 2020, Vol. 41
2020, Vol. 41



2. 青岛海洋科学与技术试点国家实验室, 青岛 266237;
3. 中国科学院大学地球与行星科学学院, 北京 100049;
4. 中国科学院海洋研究所, 青岛 266071
2. Pilot National Laboratory for Marine Science and Technology(Qingdao), Qingdao 266237, China;
3. College of Earth and Planetary Sciences, University of Chinese Academy of Sciences, Beijing 100049, China;
4. Institute of Oceanology, Chinese Academy of Sciences, Qingdao 266071, China
抗生素的滥用会带来较强毒副作用, 如直接伤害身体, 造成潜在基因污染, 使细菌产生交叉耐药性或共同耐药性等, 其危害已经引起了人们的广泛关注[1, 2].大量的抗生素及其代谢产物最终会进入海洋, 其对海洋生态系统的潜在威胁值得研究.海水中的抗生素浓度为ng·L-1至μg·L-1, 其中红霉素在水体中存在形式较为稳定, 检出频率高, 残留量较大[3, 4].
海洋生态系统中, 微藻是重要的生产者, 对环境变化很敏感.杨弯弯等[5]的研究发现硫氰酸红霉素对铜绿微囊藻(Microcystis aeruginosa)的生长有抑制作用, 且96h-EC50为48.2 g·L-1.刘滨扬[6]的研究发现红霉素可以显著抑制羊角月牙藻(Selenastrum bibraianum)的叶绿素生物合成.Perales-Vela等[7]证明了链霉素能够降低小球藻(Chlorella vulgaris)的生长速率和光合作用活性.Bashir等[8]的研究发现四环素通过调节蛋白质的合成来抑制微芒藻(Micractinium pusillum)的生长和光合作用.王洪斌等[9]的研究发现浓度低于200 μg·mL-1时, 链霉素对米氏凯伦藻(Karenia mikimotoi)有促进作用.抗生素会影响微藻的生长.目前研究多为抗生素对淡水微藻的影响, 抗生素对海洋微藻的影响研究相对较少.
在抗生素进入海洋的同时, 大量营养盐入海, 造成海水富营养化, 营养盐结构严重失衡.近年研究发现, 我国近海主要河口、海湾水体中N/P比值几乎都偏离Redfield比值[10].海水中的氮磷比将影响海洋浮游植物的生长速率和细胞内物质合成.那么红霉素与营养盐是否存在复合效应?目前这方面的研究相对匮乏.
本文以米氏凯伦藻(Karenia mikimotoi)、东海原甲藻(Prorocentrum donghaiense)和中肋骨条藻(Skeletonema costatum)为研究对象.在不同营养盐比例下, 以红霉素为目标抗生素, 以藻类生物量、光合色素含量、超氧化物歧化酶(SOD)活性、丙二醛(MDA)和可溶性蛋白质含量为评价指标, 分析了红霉素与营养盐对微藻生理的复合影响, 以期为评估抗生素对海洋生态的影响提供一定理论依据.
1 材料与方法 1.1 实验方法米氏凯伦藻、东海原甲藻和中肋骨条藻均由中国科学院海洋研究所藻种库提供.实验前取处于对数期的藻种过滤, 饥饿培养2 d后待用.将培养的藻种接种于500 mL的锥形瓶中, 每个锥形瓶的培养体积为400 mL, 接种的初始密度为5×102个·mL-1.本实验采用的光照强度为(2 500±100) lx, 光暗周期比为12 h:12 h, 培养温度(21±1)℃.培养液痕量元素及微生素按f/2配方配制, 营养盐浓度按照表 1浓度配制.其中米氏凯伦藻和东海原甲藻培养液中不添加硅营养盐.
|
|
表 1 培养液中营养盐浓度/μmol·L-1 Table 1 Nutrients concentrations in culture/μmol·L-1 |
红霉素购于北京索莱宝科技有限公司.设置红霉素的添加浓度系列为0、0.5、1、10、100和1 000 μg·L-1.设置营养比例添加系列分别为米氏凯伦藻和东海原甲藻为N:P=16:1、N:P=8:1和N:P=32:1;中肋骨条藻为N:P:Si=16:1:16、N:P:Si=8:1:16、N:P:Si=32:1:32和N:P:Si=16:1:8.
进行全因子实验, 对抗生素浓度-营养状态组合中的每一种设置3个平行.每天定时取样, 在显微镜下用计数框计数, 连续13 d测定微藻的生物量, 并绘制生长曲线.
