 2020, Vol. 41
2020, Vol. 41


2. 东华大学环境科学与工程学院, 上海 201620
2. College of Environmental Science and Engineering, Donghua University, Shanghai 201620, China
随着城市化进程加快, 市政污水处理厂建设规模不断扩大, 产生了越来越多剩余污泥需要合理处理与处置问题[1].厌氧消化处理剩余污泥, 不仅能有效缓解污泥填埋场地限制问题, 而且通过回收厌氧发酵产生的CH4可以实现对污泥蕴含能量的利用.污水处理厂剩余污泥具有高含水率、高热值和有机质丰富等特点, 理论上可以作为厌氧代谢过程理想底物[2].但实际应用中, 受制于各种因素影响, 剩余污泥高效厌氧发酵往往难以实现[3].如何提高污泥厌氧消化效率, 对于解决污泥问题意义重大.
以CO2和H2为代谢底物的嗜氢型产甲烷是厌氧过程的重要组成, 该代谢和产氢产乙酸过程密切耦合.发酵细菌和产甲烷菌形成的“互营”产CH4过程关键环节在于种间电子传递, 种间H2传递被视为该过程主要电子传递方式[4].最新研究表明[5], 互营氧化产甲烷过程也存在种间直接电子传递(direct interspecies electron transfer, DIET), 相比于种间H2传递, DIET对底物和中间代谢产物的利用更加稳定和快速.通过DIET途径, 产甲烷菌能够利用H+、e-和CO2直接产CH4, 从而克服种间H2传递过程存在的缺点.向厌氧消化系统中投加碳基和金属纳米材料可以强化发酵细菌和产甲烷菌之间的DIET[6], 颗粒活性炭(granular activated carbon, GAC)是研究最多的碳基材料, 纳米金属氧化物材料主要为纳米磁铁矿和纳米氧化锌等.目前, 大多数研究集中在单一材料对产CH4过程的影响, 多种材料的组合作用研究相对较少.
本实验即是针对剩余污泥厌氧消化效率普遍较低问题, 通过投加GAC和MnO2的方式, 以期利用DIET途径改善剩余污泥厌氧消化系统效能, 研究投加物对污泥厌氧消化系统挥发性脂肪酸(volatile fatty acids, VFAs)、产CH4效果和微生物活性(包括辅酶F420、细胞色素C、电子传递活性)等的影响, 并分析系统微生物群落结构变化特征, 以期为使用碳基和金属氧化物材料强化剩余污泥厌氧消化过程的实际应用提供理论依据和数据支撑.
1 材料与方法 1.1 接种污泥与剩余污泥接种污泥厌氧颗粒污泥取自实验室强化循环厌氧反应器(strengthen circulation anaerobic reactor, SCAR), 污泥粒径范围为0.6~1.4 mm;剩余污泥(脱水后泥饼, 含水率约为60%)取自上海市松江区污水处理厂.
将剩余污泥泥饼于60℃低温干燥24 h后, 用粉碎机粉碎过筛, 制得粒径<1 mm的污泥粉末, 根据需求配制所需浓度泥浆.接种污泥和剩余污泥基本性质如表 1所示.
|
|
表 1 接种污泥和剩余污泥基本性质 Table 1 Characteristics of seed sludge and wasted activated sludge |
1.2 材料制备
本实验棒状纳米MnO2采用Wang等[7]的方法制备:称取0.61 g KMnO4溶于78.7 mL的去离子水中, 再加入1.3 mL的HCl(37%);充分溶解后, 将混合溶液转移至100 mL反应釜内, 在140℃的条件下热处理12 h;待混合液冷却至室温后, 用定量滤纸进行过滤, 并用去离子水和无水乙醇各清洗3次, 之后在60℃的鼓风干燥箱烘干24 h;使用马弗炉对干燥样品进行煅烧处理, 煅烧温度自室温以1℃·min-1的速率升高至500℃, 并在该温度下保持3 h, 得到的固体颗粒即为α晶型的棒状纳米MnO2.
