著差异.结合细菌群落代谢活性和多样性分析来看, 在水深6 m处, 随着藻密度的大幅度降低及藻类群落结构的显著改变, 细菌数量升至最高, 其群落的生物多样性也达到最大, 并且在该水深处, 细菌对不同碳源的代谢活性最高, 而在水深3 m处, 虽然藻密度较水深0 m也有显著降低的趋势, 但由于藻类群落结构并未发生改变, 仍以微囊藻为主, 故其细菌数量、群落多样性及碳源代谢活性均处于较低水平.以上结果表明, 藻类数量及群落结构的改变对细菌丰度、生物活性及群落多样性均具有显著影响.有研究发现, 水体微生物群落Chao 1指数随深度的不同而呈现出显著差异, Shannon多样性指数(H′)在水库底层较高[50].Yang等[31]通过研究热分层时期细菌群落的动态特征发现恒温层中的细菌群落多样性显著高于变温层的细菌群落多样性, 热分层能够显著改变水生生态系统中微生物的群落结构.
|
|
表 3 李家河水库不同水深细菌群落多样性指数分析 Table 3 Bacterial community diversity and richness estimators at different water depths of the Lijiahe Reservoir |
2.4.2 细菌群落结构组成分析
在微囊藻暴发期间, 李家河水库不同水深处门水平的细菌群落呈现显著差异.如图 3所示, 高通量测序所得到的具有97%及以上相似性的1 420个OTU隶属于10个细菌门类, 分别为放线菌门(Actinobacteria)、蓝细菌门(Cyanobacteria)、变形菌门(Proteobacteria)、拟杆菌门(Bacteroidetes)、疣微菌门(Verrucomicrobia)、绿菌门(Chlorobi)、浮霉菌门(Planctomycetes)、绿弯菌门(Chloroflexi)、厚壁菌门(Firmicutes)和芽单胞菌门(Gemmatimonadetes).本文中将蓝细菌门作为藻类, 因此不对其进行细菌群落分析.在微囊藻暴发期间, 放线菌门和变形菌门在细菌群落结构中占有较大比例, 二者相对之和达50%以上.其中, 放线菌门在不同深度处的相对丰度均高于变形菌门, 并且在水深3 m处, 放线菌门的相对丰度最大为43.86%, 而变形菌门与之相反, 在该深度相对丰度最小为12.53%.拟杆菌门、疣微菌门和绿弯菌门垂向分布呈现明显规律, 随着水深增加, 拟杆菌门和疣微菌门的相对丰度逐渐升高, 而绿弯菌门则依次降低.绿菌门和浮霉菌门的相对丰度随着水深增加呈现显著增加趋势(P < 0.001), 其中绿菌门由0.94%(0 m)升至10.43%(6 m), 浮霉菌门由2.11%(0 m)升至6.88%(6 m), 并且与藻密度呈显著负相关(P < 0.05).厚壁菌门(Firmicutes)和芽单胞菌门(Gemmatimonadetes)作为稀有藻属, 虽在细菌群落中占比较低, 但其相对丰度随着藻密度降低而逐渐减小, 与藻密度呈显著正相关关系(P < 0.05).