取培养结束后的藻液5 mL过滤, 用80%丙酮萃取, 测定波长为663、645和470 nm下的吸光值, 计算其叶绿素a、叶绿素b和类胡萝卜素含量(mg·L-1).计算公式如下:

|
采用南京建成公司相应的试剂盒测定SOD (U·个-1)、MDA (nmol·个-1)和可溶性蛋白质含量(每个细胞中可溶性蛋白质的质量, 单位表示为g·个-1).
以红霉素浓度为0 μg·L-1和N:P=16:1(或N:P:Si=16:1:16)时的微藻的光合色素含量、SOD、MDA和可溶性蛋白质含量为对照组, 计算其他处理条件下(即实验组)相对对照组的微藻生理指标相对变化量(R), 计算公式为:
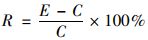
|
式中, R表示微藻的生理指标相对变化量; E表示实验组生理指标的平均值; C表示对照组生理指标的平均值.
根据各生理指标相对变化量, 绘制柱形图.R为正值表示该实验处理对微藻的生理指标有促进作用, R为负值表示有抑制作用.
1.2 数据处理分析采用SPSS 22.0进行差异性分析和相关性分析, 实验差异显著性水平为P < 0.05.
2 结果与分析 2.1 红霉素和营养盐对微藻的生长的复合影响当添加的红霉素浓度为1 000 μg·L-1时, 米氏凯伦藻的生长受到轻微抑制, 但并不显著(P>0.05).即在本实验范围内, 红霉素对米氏凯伦藻的生长并无明显影响[图 1(a)].营养盐限制时, 米氏凯伦藻的生长均受到了抑制.同一红霉素浓度下, 不同营养盐条件处理下的细胞密度:N:P=16:1>N:P=32:1>N:P=8:1;且同一红霉素浓度下, 本实验范围内缺氮对米氏凯伦藻影响最明显(P<0.05);缺磷虽会抑制米氏凯伦藻的生长, 但并不显著(P>0.05).

|
(a)和(b)中A表示N:P=16:1, B表示N:P=8:1, C表示N:P=32:1; (c)中A表示N:P:Si=16:1:16, B表示N:P:Si=8:1:16, C表示N:P:Si=32:1:32, D表示N:P:Si=16:1:8; 1~6表示红霉素的浓度分别为0、0.5、1、10、100和1 000 μg·L-1, 下同 图 1 红霉素和营养盐对微藻的生长的复合影响 Fig. 1 Combined effects of erythromycin and nutrients on the growth of microalgae |
虽然东海原甲藻和米氏凯伦藻都是甲藻, 但红霉素对东海原甲藻的生长的影响更明显(P<0.05).如图 1(b)所示, 当红霉素浓度为100 μg·L-1和1 000 μg·L-1时, 东海原甲藻的细胞密度显著减少(P<0.05).在高浓度红霉素的胁迫下, 东海原甲藻的生长受到了显著抑制(P<0.05).当红霉素浓度较低时, 红霉素对东海原甲藻的生长无明显影响(P>0.05).氮限制(N:P=8:1)和磷限制(N:P=32:1)时, 东海原甲藻的细胞密度都比对照组(N:P=16:1)低.且磷限制时, 东海原甲藻的细胞密度最低.磷限制(N:P=32:1)对东海原甲藻的生长的影响最为显著(P<0.05).红霉素浓度与营养盐限制对东海原甲藻的生物量的影响具有协同作用.
当红霉素浓度为100 μg·L-1时, 中肋骨条藻的细胞密度显著减少(P<0.05);当红霉素浓度为1 000 μg·L-1时, 其细胞密度的减少极显著[P<0.01, 图 1(c)].在高浓度红霉素的胁迫下, 中肋骨条藻的生长受到了显著抑制(P<0.05).在这3种微藻中, 同一营养盐条件下红霉素对中肋骨条藻的胁迫最严重.本实验中, 氮限制和磷限制时, 细胞密度较低.本实验范围内, N和P的限制对中肋骨条藻的影响更明显(P<0.05).