本实验GAC材料购买自国药集团, GAC的详细特征见表 2.
|
|
表 2 GAC的特征 Table 2 Characteristics of GAC |
1.3 实验装置与运行
本实验反应器为有效容积250 mL的玻璃瓶, 顶端设有取样口和出气口各1个, 出气口连接气体收集袋.接种污泥和剩余污泥按照体积比1 :4的比例接种到反应器内, 实验设置4组, 每组3个平行样, 其中包括空白组(R0)和实验组[GAC(R1)、MnO2(R2)和GAC/MnO2(R3)];各反应器材料具体添加量见表 3.用0.1 mol·L-1的NaOH调节反应器内pH为6.8~7.2, 并用N2吹脱30 min后密封置于转速130 r·min-1、温度为(37±1)℃的恒温振荡器中运行.每隔2 d取样测定, 共运行28 d.
|
|
表 3 各反应器内材料添加情况 Table 3 Material addition to each reactor |
1.4 分析项目及方法 1.4.1 基本指标
MLSS和MLVSS的测定采用标准方法[8];用排水法测定产气总量和CH4产量, 测CH4体积之前, 以酚酞为指示剂, 用2 mol·L-1的NaOH吸收酸性气体(如CO2)[9];CH4和CO2采用气相色谱(Techcomp GC 7900)法测定;挥发性脂肪酸(VFAs)采用气相色谱(GC-2010 Pro)法测定, 测定前样品用0.45 μm滤膜过滤并用甲酸(3%)酸化[10];多糖采用蒽酮-硫酸法测定[11], 蛋白质采用BCA法测定[12];细胞色素C(Cyt C)和辅酶F420采用荧光分光光度法测量[13];锰离子含量采用原子发射光谱法测量.
1.4.2 电子传递活性(INT-ETS)采用碘硝基氯化四氮唑蓝(INT)法[13]测定微生物电子传递活性(INT-ETS):取0.5 mL混合污泥于10 mL离心管中, 加入0.1 mL 0.2% INT溶液, 在黑暗条件下于摇床反应30 min(150 r·min-1, 37℃), 结束后加入1 mL 37%甲醛溶液停止该反应, 8 000 g转速下离心5 min, 弃去上清液, 加5 mL甲醇在摇床中反应10 min(150 r·min-1, 37℃), 在8 000 g转速下离心5 min, 最后取上清液于485 nm波长处测得其吸光度.沉淀物烘干(105℃)至恒重.
计算公式:
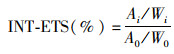
|
式中, Ai和A0分别代表实验组和空白组在485 nm处的吸光度, Wi和W0分别代表实验组和空白组的质量.
1.5 DNA提取与高通量测序本实验采用高通量测序技术对污泥样品进行微生物群落分析.污泥样品用磷酸缓冲溶液溶液清洗2次, 然后在8 000 g转速下离心15 min(4℃), 使用DNA萃取试剂盒(Bioteke Corporation, Beijing, China)提取DNA.分别采用(Arch340F/Arch806R)和(338F/806R)引物对古菌和细菌的16S rRNA进行PCR扩增.经过纯化和量化处理后, 委托上海美吉生物医药科技有限公司进行测序分析.
2 结果与讨论 2.1 投加GAC、MnO2和GAC/MnO2对产气量的影响GAC和MnO2对厌氧系统产CH4效果影响见图 1, 其分别反映R0、R1、R2和R3的CH4和CO2累计产生量以及产CH4速率随时间变化情况.

|
图 1 剩余污泥厌氧消化过程累计CH4、CO2气体产生量和产CH4速率 Fig. 1 Changes in cumulative CH4 production, cumulative CO2 production, and CH4 yield rate during anaerobic digestion of waste activated sludge |
由图 1(a)可见, 前14 d, R1的CH4产量与R0无明显差别, R3的CH4产量提高了18.42%(第6 d);14 d后, R1的CH4产量逐渐高于R0;运行至28 d, 与R0相比, R1和R3的总CH4累积量分别提高了6.15%和3.99%, R2的CH4产量降低了21.25%.图 1(b)中, 第28 d, 与R0相比, R1和R2的CO2累积产量分别增加了7.14%和1.32%, R3降低了1.06%.在图 1(c)中, 与R0相比, R1和R3的产CH4速率分别提高了68.18%(第26 d)和51.35%(第4 d), 而R2的产CH4速率基本上均低于R0.