|
字母A~J分别代表Actinobacteria、Cyanobacteria、Proteobacteria、Bacteroidetes、Verrucomicrobia、Chloroflexi、Planctomycetes、Chlorobi、Firmicutes、Gemmatimonadetes 图 3 李家河水库不同水深细菌门水平群落结构Circos分析 Fig. 3 Circos analysis of the bacterial community structure at the phylum level in different water depths of the Lijiahe Reservoir |
以上结果说明, 李家河水库在藻类暴发期间, 细菌群落变化特征明显, 且细菌与藻类之间均存在正相关或负相关关联, 这表明细菌与微囊藻暴发存在紧密联系, 可能会在藻华的形成过程中起到关键作用.其中, 藻类和细菌之间的正相关关系意味着二者之间互利共生, 占据相同的生态位, 且细菌对藻类暴发可能起到促进作用, 而负相关关系则表明藻类同细菌之间可能存在竞争或相互抑制或寄生等拮抗作用[41].Liu等[4]在研究微囊藻藻华期间浮游细菌和附着细菌的群落动态时发现, 藻华的发生使得放线菌门、拟杆菌门、厚壁菌门和γ-变形杆菌门的生活方式发生显著改变, 并且发现相比于浮游细菌, 附着细菌与藻类之间存在更多的显著正相关, 且相关系数很高, 表明这些细菌对藻华的发生具有一定的促进作用.Yang等[31]的研究发现在蓝藻暴发期间, 放线菌门成为主导细菌门类, 而在水库热分层期间, 拟杆菌门和浮霉菌门的相对丰度则有所降低.Ji等[52]对3个富营养湖泊的研究发现, 在藻类暴发期, 放线菌门、变形菌门、拟杆菌门以及浮霉菌是主要的优势细菌门类.Zhang等[1]探究了周村水库暴发蓝藻藻华的前期、中期和衰亡期间细菌群落结构演替规律, 发现在藻华期间, 放线菌门(23.18%)、变形菌门(22.77%)、拟杆菌门(8.33%)及厚壁菌门(4.71%)为主要的细菌门类, 并且在藻密度降低时, 放线菌门由23.18%升高至36.54%.Wei等[53]采用分子生物学方法, 对中国东部黄前水库的浮游细菌群落进行研究时发现, β-变形菌、拟杆菌门、放线菌门及γ-变形菌是该水库细菌群落中主要组成门类.
为了进一步研究微囊藻暴发对细菌群落结构的影响, 用热图分析了不同水深处细菌群落属水平相对丰度位于前49位的代表性物种, 如图 4所示.与门水平的变化特征相一致, 李家河水库属水平细菌群落结构在垂向分布上呈现显著差异, 随着水深的增加, 藻密度持续降低, 属水平的细菌群落分布更均匀且趋于多样性.其中, 聚球藻属(Synechococcus)和微囊藻属(Microcystis)属于蓝藻, 在此不作讨论.新鞘脂菌属(Novosphingobium)和Fluviicola在深度0 m处相对丰度最大, 分别为5.64%和5.45%, 并且伴随藻密度的降低, 其相对丰度也呈下降趋势, 这意味着这两种菌属对于藻类生长可能起到促进作用, 与藻华的形成具有紧密联系.Hahn等[54]在研究中也检测到了Fluviicola, 并发现其与其他菌属如芽孢杆菌属(Bacillus)和硝化螺菌属(Nitrospira)在较深水层处丰度最高.而玫瑰弯菌属(Roseiflexus)与藻密度呈显著负相关(P < 0.05), 其相对丰度随藻密度降低而依次增加, 且在水深6 m处玫瑰弯菌属相对丰度达到最大值10.61%, 这表明玫瑰弯菌属对藻类暴发可能产生抑制作用.玫瑰单胞菌属(Roseomonas)在水深0 m和3 m处占优, 最大相对丰度为5.86%, 与藻密度呈显著正相关(P < 0.05).有研究发现玫瑰单胞菌属(Roseomonas)和鞘氨醇单胞菌属(Sphingomonas)均具有溶藻特性[55].而Magnetospirillum、Terrimicrobium和Sediminibacterium只在水深6 m处占优, 相对丰度分别为7.11%、5.95%和5.54%, 而在其他深度所占比例均不足2%, 说明上述3种菌属为水深6 m处的特有优势藻属, 且与藻密度呈反比.Limnohabitans作为水生生态系统中最丰富的类群之一, 其也在水深6 m处相对丰度达最大, 值为4.54%.Limnohabitans属于β-变形菌, 具有在藻类基质上生长的能力, 且其对基质具有较高的吸收率, 这使得其在生态系统中具有独特的竞争优势, 在淡水浮游生物群落中起着关键作用[31].玫瑰单胞菌属(Roseomonas)、鞘脂菌属(Sphingobium)和Limnohabitans等在湖泊或水库中普遍存在[1, 10, 12, 56, 57].