2.2 红霉素和营养盐对微藻的光合色素含量的复合影响当N:P=16:1时, 米氏凯伦藻细胞中叶绿素a含量在红霉素浓度为0.5 μg·L-1时最高, 为1.17×10-8 mg·个-1, 增高了23.7%, 之后随着处理浓度升高藻细胞中叶绿素a含量逐渐降低[图 2(a)].在氮限制组(N:P=8:1)中, 除B2外红霉素各处理浓度中的米氏凯伦藻叶绿素a含量均显著低于0 μg·L-1时的叶绿素a含量(P<0.05).在磷限制组(N:P=32:1)中, 除红霉素浓度为10 μg·L-1和100 μg·L-1时叶绿素a含量较0 μg·L-1时略有升高(分别升高了6.3%和10.5%), 在红霉素其他处理浓度中, 藻细胞中叶绿素a含量均显著降低(P<0.05).因此, 高浓度红霉素会抑制米氏凯伦藻叶绿素a的合成.当N:P=16:1时, 米氏凯伦藻细胞中叶绿素a含量在红霉素浓度为0.5 μg·L-1时最高; 但氮限制时, 米氏凯伦藻细胞中叶绿素a含量在红霉素浓度为0 μg·L-1时最高, 即氮限制时米氏凯伦藻的叶绿素a合成会对红霉素的浓度更敏感.不同营养条件下, 米氏凯伦藻类胡萝卜素均呈现先升后降的趋势, 低浓度红霉素对米氏凯伦藻的类胡萝卜素有促进效果, 高浓度红霉素会抑制类胡萝卜素的合成.

|
(a)和(b)中A表示N:P=16:1, B表示N:P=8:1, C表示N:P=32:1; (c)中A表示N:P:Si=16:1:16, B表示N:P:Si=8:1:16, C表示N:P:Si=32:1:32, D表示N:P:Si=16:1:8; 1~6表示红霉素的浓度分别为0、0.5、1、10、100和1 000 μg·L-1 图 2 3种微藻不同实验组的光合色素相对变化量 Fig. 2 Relative variation of photosynthetic pigments in different experimental groups of three microalgae |
当N:P=16:1时, 不同于米氏凯伦藻, 东海原甲藻藻细胞中叶绿素a含量在红霉素浓度为0.5 μg·L-1时最高, 为3.80×10-9 mg·个-1, 之后随着处理浓度升高藻细胞中叶绿素a含量呈现先降后升的趋势.当红霉素浓度为10 μg·L-1时, 藻细胞中叶绿素a含量最低, 为2.23×10-9 mg·个-1 [图 2(b)].在氮限制组(N:P=8:1)中, 叶绿素a含量也显现出先降后升的趋势.在磷限制组(N:P=32:1)中, 叶绿素a含量随红霉素浓度的升高出现先升后降的现象.此外, 当N:P=16:1时, 随着红霉素浓度升高, 东海原甲藻类胡萝卜素含量逐渐增多.氮限制和磷限制时东海原甲藻细胞内类胡萝卜素含量先升高后降低.氮限制时, 红霉素浓度为100 μg·L-1时的类胡萝卜素含量最高, 为1.20×10-9 mg·个-1.磷限制时, 红霉素浓度为10 μg·L-1时的类胡萝卜素含量最高, 为2.02×10-9 mg·个-1.
红霉素会抑制中肋骨条藻的叶绿素a的合成.由图 2(c)可知, N:P=16:1时, 中肋骨条藻的叶绿素a含量随红霉素浓度升高逐渐降低.当N:P:Si=8:1:16和N:P:Si=32:1:32时, 除红霉素浓度为1 000 μg·L-1外, 其他红霉素处理浓度下的中肋骨条藻叶绿素a含量均随红霉素浓度升高逐渐降低.氮限制时, 叶绿素a含量明显低于其他营养条件(P<0.05).缺氮会显著抑制中肋骨条藻叶绿素a的合成.当N:P:Si=16:1:16时, 中肋骨条藻类胡萝卜素无明显变化(P>0.05).其他营养比例下, 中肋骨条藻类胡萝卜素含量随红霉素浓度升高逐渐降低, 即红霉素会抑制类胡萝卜素的合成.
2.3 红霉素和营养盐对微藻的可溶性蛋白质含量的复合影响如图 3(a)所示, 当N:P=16:1时, 红霉素各处理浓度中的米氏凯伦藻细胞内的可溶性蛋白质含量均高于对照组(0 μg·L-1), 为先增高后下降的趋势; 当红霉素浓度为1 μg·L-1时米氏凯伦藻细胞内的可溶性蛋白质含量最多, 为4.04×10-10 g·个-1, 较对照组增多了31.5%.当N:P=32:1时, 米氏凯伦藻细胞内的可溶性蛋白质含量随红霉素浓度的增高先增高后下降, 在红霉素浓度为1 μg·L-1时可溶性蛋白质含量最多.当N:P=8:1时, 随着红霉素处理浓度的增高, 米氏凯伦藻细胞内的可溶性蛋白质含量逐渐降低.当N:P=16:1和N:P=32:1时, 红霉素对米氏凯伦藻细胞内可溶性蛋白质存在低促高抑的现象.当N:P=8:1时, 红霉素会抑制米氏凯伦藻蛋白质的合成.