结果表明, 单纯投加GAC和MnO2, 前者促进了剩余污泥厌氧消化过程, 但也增加了系统CO2产量, 后者降低了厌氧系统的产CH4效率;联合投加GAC/MnO2, 可以缓解单独投加MnO2对剩余污泥厌氧消化过程的抑制作用, 而且一定时间内(前14 d)对剩余污泥厌氧消化过程的促进效果优于单纯投加GAC.投加GAC可以促进嗜乙酸产甲烷菌的代谢, 同时增加CO2的生成, Ryue等[13]在探究GAC对食品垃圾厌氧消化影响中也得到相同结论.Tian等[14]以葡萄糖为底物, 研究纳米MnO2对厌氧系统产CH4效能的影响, 发现通过Mn4+/Mn2+氧化还原过程释放的电子可以促进嗜氢产甲烷菌将CO2还原成CH4, 从而强化厌氧消化过程, 而本实验结果与此恰恰相反;这可能归咎于剩余污泥组分相对复杂, Mn2+与剩余污泥中的无机盐离子(如磷酸盐)反应生成沉淀, 导致MnO2催化性能失活, 生成的锰盐沉淀物堵塞了厌氧颗粒污泥代谢通道, 阻隔了产甲烷菌对酸性发酵产物的充分利用, 从而降低了产CH4效率.投加GAC/MnO2, GAC对锰盐沉淀物具有良好吸附作用, 可以减缓锰盐沉淀物对厌氧产CH4代谢通道的阻塞, 同时GAC在MnO2、厌氧微生物和剩余污泥之间具有桥梁作用, 其优良导电性促使Mn4+/Mn2+氧化还原过程产生的电子在酸性发酵过程和CO2还原成CH4中得以利用, 因此R3的累计产CH4量高于R0和R2, 同时累计CO2产量最低.
2.2 投加GAC、MnO2和GAC/MnO2对VFAs、蛋白质和多糖的影响 2.2.1 对VFAs的影响VFAs是厌氧消化过程主要中间代谢产物, 其产生与消耗速率可以用来评价厌氧消化效率[15];图 2为实验期间各组反应器内VFAs(包括乙酸、丙酸、丁酸和戊酸)的变化情况.

|
图 2 剩余污泥厌氧消化过程各组反应器VFAs的变化 Fig. 2 Changes in VFAs concentrations in each reactor during anaerobic digestion of waste activated sludge |
图 2中, R0、R1、R2和R3反应器VFAs浓度在8~10 d时达到最高, 实验后期VFAs浓度均降至100 mg·L-1左右.各组反应器VFAs在第2 d丁酸含量均较高, 第4 d丁酸含量降低, 而后至第14 d丁酸含量均较高.丁酸在系统内出现积累是厌氧产CH4受抑制的一个表征[16], 各组反应器在前4 d产生的丁酸均被较快降解, 表明系统存在有效的嗜氢产甲烷过程;6~12 d出现的丁酸积累意味着嗜氢产甲烷过程的受抑制, 14 d后随着代谢底物的消耗和有机酸生成速度的减弱, 反应器内各类有机酸含量逐渐降低.另外, R1、R2和R3的VFAs浓度在前10 d均高于R0, 单独或者联合投加GAC和MnO2可能对酸性发酵过程的电子转移有促进作用, 同时GAC和GAC/MnO2促进了产CH4过程和提高酸化效率.
2.2.2 对蛋白质和多糖的影响蛋白质和多糖是剩余污泥重要有机物, 作为厌氧消化的代谢基质, 其浓度变化可以间接反映厌氧消化速率[17, 18].图 3为各组反应器蛋白质和多糖的浓度变化情况.