|
图 4 李家河水库不同水深细菌群落结构在属水平热图分析 Fig. 4 Color-scale heat map of bacterial communities at the genus level at different water depths of the Lijiahe Reservoir |
为了更好地了解水质对细菌和藻类群落结构的影响, 通过冗余分析研究了细菌和藻类群落结构同水质指标之间的偶联关系.由图 5可知, 所选环境因子共解释了细菌和藻类群落结构的100%的变异, 其中RDA1轴解释89.3%, RDA2轴解释10.7%.在水深0、3和6 m处的细菌和藻类的群落结构受不同水质指标调控, 且差异显著.在水深0 m处, 细菌和藻类群落结构与温度(T)、溶解氧(DO)、pH、浊度(Tur)、TP、氨氮(NH4+-N)和叶绿素a(Chla)显著相关, 且这些水质指标尤其对藻类群落中微囊藻属(Microcystis)和细菌群落中放线菌门(Actinobacteria)以及疣微菌门(Verrucomicrobia)具有较大影响, 对上述细菌和藻类具有促进作用.而在水深3 m处, 溶解性有机碳(DOC)和锰(Mn)对细菌和藻类群落结构影响显著, 其中绿弯菌门(Chloroflexi)与其呈显著正相关(P < 0.01), 而栅藻(Scenedesmus)与其呈显著负相关(P < 0.01).在水深6 m处, 总氮(TN)、硝氮(NO3--N)、电导率(Cond)和铁(Fe)是细菌和藻类群落结构的关键调控因子, 且与绿菌门(Chlorobi)、浮霉菌门(Planctomycetes)、小球藻属(Chlorella)、针杆藻属(Synedra)和小环藻属(Cyclotella)呈显著正相关(P < 0.01).以上结果表明, 李家河水库细菌和浮游藻类的群落结构受环境因子调控特征明显, 其群落结构能够响应环境因子发生显著改变.Zhang等[1]通过RDA分析发现, 在藻华前后, 细菌群落结构与电导率(P < 0.05)和氨氮(P < 0.01)显著相关, 表明在藻类暴发前后细菌群落结构主要受电导率和氨氮调控.Wang等[5]在研究淡水水库中蓝藻藻华对调控浮游细菌群落组装的机制时发现, 浮游细菌群落的时间分布在很大程度上受温度、电导率、Chla、DON、TOC和HCO3-来调控, 且在非水华时期, 相同的环境变量可以解释更高的浮游细菌群落变化, 而浮游藻类群落的时间分布受温度、电导率、pH、NH4+、NOx、DON、SRP和TOC影响显著, 占观察到的变异的21.6% ~52.8%.Su等[27]发现水体中浮游植物和浮游细菌均受环境条件变化的影响, 其中水温和氮是影响浮游植物和浮游细菌的主要因素.冗余分析结果表明, 环境参数分别解释了浮游植物和浮游细菌51.0%的和46.7%的变异.谢丽娟等[58]发现在微囊藻藻华衰亡阶段, 浮游细菌群落组成与pH、NO3--N、浮游藻类的生物量密切相关, 而附着细菌群落除了受pH和浮游藻类生物量影响外, 还与DO和TP呈显著相关.Bell等[59]认为浮游藻类的大规模暴发会提高对水体中磷营养盐的吸收, 从而限制其他微生物对磷的利用.Wei等[53]则发现在黄前水库表层水体中, NO3--N和DO是影响细菌群落结构的关键因素, 冗余分析表明NO3--N、DO、pH等与大多数细菌群落呈负相关.

|
红色三角和红色字体表示浮游藻类, 蓝色三角和和蓝色字体表示细菌 图 5 李家河水库不同水深细菌和藻类群落同水质因子的冗余分析 Fig. 5 Redundancy analysis of the bacterial and algal community with water quality at different depths of the Lijiahe Reservoir |
(1) 李家河水库水质指标在垂向分布呈现显著差异.随水深增加, 藻密度和Chla呈现同步降低趋势(P < 0.001).pH随深度增加其下降趋势显著(P < 0.001), DO从(6.41±0.22) mg·L-1(0 m)迅速降至(0.12±0.00) mg·L-1(6 m), 与藻密度显著降低相关.TN和NO3--N浓度随深度逐渐增加, 而NH4+-N则逐渐降低(P < 0.001).