|
(a)和(b)中A表示N:P=16:1, B表示N:P=8:1, C表示N:P=32:1; (c)中A表示N:P:Si=16:1:16, B表示N:P:Si=8:1:16, C表示N:P:Si=32:1:32, D表示N:P:Si=16:1:8; 1~6表示红霉素的浓度分别为0、0.5、1、10、100和1 000 μg·L-1 图 3 3种微藻不同实验组的蛋白质相对变化量 Fig. 3 Relative variation of soluble proteins in different experimental groups of three microalgae |
而对于东海原甲藻来说, 低浓度的红霉素对东海原甲藻细胞内的蛋白质含量无明显影响(P>0.05).当红霉素处理浓度为1 000 μg·L-1时, 东海原甲藻细胞内的蛋白质含量显著增多(P<0.05)[图 3(b)].
中肋骨条藻对红霉素的胁迫最敏感.如图 3(c)所示, 当N:P:Si=16:1:16和N:P:Si=32:1:32时, 中肋骨条藻的蛋白质含量随红霉素浓度升高逐渐降低; 当红霉素浓度为1 000 μg·L-1时, 其蛋白质含量较0 μg·L-1时分别降低了33.5%和26.1%.当N:P:Si=16:1:8时, 细胞内蛋白质含量先增后减, 当红霉素浓度为10μg·L-1时, 细胞内蛋白质含量最高, 为4.15×10-11 g·个-1, 增多了20.3%.氮限制时, 蛋白质含量明显低于其他营养条件(P<0.05).此时氮为主要限制因子, 红霉素的影响不明显(P>0.05).综上所述, 红霉素会抑制中肋骨条藻可溶性蛋白质的合成.
2.4 红霉素和营养盐对微藻的SOD和MDA的复合影响当N:P=16:1时, 米氏凯伦藻细胞内SOD活性随红霉素浓度增高逐渐升高, 当红霉素浓度为1 000 μg·L-1时, SOD活性增高了40.7%[图 4(a1)]; 红霉素处理后细胞内的MDA含量也要高于对照组(0 μg·L-1), 且MDA含量逐渐增多[图 4(a2)].当N:P=8:1时, 米氏凯伦藻细胞内的SOD活性随红霉素浓度增高而逐渐降低, MDA则逐渐升高, 两者成显著负相关(P<0.05, r=-0.863).当N:P=32:1时, 细胞内的SOD呈现先降低后升高的趋势, MDA含量在红霉素处理浓度为1 000 μg·L-1时显著增高(P<0.01), 增高了138%.当红霉素浓度为0 μg·L-1时, N和P限制时细胞内的MDA含量显著增多(P<0.05), 分别增多了276%和245%.氮限制MDA含量更多, 氧化损伤更严重.红霉素浓度升高, 细胞内MDA增多, 细胞氧化损伤加重.氮限制时, 红霉素对米氏凯伦藻的胁迫会更严重.

|
(a1)、(a2)、(b1)和(b2)中A表示N:P=16:1, B表示N:P=8:1, C表示N:P=32:1; (c1)和(c2)中A表示N:P:Si=16:1:16, B表示N:P:Si=8:1:16, C表示N:P:Si=32:1:32, D表示N:P:Si=16:1:8; 1~6表示红霉素的浓度分别为0、0.5、1、10、100和1 000 μg·L-1 图 4 3种微藻不同实验组的SOD和MDA相对变化量 Fig. 4 Relative variation of SOD and MDA in different experimental groups of three microalgae |
当N:P=16:1和氮限制(N:P=8:1)时, 东海原甲藻细胞内的SOD和MDA均成显著正相关(P<0.01, r=0.838和P<0.01, r=0.802), 都随着红霉素处理浓度的增高而增高; 受到高浓度红霉素(≥100 μg·L-1)的胁迫后, 细胞内的SOD和MDA显著增高(P<0.05)[图 4(b)].磷限制(N:P=32:1)时, 当红霉素浓度≤100 μg·L-1时, 细胞内的SOD活性随红霉素浓度增高而逐渐降低.磷限制(N:P=32:1)时, 细胞内的MDA含量最多, 细胞氧化损伤最严重.当氮磷比为16:1时, 东海原甲藻细胞的抗氧化能力随着红霉素浓度升高而逐渐增高; 当磷限制时, 其抗氧化能力随红霉素浓度升高而逐渐降低.磷限制时, 红霉素对东海原甲藻的胁迫最严重.