|
图 3 剩余污泥厌氧消化过程中蛋白质和多糖浓度变化 Fig. 3 Changes of the concentration of proteins and polysaccharides in each reactor during anaerobic digestion of wasted activated sludge |
由图 3可见, R0、R1、R2和R3的多糖浓度有着近似的变化趋势, 在第4 d浓度最高, 而后出现不同程度的波动, 但整体呈下降趋势;R0和R2蛋白质浓度逐渐升高, 第22 d达到峰值, R3蛋白质浓度第4 d最高, 第8 d降到最低, 随后又逐渐升高.R1和R3中蛋白质和多糖浓度均低于R0, 尤其R1中蛋白质和多糖浓度均有大幅度降低, 相反R2的蛋白质和多糖浓度基本上均高于R0.投加GAC和GAC/MnO2均加速了系统的厌氧消化过程, 相较于R0, R1和R3多糖和蛋白质作为代谢底物被更多降解, 浓度也更低;R2由于产CH4过程受到抑制, 导致其蛋白质和多糖浓度高于R1和R3.
2.3 投加GAC、MnO2和GAC/MnO2对微生物活性的影响Cyt C是参与电子传递过程的氧化还原蛋白, 与厌氧污泥的微生物活性有关, 互营微生物可以通过Cyt C直接传递电子[19].辅酶F420是产CH4过程的关键辅酶[20];有关研究表明CH4的产生与INT-ETS活性呈正相关[21].为研究投加物对微生物活性影响, 分别测定第14 d和28 d各组反应器Cyt C浓度、INT-ETS活性和辅酶F420含量, 具体见图 4.
由图 4可见, 第14 d R3的Cyt C浓度为38.21 nmol·L-1, 远高于R0, R1的Cyt C浓度也高于R0, 而R2的Cyt C浓度小于R1和R3, 略高于R0, 这表明GAC和GAC/MnO2的投加明显促进了Cyt C的产生.单纯投加MnO2, 虽然R2的CH4产量和速率均低于R0, 但R2强化酸性发酵, 因此R2的Cyt C高于R0.第28 d, 代谢底物匮乏, R1、R2和R3的Cyt C浓度均低于R0.

|
图 4 投加不同材料对INT-ETS、辅酶F420和Cyt C的影响 Fig. 4 Changes in electron transfer activities, coenzyme F420, and Cyt C with the addition of materials |
第14 d, 与R0相比, R1、R2和R3的INT-ETS活性和辅酶F420含量均有所增加, 其中R3的INT-ETS活性和辅酶F420含量分别是R0的1.6倍和2.3倍, GAC和MnO2能提高微生物的INT-ETS活性和产甲烷菌关键酶的含量.第28 d, R1、R2和R3的INT-ETS活性和辅酶F420含量均低于R0, 其中R1、R2和R3的INT-ETS活性分别降低了16.05%、31.35%和30.67%;产生此现象的原因与Cyt C相同.对应的R1、R2和R3的产CH4效率均在14 d之前较高, 而在14 d之后CH4产量增长缓慢.总之, 在底物充分和后续代谢正常进行条件下, 投加GAC、MnO2和GAC/MnO2均可以提高微生物活性.
2.4 投加MnO2、GAC/MnO2系统锰离子和磷酸盐的变化为进一步探究GAC和MnO2影响剩余污泥厌氧消化作用机制, 实验期间监测了R2和R3反应器游离锰离子和磷酸盐的含量, 具体见图 5.

|
图 5 厌氧消化过程中锰离子和磷酸盐的释放 Fig. 5 Concentration of manganese ions and phosphate during anaerobic digestion |
由图 5(a)可见, R2和R3的锰离子浓度变化趋势相同, 在2 d内迅速增加, 第4 d达到最大, 之后迅速下降. 14 d后锰离子含量低于1 mg·L-1, 并一直维持在较低水平, 对应的R2和R3的产CH4效率也是在第14 d后开始下降.系统锰离子浓度逐步下降可能有以下3种途径:①随着酸性发酵的减弱, 系统pH升高, 抑制了MnO2向Mn2+转化;②锰离子与污泥释放的磷酸盐反应生成磷酸锰沉淀;③锰离子被活性污泥或GAC吸附[22].图 5(b)中, R2和R3磷酸盐含量逐渐升高, 厌氧发酵过程剩余污泥储存的磷不断释放, 锰离子与之结合生成磷酸锰沉淀[23];R3的锰离子浓度始终低于R2, 符合GAC吸附锰离子的假设.