(2) 随水库水体深度增加, 藻类群落结构变化显著.其中水深0 m和3 m以微囊藻为绝对优势藻属, 其相对丰度均为90%以上, 而在水深6 m处, 小环藻取代微囊藻成为最优藻属, 相对丰度达57.28%, 且针杆藻比例明显增大, 其相对丰度由表层的0.42%增至7.44%.
(3) 微囊藻的暴发对细菌代谢活性及其相对丰度产生较大影响, 但不改变细菌多样性的变化.细菌代谢活性在水深6 m处最大, 其次为3 m和0 m.在水深3 m处, 细菌群落对酚类碳源利用率显著降低.细菌群落丰度随水深增加呈增大趋势, 但其多样性并无显著差异.
(4) 放线菌门(Actinobacteria)和变形菌门(Proteobacteria)在藻类暴发期占较大比例, 其相对丰度之和达50%以上.绿菌门(Chlorobi)和浮霉菌门(Planctomycetes)与藻密度呈显著负相关(P < 0.05), 且相对丰度均在水深6 m达最大, 分别为10.29%和6.78%.随藻密度下降, 细菌群落分布更均匀且趋于多样性.新鞘脂菌属(Novosphingobium)等与藻密度呈显著正相关(P < 0.05), 而玫瑰弯菌属(Roseiflexus)等则与藻密度呈反比.
(5) 在垂向分布上, 细菌和藻类的群落结构受不同水质指标调控且差异显著, 这些水质指标对细菌和藻类群落的演替起到关键作用.在水深0 m处, 细菌和藻类群落结构与T、DO、pH、浊度、TP、NH4+-N和Chla显著相关.而DOC和Mn对水深3 m处的细菌和藻类群落结构影响较为显著.水深6 m处的细菌和藻类群落主要受到TN、NO3--N、Cond和Fe的调控.
| [1] | Zhang H H, Jia J Y, Chen S N, et al. Dynamics of bacterial and fungal communities during the outbreak and decline of an algal bloom in a drinking water reservoir[J]. International Journal of Environmental Research and Public Health, 2018, 15(2): 361. DOI:10.3390/ijerph15020361 |
| [2] | 商潘路.水源水库水质及浮游藻类种群结构时空演替规律研究[D].西安: 西安建筑科技大学, 2018. http://cdmd.cnki.com.cn/Article/CDMD-10703-1018861407.htm |
| [3] |
周川, 蔚建军, 付莉, 等. 三峡库区支流澎溪河水华高发期环境因子和浮游藻类的时空特征及其关系[J]. 环境科学, 2016, 37(3): 873-883. Zhou C, Wei J J, Fu L, et al. Temporal and spatial distribution of environmental factors and phytoplankton during algal bloom season in Pengxi River, Three Gorges Reservoir[J]. Environmental Science, 2016, 37(3): 873-883. |
| [4] | Liu M, Liu L M, Chen H H, et al. Community dynamics of free-living and particle-attached bacteria following a reservoir Microcystis bloom[J]. Science of the Total Environment, 2019, 660: 501-511. DOI:10.1016/j.scitotenv.2018.12.414 |
| [5] | Wang K, Razzano M, Mou X Z. Cyanobacterial blooms alter the relative importance of neutral and selective processes in assembling freshwater bacterioplankton community[J]. Science of the Total Environment, 2020, 706: 135724. DOI:10.1016/j.scitotenv.2019.135724 |
| [6] | Dalu T, Wasserman R J. Cyanobacteria dynamics in a small tropical reservoir:understanding spatio-temporal variability and influence of environmental variables[J]. Science of the Total Environment, 2018, 643: 835-841. DOI:10.1016/j.scitotenv.2018.06.256 |
| [7] | Oh H M, Ahn C Y, Lee J W, et al. Community patterning and identification of predominant factors in algal bloom in Daechung Reservoir (Korea) using artificial neural networks[J]. Ecological Modelling, 2007, 203(1-2): 109-118. DOI:10.1016/j.ecolmodel.2006.04.030 |
| [8] | Mello F D, Braidy N, Marçal H, et al. Mechanisms and effects posed by neurotoxic products of cyanobacteria/microbial eukaryotes/dinoflagellates in algae blooms:a review[J]. Neurotoxicity Research, 2018, 33(1): 153-167. DOI:10.1007/s12640-017-9780-3 |
| [9] | Van Wichelen J, Vanormelingen P, Codd G A, et al. The common bloom-forming cyanobacterium Microcystis is prone to a wide array of microbial antagonists[J]. Harmful Algae, 2016, 55: 97-111. DOI:10.1016/j.hal.2016.02.009 |