当N:P:Si=16:1:16时, 中肋骨条藻的SOD活性随红霉素浓度升高逐渐增大; 其他营养状态时SOD活性随红霉素浓度升高逐渐减小[图 4(c1)].磷限制和硅限制时, SOD活性明显低于其他营养条件(P<0.05).当N:P:Si=16:1:16和N:P:Si=16:1:8时, 中肋骨条藻的MDA含量随红霉素浓度升高逐渐增多.当N:P:Si=16:1:16时, 中肋骨条藻的SOD和MDA成显著正相关(P<0.01, r=0.735).当N:P:Si=16:1:16时, 中肋骨条藻细胞的抗氧化能力随着红霉素浓度升高而逐渐增高, 在营养盐限制时其抗氧化能力会逐渐降低.营养盐限制使得中肋骨条藻对红霉素的耐受性降低.
3 讨论营养盐比例对于微藻的影响的研究一直是热点.马龙等[11]的研究认为米氏凯伦藻生长的最适氮磷比值为24.4, 缺氮和缺磷都不利于米氏凯伦藻的生长.沈盎绿等[12]的研究认为磷更能影响东海原甲藻的增殖.刘东艳等[13]的研究认为中肋骨条藻的最适氮磷比为16:1, 氮磷比大于16时的生长速度要优于氮磷比小于16时的生长速度.本实验中, 米氏凯伦藻、东海原甲藻和中肋骨条藻的最适营养盐比例为N:P=16:1.缺氮对米氏凯伦藻影响最明显; 缺磷对东海原甲藻影响最明显; 对中肋骨条藻来说, N和P的限制对其影响较硅更明显, 缺氮对中肋骨条藻影响最明显.本次实验结果与上述结果相同.
目前研究发现, 红霉素抑制了淡水微藻莱茵衣藻(Chlamydomonas reinhardtii)[14]和铜绿微囊藻[3]种群的生长, 对海洋硅藻三角褐指藻(Phaeodactylum tricornutum)[14]的生长有抑制作用.本研究发现, 实验范围内, 红霉素对米氏凯伦藻的生长并无明显影响, 高浓度红霉素(≥100 μg·L-1)会显著抑制东海原甲藻和中肋骨条藻的生长.
光合色素是反映逆境胁迫状况的重要指标之一.刘滨扬[6]的研究发现红霉素可以显著抑制羊角月牙藻的叶绿素a生物合成.Bishop等[15, 16]的研究发现, 氯霉素可以显著抑制纤细裸藻(Euglena gracilis)的叶绿素生物合成.张文慧等[17]的研究也发现在土霉素胁迫作用下, 纤细裸藻叶绿素a合成受到抑制.红霉素、氯霉素和土霉素同属于蛋白质合成抑制类抗生素, 应具有相似的效应.本实验中, 红霉素浓度低于0.5 μg·L-1会促进米氏凯伦藻和东海原甲藻的叶绿素a的合成, 此时低浓度红霉素对米氏凯伦藻和东海原甲藻的生长有抑菌作用, 有利于米氏凯伦藻和东海原甲藻的生长.红霉素是蛋白质合成抑制类抗生素, 其作用机制主要是与核糖核蛋白体的50S亚单位相结合, 抑制肽酰基转移酶, 抑制细菌蛋白质的合成[18].而叶绿体基因与原核生物基因相似, 因此红霉素可能对叶绿体基因的表达有抑制作用, 对叶绿体蛋白质的合成有抑制作用, 导致叶绿素含量降低[19~21].所以红霉素浓度高于0.5 μg·L-1时会抑制米氏凯伦藻和东海原甲藻中叶绿素a的合成; 红霉素会抑制中肋骨条藻叶绿素a的合成.在叶绿素a的合成方面, 中肋骨条藻对红霉素的敏感性最高.低浓度红霉素对不同藻影响的差异机制还有待进一步研究.但本实验中, 当红霉素浓度为1 000 μg·L-1时的东海原甲藻细胞中叶绿素a含量较红霉素浓度为100 μg·L-1时的含量高, 这可能是由于红霉素1 000 μg·L-1时东海原甲藻细胞密度较低, 营养盐相对充足, 高营养盐组的东海原甲藻叶绿素a浓度要高于营养盐缺乏组的浓度[22].