MnO2是一种半导体, 与一些导电性好的材料共同作用, 可以强化污泥活性[24].GAC作为互营细菌和产甲烷菌之间的电子传递介质, 不仅可以促进功能菌的富集, 还可以促进DIET代谢途径[25].具有良好导电性和吸附性的GAC与作为“催化剂”的MnO2共同作用, 不仅缓解了磷酸锰沉淀对产CH4代谢通道的阻塞, 还克服了MnO2导电性弱的缺点, 与单纯投加GAC或MnO2相比, 联合投加GAC/MnO2的电子传递活性有了较大提高, 它们共同作用提高了剩余污泥厌氧消化效率.
基于颗粒污泥内部孔道可能被磷酸锰沉淀堵塞的推测, 对污泥样品进行SEM分析.R0厌氧颗粒污泥的表面呈现多孔、多层结构, 可以观察到清晰的通道孔结构[图 6(a)和6(b)], 而R2中几乎没有观察到通道[图 6(c)];R3中尽管有些通道被堵塞, 但其表面仍存在三维多孔结构[图 6(d)].说明GAC可以将磷酸盐和锰离子或者生成的磷酸锰沉淀吸附固定, 从而缓解磷酸锰沉淀的产生, 减缓厌氧颗粒污泥的代谢孔道被堵塞.

|
图 6 厌氧颗粒污泥的SEM图像 Fig. 6 SEM images of granular sludge from R0, R1, and R3 |
为了探究投加GAC和MnO2对反应器中微生物群落的影响, 采用高通量测序技术对各反应器中厌氧污泥微生物群落结构进行了分析, 具体见图 7.

|
图 7 反应器污泥中微生物细菌和古菌在门和属水平的相对丰度 Fig. 7 Relative abundance of sludge samples in each reactor at phylum and genus levels of bacteria and archaea |
在细菌门水平分布中, 主要有Firmicutes、Bacteroidetes、Actinobacteria和Chloroflexi等优势菌, 其中Firmicutes门占绝对优势地位, 在4个样品中的丰度均超过31%[图 7(a)].加入GAC和MnO2, 这些细菌始终保持优势地位, 但是其相对丰度有所改变.进一步地分析细菌的属水平[图 7(b)], R0的优势菌主要有Petrimonas(7.18%)和Syntrophomonas(7.03%), R2和R3的优势菌有所改变, Bacillus和Syntrophomonas是R2的优势菌, 相对丰度分别为14.69%和10.09%, R3的优势菌为Fonticella(13.67%)和Syntrophomonas(9.76%).Petrimonas是中温发酵微生物, 能够将复杂的有机物转化为乙酸[26].Syntrophomonas是利用丁酸的典型产氢产乙酸细菌, 可以将丁酸转化为乙酸, 为嗜乙酸产甲烷菌提供代谢底物.实验中丁酸的大量产生可能是刺激Syntrophomonas富集的原因之一, 同时Syntrophomonas的富集会促进乙酸的积累和嗜乙酸产甲烷菌Methanosaeta的生长.电活性微生物Geobacter在R0中的相对丰度仅为0.13%, 加入GAC和MnO2后, R1、R2和R3的相对丰度均有所增加.