| [10] | Isabwe A, Yang J R, Wang Y M, et al. Community assembly processes underlying phytoplankton and bacterioplankton across a hydrologic change in a human-impacted river[J]. Science of the Total Environment, 2018, 630: 658-667. DOI:10.1016/j.scitotenv.2018.02.210 |
| [11] | Zhao D Y, Shen F, Zeng J, et al. Network analysis reveals seasonal variation of co-occurrence correlations between cyanobacteria and other bacterioplankton[J]. Science of the Total Environment, 2016, 573: 817-825. DOI:10.1016/j.scitotenv.2016.08.150 |
| [12] | Su X M, Steinman A D, Tang X M, et al. Response of bacterial communities to cyanobacterial harmful algal blooms in Lake Taihu, China[J]. Harmful Algae, 2017, 68: 168-177. DOI:10.1016/j.hal.2017.08.007 |
| [13] | Wilhelm S W, Farnsley S E, LeCleir G R, et al. The relationships between nutrients, cyanobacterial toxins and the microbial community in Taihu (Lake Tai), China[J]. Harmful Algae, 2011, 10(2): 207-215. DOI:10.1016/j.hal.2010.10.001 |
| [14] | Li H B, Xing P, Chen M J, et al. Short-term bacterial community composition dynamics in response to accumulation and breakdown of Microcystis blooms[J]. Water Research, 2011, 45(4): 1702-1710. DOI:10.1016/j.watres.2010.11.011 |
| [15] | Berry M A, Davis T W, Cory R M, et al. Cyanobacterial harmful algal blooms are a biological disturbance to Western Lake Erie bacterial communities[J]. Environmental Microbiology, 2017, 19(3): 1149-1162. DOI:10.1111/1462-2920.13640 |
| [16] | Parulekar N N, Kolekar P, Jenkins A, et al. Characterization of bacterial community associated with phytoplankton bloom in a eutrophic lake in South Norway using 16S rRNA gene amplicon sequence analysis[J]. PLoS One, 2017, 12(3): e0173408. DOI:10.1371/journal.pone.0173408 |
| [17] | Scherer P I, Millard A D, Miller A, et al. Temporal dynamics of the microbial community composition with a focus on toxic cyanobacteria and toxin presence during harmful algal blooms in two South German lakes[J]. Frontiers in Microbiology, 2017, 8: 2387. DOI:10.3389/fmicb.2017.02387 |
| [18] | Liu M, Xiao T, Sun J, et al. Bacterial community structures associated with a natural spring phytoplankton bloom in the Yellow Sea, China[J]. Deep Sea Research Part Ⅱ:Topical Studies in Oceanography, 2013, 97: 85-92. DOI:10.1016/j.dsr2.2013.05.016 |
| [19] | Sun J Y, Song Y, Ma Z P, et al. Fungal community dynamics during a marine dinoflagellate (Noctiluca scintillans) bloom[J]. Marine Environmental Research, 2017, 131: 183-194. DOI:10.1016/j.marenvres.2017.10.002 |
| [20] | Tan S J, Zhou J, Zhu X S, et al. An association network analysis among microeukaryotes and bacterioplankton reveals algal bloom dynamics[J]. Journal of Phycology, 2015, 51(1): 120-132. DOI:10.1111/jpy.12259 |
| [21] | Teeling H, Fuchs B M, Becher D, et al. Substrate-controlled succession of marine bacterioplankton populations induced by a phytoplankton bloom[J]. Science, 2012, 336(6081): 608-611. DOI:10.1126/science.1218344 |