类胡萝卜素除了在光合作用中有一定作用外, 还可以淬灭活性氧, 防止膜脂过氧化, 保护叶绿素和光合机能[23].因此非营养盐限制时, 米氏凯伦藻、东海原甲藻和中肋骨条藻受红霉素胁迫, 类胡萝卜素含量增多, 细胞抗氧化能力增强, 有利于保护细胞光合机能.但营养盐限制时, 米氏凯伦藻和东海原甲藻受红霉素胁迫细胞中类胡萝卜素含量先增多后减少, 高浓度红霉素胁迫下, 细胞不能再维持产生和清除自由基的动态平衡, 此时的红霉素浓度已经超过调节限度, 细胞氧化损伤严重, 光合作用受到抑制; 营养盐限制时, 中肋骨条藻受红霉素胁迫细胞中类胡萝卜素含量逐渐减少, 即营养盐限制时, 0.5 μg·L-1红霉素就已超过中肋骨条藻的调节限度, 膜脂过氧化, 光合作用受到抑制.营养盐限制使得微藻在红霉素对微藻光合色素合成方面的胁迫更加敏感.
可溶性蛋白质与细胞内多种代谢过程紧密相关, 在受到外界胁迫时, 可溶性蛋白质含量增多, 以维持细胞较低的渗透势, 提高自身抗性; 当超过其调节限度时, 蛋白质含量降低, 蛋白质合成受损[24].本实验中, 非氮限制时, 低浓度红霉素(≤1 μg·L-1)胁迫下米氏凯伦藻可溶性蛋白质含量上升, 以提高自身抗性; 高浓度红霉素超过其调节限度, 蛋白质含量降低, 蛋白质合成受损.氮限制时, 红霉素会抑制米氏凯伦藻蛋白质的合成, 即氮限制时, 低浓度红霉素已超过米氏凯伦藻的调节限度, 蛋白质合成受损.而对于东海原甲藻来说, 当红霉素浓度为1 000 μg·L-1时, 细胞内的蛋白质含量较其他处理组显著增多, 高浓度红霉素有利于东海原甲藻可溶性蛋白质的合成, 提高自身抗性.红霉素会抑制中肋骨条藻的蛋白质合成, 即0.5 μg·L-1红霉素就已超过中肋骨条藻的调节限度, 因此蛋白质合成受损.中肋骨条藻对红霉素的敏感性最高.
MDA是脂质过氧化的产物, 其含量可以解释胁迫下的氧化和脂质损伤.SOD是氧化应激的重要生物指标.受到红霉素胁迫时, 微藻产生大量的MDA, 细胞氧化损伤程度随红霉素浓度的升高而增大, 抗氧化能力也随之升高.当超过其调节限度时, 抗氧化能力降低.Wu等[25]的研究发现添加土霉素可加重等鞭金藻(Isochrysis galbana)细胞膜过氧化.Liu等[26]的研究也发现具有抑制蛋白质功能的氟苯尼考能加重中肋骨条藻的氧化损伤.此次实验也发现, 红霉素可以加重米氏凯伦藻、东海原甲藻和中肋骨条藻的氧化损伤.同时营养盐限制会使微藻氧化损伤加重, 抗氧化能力降低.
综上所述, 红霉素和营养盐结构会影响米氏凯伦藻、东海原甲藻和中肋骨条藻的生长.不同的营养结构下, 同种微藻对红霉素的响应是存在差异的.因此, 在评估海水中的红霉素对微藻生理的影响时, 需要考虑海域的营养盐结构.受人类活动、降水变化等的影响, 海域营养盐结构失衡, 存在时空变化[27].营养盐结构不同的海域里, 同种微藻对红霉素的响应是不同的.同一海域, 不同季节时, 同种微藻对红霉素的响应也是有差异的.不同微藻对红霉素的敏感性是不同的.在这3种微藻中, 中肋骨条藻最敏感, 其次是东海原甲藻, 米氏凯伦藻最不敏感.在红霉素的胁迫下, 这3种微藻中米氏凯伦藻可能更具竞争力.在种间竞争中, 米氏凯伦藻在面临红霉素胁迫时可能更具优势.
4 结论本实验范围内, 红霉素的浓度对米氏凯伦藻的生长影响不显著, 高于100 μg·L-1的红霉素会显著抑制东海原甲藻和中肋骨条藻的生长.红霉素可以加重米氏凯伦藻、东海原甲藻和中肋骨条藻的氧化损伤.不同微藻对红霉素的敏感性是不同的, 其中中肋骨条藻最敏感, 而米氏凯伦藻最不敏感.不同的营养结构下, 同种微藻对红霉素的响应是存在差异的.营养盐限制会降低米氏凯伦藻、东海原甲藻、中肋骨条藻对红霉素的耐受性.红霉素和营养盐会对微藻的生长产生复合影响, 会对海洋生态产生一定影响.目前, 海水营养盐结构严重失衡, 虽然海水中的抗生素浓度为ng·L-1至μg·L-1, 可能没有实验中的红霉素浓度那么高, 但若不加以控制, 可能会对海水中的微藻造成严重影响, 进而破坏海洋生态环境的生态平衡.