在古菌门水平分布中, R0的优势菌为Euryarchaeota(64.68%), Euryarchaeota门在R1、R2和R3的相对丰度有大幅度的增加, 分别为95.35%、85.11%和92.77%[图 7(c)], 产甲烷菌属于Euryarchaeota门, GAC和MnO2促进了产甲烷菌的富集.古菌的属水平分布如图 7(d)所示, R0的优势菌属是Methanobacterium(60.37%), 在GAC和MnO2存在的条件下, 实验组的优势菌属发生了改变, Methanobacterium和Methanosaeta成为了R1、R2和R3的优势菌属, 其中Methanosaeta在R1、R2和R3的相对丰度从2.31%(R0)分别增加为47.04%、20.83%和31.69%.R2和R3中的Methanobacterium的相对丰度有所增加, 分别为69.81%和61.52%, Methanobacterium在R1中的相对丰度下降为34.65%.Methanobacterium是一种典型嗜氢产甲烷菌, 可以利用H2还原CO2产CH4, 在维持厌氧系统氢分压方面发挥着重要作用[27].Methanosaeta属于典型嗜乙酸产甲烷菌[28], 能够将厌氧代谢中间产物乙酸转化为CH4.投加GAC和MnO2促进了乙酸的转化和CO2的还原, 提高了产CH4效率, 降低了CO2产量.另外, GAC和MnO2促进了水解酸化, 生成的乙酸可以被富集的嗜乙酸产甲烷菌Methanosaeta利用.同时, 产甲烷菌Methanosaeta不仅可以利用乙酸生成CH4, 也可以通过DIET途径产CH4[29].大量的研究都证实GAC的导电性可以促进CH4的产生[30].在GAC体系中, Methanobacterium菌的丰度低于GAC/MnO2, 这表明嗜氢产甲烷菌丰度的增加是由MnO2的存在引起的.Geobacter是目前发现具有胞外电子传递能力的菌属, 也是能够与产甲烷菌通过DIET互营代谢合作的细菌之一[16].有研究发现Geobacter可以在GAC孔隙中富集, Methanosaeta在GAC表面富集, GAC可以作为Geobacter和Methanosaeta之间的电子连接促进污泥降解和CH4产量[31];这与R1和R3具有更高产CH4效率相一致.
3 结论(1) 投加GAC、MnO2和GAC/MnO2, 对剩余污泥厌氧消化系统的产酸发酵过程均有促进作用, 但是对系统产CH4影响不同, 其中投加GAC和GAC/MnO2对系统产CH4过程起到促进作用, 而单独投加MnO2对系统产CH4有抑制作用.
(2) GAC良好的导电性和Mn4+/Mn2+转换的催化剂作用, 提高了酸性发酵和产CH4过程的种间电子传递.但是单独投加MnO2时, 剩余污泥释放的磷酸盐和Mn2+生成沉淀堵塞了厌氧颗粒污泥代谢通道, 造成对系统产CH4代谢的抑制.
(3) 投加GAC和MnO2促进乙酸菌Syntrophomonas和产甲烷菌Methanobacterium富集, 进而可以改善细菌和产甲烷菌之间直接或间接的种间电子传递, 提高系统厌氧消化效率.
| [1] | Eskicioglu C, Prorot A, Marin J, et al. Synergetic pretreatment of sewage sludge by microwave irradiation in presence of H2 O2 for enhanced anaerobic digestion[J]. Water Research, 2008, 42(18): 4674-4682. DOI:10.1016/j.watres.2008.08.010 |
| [2] |
董慧峪, 季民. 剩余污泥厌氧消化甲烷生成势与产甲烷菌群多样性的比较研究[J]. 环境科学, 2014, 35(4): 1421-1427. Dong H Y, Ji M. Comparative study on biological methane potential and methanogen biodiversity in the anaerobic digestion of excess sludge[J]. Environmental Science, 2014, 35(4): 1421-1427. |
| [3] | Wei W, Cai Z Q, Fu J, et al. Zero valent iron enhances methane production from primary sludge in anaerobic digestion[J]. Chemical Engineering Journal, 2018, 351: 1159-1165. DOI:10.1016/j.cej.2018.06.160 |
| [4] | Li Y, Chen Y G, Wu J. Enhancement of methane production in anaerobic digestion process:a review[J]. Applied Energy, 2019, 240: 120-137. DOI:10.1016/j.apenergy.2019.01.243 |
| [5] | Wang G J, Li Q, Gao X, et al. Sawdust-derived biochar much mitigates VFAs accumulation and improves microbial activities to enhance methane production in thermophilic anaerobic digestion[J]. ACS Sustainable Chemistry & Engineering, 2019, 7(2): 2141-2150. |
| [6] | Martins G, Salvador A F, Pereira L, et al. Methane production and conductive materials:a critical review[J]. Environmental Science & Technology, 2018, 52(18): 10241-10253. |
| [7] | Wang F, Dai H X, Deng J G, et al. Manganese oxides with rod-, wire-, tube-, and flower-like morphologies:highly effective catalysts for the removal of toluene[J]. Environmental Science & Technology, 2012, 46(7): 4034-4041. |
| [8] | 国家环境保护总局. 水和废水监测分析方法[M]. (第四版).北京: 中国环境科学出版社, 2002: 121-124. |