| [22] | Guo Y Y, Liu M, Liu L M, et al. The antibiotic resistome of free-living and particle-attached bacteria under a reservoir cyanobacterial bloom[J]. Environment International, 2018, 117: 107-115. DOI:10.1016/j.envint.2018.04.045 |
| [23] | Xiao X, Pei M, Liu X, et al. Planktonic algal bloom significantly alters sediment bacterial community structure[J]. Journal of Soils and Sediments, 2017, 17(10): 2547-2556. DOI:10.1007/s11368-017-1691-7 |
| [24] | Jia Y, Han G M, Wang C Y, et al. The efficacy and mechanisms of fungal suppression of freshwater harmful algal bloom species[J]. Journal of Hazardous Materials, 2010, 183(1-3): 176-181. DOI:10.1016/j.jhazmat.2010.07.009 |
| [25] | Naeem S, Hahn D R, Schuurman G. Producer-decomposer co-dependency influences biodiversity effects[J]. Nature, 2000, 403(6771): 762-764. DOI:10.1038/35001568 |
| [26] | Niu Y, Shen H, Chen J, et al. Phytoplankton community succession shaping bacterioplankton community composition in Lake Taihu, China[J]. Water Research, 2011, 45(14): 4169-4182. DOI:10.1016/j.watres.2011.05.022 |
| [27] | Su X M, Steinman A D, Xue Q J, et al. Temporal patterns of phyto- and bacterioplankton and their relationships with environmental factors in Lake Taihu, China[J]. Chemosphere, 2017, 184: 299-308. DOI:10.1016/j.chemosphere.2017.06.003 |
| [28] | Tada Y, Nakaya R, Goto S, et al. Distinct bacterial community and diversity shifts after phytoplankton-derived dissolved organic matter addition in a coastal environment[J]. Journal of Experimental Marine Biology and Ecology, 2017, 495: 119-128. DOI:10.1016/j.jembe.2017.06.006 |
| [29] |
温成成, 黄廷林, 李楠, 等. 人工强制混合充氧及诱导自然混合对水源水库水质改善效果分析[J]. 环境科学, 2020, 41(3): 1227-1235. Wen C C, Huang T L, Li N, et al. Effects of artificial destratification and induced-natural mixing on water quality improvement in a drinking water reservoir[J]. Environmental Science, 2020, 41(3): 1227-1235. |
| [30] | 国家环境保护总局. 水和废水监测分析方法[M]. (第四版). 北京: 中国环境科学出版社, 2002. |
| [31] | Yang X, Huang T L, Zhang H H. Effects of seasonal thermal stratification on the functional diversity and composition of the microbial community in a drinking water reservoir[J]. Water, 2015, 7(10): 5525-5546. DOI:10.3390/w7105525 |
| [32] | Zhang H H, Huang T L, Chen S N, et al. Microbial community functional diversity and enzymatic activity in the sediments of drinking water reservoirs, Northwest China[J]. Desalination and Water Treatment, 2014, 52(7-9): 1608-1614. DOI:10.1080/19443994.2013.827383 |
| [33] | Xue Y Y, Chen H H, Yang J R, et al. Distinct patterns and processes of abundant and rare eukaryotic plankton communities following a reservoir cyanobacterial bloom[J]. The ISME Journal, 2018, 12(9): 2263-2277. DOI:10.1038/s41396-018-0159-0 |
| [34] | Liu L M, Liu M, Wilkinson D M, et al. DNA metabarcoding reveals that 200-μm-size-fractionated filtering is unable to discriminate between planktonic microbial and large eukaryotes[J]. Molecular Ecology Resources, 2017, 17(5): 991-1002. DOI:10.1111/1755-0998.12652 |