| [1] | Gothwal R, Shashidhar T. Antibiotic pollution in the environment:a review[J]. Clean-Soil, Air, Water, 2015, 43(4): 479-489. DOI:10.1002/clen.201300989 |
| [2] |
刘昌孝. 当代抗生素发展的挑战与思考[J]. 中国抗生素杂志, 2017, 42(1): 1-12. Liu C X. Challenges and thinking of current antibiotic development[J]. Chinese Journal of Antibiotics, 2017, 42(1): 1-12. DOI:10.3969/j.issn.1001-8689.2017.01.002 |
| [3] | Zhang R J, Tang J H, Li J, et al. Antibiotics in the offshore waters of the Bohai Sea and the Yellow Sea in China:occurrence, distribution and ecological risks[J]. Environmental Pollution, 2013, 174: 71-77. DOI:10.1016/j.envpol.2012.11.008 |
| [4] | Zheng Q, Zhang R J, Wang Y H, et al. Occurrence and distribution of antibiotics in the Beibu Gulf, China:impacts of river discharge and aquaculture activities[J]. Marine Environmental Research, 2012, 78: 26-33. DOI:10.1016/j.marenvres.2012.03.007 |
| [5] |
杨弯弯, 武氏秋贤, 吴亦潇, 等. 恩诺沙星和硫氰酸红霉素对铜绿微囊藻的毒性研究[J]. 中国环境科学, 2013, 33(10): 1829-1834. Yang W W, Hien V T T, Wu Y X, et al. Toxicity of enrofloxacin and erythromycin thiocyanate on Microcystis aeruginosa[J]. China Environmental Science, 2013, 33(10): 1829-1834. |
| [6] |
刘滨扬.红霉素、环丙沙星和磺胺甲唑对羊角月牙藻的毒性效应及其作用机理[D].广州: 暨南大学, 2011. Liu B Y. Toxic effects and its mechanism of erythromycin, ciprofloxacin and sulfamethoxazole to Selenastrum capricornutum[D]. Guangzhou: Ji'nan University, 2011. http://cdmd.cnki.com.cn/Article/CDMD-10559-1011128731.htm |
| [7] | Perales-Vela H V, García R V, Gómez- Juárez E A, et al. Streptomycin affects the growth and photochemical activity of the alga Chlorella vulgaris[J]. Ecotoxicology and Environmental Safety, 2016, 132: 311-317. DOI:10.1016/j.ecoenv.2016.06.019 |
| [8] | Bashir K M I, Cho M G. The effect of kanamycin and tetracycline on growth and photosynthetic activity of two chlorophyte algae[J]. BioMed Research International, 2016, 2016: 5656304. |
| [9] |
王洪斌, 成明, 杨艳艳, 等. 绿色巴夫藻和米氏凯伦藻对3种常见抗生素胁迫的响应研究[J]. 水产科学, 2012, 31(6): 329-332. Wang H B, Cheng M, Yang Y Y, et al. The responses of algae Pavlova viridis and Karenia mikimotoi to three conventional antibiotics stress[J]. Fisheries Science, 2012, 31(6): 329-332. DOI:10.3969/j.issn.1003-1111.2012.06.004 |
| [10] |
祝雅轩, 裴绍峰, 张海波, 等. 莱州湾营养盐和富营养化特征与研究进展[J]. 海洋地质前沿, 2019, 35(4): 1-9. Zhu Y X, Pei S F, Zhang H B, et al. Characteristics and research progress of nutrients and eutrophication in Laizhou Bay, China[J]. Marine Geology Frontiers, 2019, 35(4): 1-9. |
| [11] |
马龙, 王培刚, 朱葆华, 等. 米氏凯伦藻与亚心形扁藻在不同营养盐水平下的种间竞争[J]. 海洋环境科学, 2013, 32(2): 221-226. Ma L, Wang P G, Zhu B H, et al. Inter-specific competition of Karenia mikimotoi and Platymonas subcordiformis under different level of nutrients[J]. Marine Environmental Science, 2013, 32(2): 221-226. |
| [12] |
沈盎绿, 李道季. 不同营养盐水平对东海原甲藻和米氏凯伦藻生长的影响[J]. 海洋渔业, 2016, 38(4): 415-423. Shen A L, Li D J. Effects of different nutrients levels on the growth of Prorocentrum donghaiense and Karenia mikimotoi[J]. Marine Fisheries, 2016, 38(4): 415-423. DOI:10.3969/j.issn.1004-2490.2016.04.009 |
| [13] |