| [35] | Huang Y W, Yang C, Wen C C, et al. S-type dissolved oxygen distribution along water depth in a canyon-shaped and algae blooming water source reservoir:reasons and control[J]. International Journal of Environmental Research and Public Health, 2019, 16(6): 987. DOI:10.3390/ijerph16060987 |
| [36] | Kosakowska A, Nędzi M, Pempkowiak J. Responses of the toxic cyanobacterium Microcystis aeruginosa to iron and humic substances[J]. Plant Physiology and Biochemistry, 2007, 45(5): 365-370. DOI:10.1016/j.plaphy.2007.03.024 |
| [37] | Shi X L, Li S N, Zhang M, et al. Temperature mainly determines the temporal succession of the photosynthetic picoeukaryote community in Lake Chaohu, a highly eutrophic shallow lake[J]. Science of the Total Environment, 2020, 702: 134803. DOI:10.1016/j.scitotenv.2019.134803 |
| [38] | Zhang T, He J, Luo X Z. Effect of Fe and EDTA on freshwater cyanobacteria bloom formation[J]. Water, 2017, 9(5): 326. DOI:10.3390/w9050326 |
| [39] | Landa M, Blain S, Christaki U, et al. Shifts in bacterial community composition associated with increased carbon cycling in a mosaic of phytoplankton blooms[J]. The ISME Journal, 2015, 10(1): 39-50. |
| [40] | Wiltsie D, Schnetzer A, Green J, et al. Algal blooms and cyanotoxins in Jordan lake, North Carolina[J]. Toxins, 2018, 10(2): 92. DOI:10.3390/toxins10020092 |
| [41] | Chen M J, Chen F Z, Xing P, et al. Microbial eukaryotic community in response to Microcystis spp. bloom, as assessed by an enclosure experiment in lake Taihu, China[J]. FEMS Microbiology Ecology, 2010, 74(1): 19-31. DOI:10.1111/j.1574-6941.2010.00923.x |
| [42] | Xu H, Paerl H W, Qin B Q, et al. Nitrogen and phosphorus inputs control phytoplankton growth in eutrophic Lake Taihu, China[J]. Limnology and Oceanography, 2010, 55(1): 420-432. DOI:10.4319/lo.2010.55.1.0420 |
| [43] | Li J, Hansson L A, Persson K M. Nutrient control to prevent the occurrence of cyanobacterial blooms in a eutrophic lake in southern Sweden, used for drinking water supply[J]. Water, 2018, 10(7): 919. DOI:10.3390/w10070919 |
| [44] | Zhang M, Shi X L, Yang Z, et al. Long-term dynamics and drivers of phytoplankton biomass in eutrophic Lake Taihu[J]. Science of the Total Environment, 2018, 645: 876-886. DOI:10.1016/j.scitotenv.2018.07.220 |
| [45] | Paerl H W, Otten T G. Harmful cyanobacterial blooms:causes, consequences, and controls[J]. Microbial Ecology, 2013, 65(4): 995-1010. DOI:10.1007/s00248-012-0159-y |
| [46] |
许珍, 殷大聪, 陈进, 等. 温度和光强对4种常见水华藻叶绿素荧光特性的影响[J]. 长江科学院院报, 2017, 34(6): 39-44. Xu Z, Yin D C, Chen J, et al. Effects of temperature and illumination on chlorophyll fluorescence characteristics of four common bloom algae[J]. Journal of Yangtze River Scientific Research Institute, 2017, 34(6): 39-44. |
| [47] |
王敏, 袁绍春, 徐炜, 等. 嘉陵江重庆出口段小环藻水华影响因素分析[J]. 重庆交通大学学报(自然科学版), 2016, 35(6): 86-90, 94. Wang M, Yuan S C, Xu W, et al. Influence factor analysis on Cyclotella bloom of Jialing River Estuary in Chongqing[J]. Journal of Chongqing Jiaotong University (Natural Science), 2016, 35(6): 86-90, 94. |
| [48] | Yang W, Zheng Z M, Zheng C, et al. Temporal variations in a phytoplankton community in a subtropical reservoir:an interplay of extrinsic and intrinsic community effects[J]. Science of The Total Environment, 2018, 612: 720-727. DOI:10.1016/j.scitotenv.2017.08.044 |
| [49] | Guidi F, Pezzolesi L, Vanucci S. Microbial dynamics during harmful dinoflagellate Ostreopsis cf. ovata growth:bacterial succession and viral abundance pattern[J]. Microbiologyopen, 2018, 7(4): e00584. DOI:10.1002/mbo3.584 |
| [50] | Zhang H H, Chen S N, Huang T L, et al. Vertical distribution of bacterial community diversity and water quality during the reservoir thermal stratification[J]. International Journal of Environmental Research and Public Health, 2015, 12(6): 6933-6945. DOI:10.3390/ijerph120606933 |
| [51] |
商潘路, 陈胜男, 黄廷林, 等. 深水型水库热分层诱导水质及真菌种群结构垂向演替[J]. 环境科学, 2018, 39(3): 1141-1150. Shang P L, Chen S N, Huang T L, et al. Vertical distribution of fungal community composition and water quality during the deep reservoir thermal stratification[J]. Environmental Science, 2018, 39(3): 1141-1150. |
| [52] | Ji B, Qin H, Guo S D, et al. Bacterial communities of four adjacent fresh lakes at different trophic status[J]. Ecotoxicology and Environmental Safety, 2018, 157: 388-394. DOI:10.1016/j.ecoenv.2018.03.086 |
| [53] | Wei G S, Li J, Wang N X, et al. Spatial abundance and diversity of bacterioplankton in a typical stream-forming ecosystem, Huangqian Reservoir, China[J]. Journal of Microbiology and Biotechnology, 2014, 24(10): 1308-1318. DOI:10.4014/jmb.1403.03067 |
| [54] | Hahn M W, Pöckl M, Wu Q L. Low intraspecific diversity in a polynucleobacter subcluster population numerically dominating bacterioplankton of a freshwater pond[J]. Applied and Environmental Microbiology, 2005, 71(8): 4539-4547. DOI:10.1128/AEM.71.8.4539-4547.2005 |
| [55] | Bai S J, Huang L P, Su J Q, et al. Algicidal effects of a novel marine actinomycete on the toxic dinoflagellate Alexandrium tamarense[J]. Current Microbiology, 2011, 62(6): 1774-1781. DOI:10.1007/s00284-011-9927-z |
| [56] | Zhang H H, Huang T L, Chen S N, et al. Abundance and diversity of bacteria in oxygen minimum drinking water reservoir sediments studied by quantitative PCR and pyrosequencing[J]. Microbial Ecology, 2015, 69(3): 618-629. DOI:10.1007/s00248-014-0539-6 |
| [57] | Zeder M, Peter S, Shabarova T, et al. A small population of planktonic Flavobacteria with disproportionally high growth during the spring phytoplankton bloom in a prealpine lake[J]. Environmental Microbiology, 2009, 11(10): 2676-2686. DOI:10.1111/j.1462-2920.2009.01994.x |
| [58] |
谢丽娟, 余得昭, 曾诚, 等. 细菌群落组成对微囊藻水华分解过程的响应[J]. 湖泊科学, 2016, 28(1): 22-30. Xie L J, Yu D Z, Zeng C, et al. The response of bacterial community composition to the decomposition of Microcystis blooms[J]. Journal of Lake Science, 2016, 28(1): 22-30. |
| [59] | Bell T A S, Sen-Kilic E, Felföldi T, et al. Microbial community changes during a toxic cyanobacterial bloom in an alkaline Hungarian lake[J]. Antonie van Leeuwenhoek, 2018, 111(12): 2425-2440. DOI:10.1007/s10482-018-1132-7 |