刘东艳, 孙军, 陈宗涛, 等. 不同氮磷比对中肋骨条藻生长特性的影响[J]. 海洋湖沼通报, 2002(2): 39-44. Liu D Y, Sun J, Chen Z T, et al. Effect of N/P ratio on the growth of a red tide diatom Skeletonema costatum[J]. Transactions of Oceanology and Limnology, 2002(2): 39-44. DOI:10.3969/j.issn.1003-6482.2002.02.006 |
| [14] | Sendra M, Moreno-Garrido I, Blasco J, et al. Effect of erythromycin and modulating effect of CeO2 NPs on the toxicity exerted by the antibiotic on the microalgae Chlamydomonas reinhardtii and Phaeodactylum tricornutum[J]. Environmental Pollution, 2018, 242: 357-366. DOI:10.1016/j.envpol.2018.07.009 |
| [15] | Bishop D G, Bain J M, Smillie R M. The effect of antibiotics on the ultrastructure and photochemical activity of a developing chloroplast[J]. Journal of Experimental Botany, 1973, 24(2): 361-362. DOI:10.1093/jxb/24.2.361 |
| [16] | Bishop D G. The effect of polyene antibiotics on photosynthetic electron transfer[J]. Journal of Experimental Botany, 1974, 25(86): 491-502. |
| [17] |
张文慧, 窦勇, 高金伟, 等. 纤细裸藻对5种抗生素的胁迫响应[J]. 水生态学杂志, 2017, 38(6): 48-56. Zhang W H, Dou Y, Gao J W, et al. Response of Euglena gracilis to five kinds of antibiotic stress[J]. Journal of Hydroecology, 2017, 38(6): 48-56. |
| [18] | 刘巨源, 张晓丹. 药理学[M]. 郑州: 郑州大学出版社, 2007. |
| [19] | Sandmann G, Böger P. Inhibition of photosynthetic electron transport by amphotericin B[J]. Physiologia Plantarum, 1981, 51(3): 326-328. DOI:10.1111/j.1399-3054.1981.tb04486.x |
| [20] | Koussevitzky S, Nott A, Mockler T C, et al. Signals from chloroplasts converge to regulate nuclear gene expression[J]. Science, 2007, 316(5825): 715-719. DOI:10.1126/science. 1140516 |
| [21] | Vannini C, Domingo G, Marsoni M, et al. Effects of a complex mixture of therapeutic drugs on unicellular algae Pseudokirchneriella subcapitata[J]. Aquatic Toxicology, 2010, 101(2): 459-465. |
| [22] |
孙开明, 李瑞香, 李艳, 等. 东海原甲藻光合系统Ⅱ荧光参数在种群生长中的变化[J]. 海洋环境科学, 2016, 35(2): 226-230. Sun K M, Li R X, Li Y, et al. Changes of fluorescence parameters of photosynthetic system Ⅱ in Prorocentrum donghaisense Lu during population growth[J]. Marine Environmental Science, 2016, 35(2): 226-230. |
| [23] | Thurnham D. Anti-oxidant vitamins and cancer prevention[J]. Journal of Micronutrient Analysis, 1990, 7(3-4): 279-299. |
| [24] |
万禁禁.淡水微藻对几种抗生素胁迫的响应研究[D].泉州: 华侨大学, 2014. Wan J J. Study the response of freshwater microalgae to several antibiotics stress[D]. Quanzhou: Huaqiao University, 2014. http://www.wanfangdata.com.cn/details/detail.do?_type=degree&id=D536444 |
| [25] | Wu C L, He C Q. Interaction effects of oxytetracycline and copper at different ratios on marine microalgae Isochrysis galbana[J]. Chemosphere, 2019, 225: 775-784. DOI:10.1016/j.chemosphere.2019.03.067 |
| [26] | Liu W H, Ming Y, Huang Z W, et al. Impacts of florfenicol on marine diatom Skeletonema costatum through photosynthesis inhibition and oxidative damages[J]. Plant Physiology and Biochemistry, 2012, 60: 165-170. DOI:10.1016/j.plaphy.2012.08.009 |
| [27] | Wei Q S, Yao Q Z, Wang B D, et al. Long-term variation of nutrients in the southern Yellow Sea[J]. Continental Shelf Research, 2015, 111: 184-196. DOI:10.1016/j.csr.2015.08.003 |