| [9] | Yang B, Xu H, Liu Y B, et al. Role of GAC-MnO2 catalyst for triggering the extracellular electron transfer and boosting CH4 production in syntrophic methanogenesis[J]. Chemical Engineering Journal, 2019. DOI:10.1016/j.cej.2019.123211 |
| [10] | Zhu T T, Zhang Y B, Quan X, et al. Effects of an electric field and iron electrode on anaerobic denitrification at low C/N ratios[J]. Chemical Engineering Journal, 2015, 266: 241-248. DOI:10.1016/j.cej.2014.12.082 |
| [11] | DuBois M, Gilles K A, Hamilton J K, et al. Colorimetric method for determination of sugars and related substances[J]. Analytical Chemistry, 1956, 28(3): 350-356. DOI:10.1021/ac60111a017 |
| [12] | Walker J M. The protein protocols handbook (3rd ed.)[M]. New York: Humana Press, 2009. |
| [13] | Ryue J, Lin L, Liu Y, et al. Comparative effects of GAC addition on methane productivity and microbial community in mesophilic and thermophilic anaerobic digestion of food waste[J]. Biochemical Engineering Journal, 2019, 146: 79-87. DOI:10.1016/j.bej.2019.03.010 |
| [14] | Tian T, Qiao S, Yu C, et al. Distinct and diverse anaerobic respiration of methanogenic community in response to MnO2 nanoparticles in anaerobic digester sludge[J]. Water Research, 2017, 123: 206-215. DOI:10.1016/j.watres.2017.06.066 |
| [15] | Scoma A, Varela-Corredor F, Bertin L, et al. Recovery of VFAs from anaerobic digestion of dephenolized olive mill wastewaters by electrodialysis[J]. Separation and Purification Technology, 2016, 159: 81-91. DOI:10.1016/j.seppur.2015.12.029 |
| [16] | Zhang J S, Zhao W Q, Zhang H W, et al. Recent achievements in enhancing anaerobic digestion with carbon- based functional materials[J]. Bioresource Technology, 2018, 266: 555-567. DOI:10.1016/j.biortech.2018.07.076 |
| [17] | Yang X J, Nie J M, Wang D, et al. Enhanced hydrolysis of waste activated sludge for methane production via anaerobic digestion under N2-nanobubble water addition[J]. Science of the Total Environment, 2019, 693: 133524. DOI:10.1016/j.scitotenv.2019.07.330 |
| [18] | Basuvaraj M, Fein J, Liss S N. Protein and polysaccharide content of tightly and loosely bound extracellular polymeric substances and the development of a granular activated sludge floc[J]. Water Research, 2015, 82: 104-117. DOI:10.1016/j.watres.2015.05.014 |
| [19] | Nagarajan H, Embree M, Rotaru A E, et al. Characterization and modelling of interspecies electron transfer mechanisms and microbial community dynamics of a syntrophic association[J]. Nature Communications, 2013, 4: 2809. DOI:10.1038/ncomms3809 |
| [20] |
苏润华, 丁丽丽, 任洪强. 纳米零价铁(NZVI)对厌氧产甲烷活性、污泥特性和微生物群落结构的影响[J]. 环境科学, 2018, 39(7): 3286-3296. Su R H, Ding L L, Ren H Q. Impact of nano zero-valent iron (NZVI) on methanogenic activity, physiological traits, and microbial community structure in anaerobic digestion[J]. Environmental Science, 2018, 39(7): 3286-3296. |
| [21] | Zhang F, Hou J, Miao L Z, et al. Chlorpyrifos and 3, 5, 6-trichloro-2-pyridinol degradation in zero valent iron coupled anaerobic system:performances and mechanisms[J]. Chemical Engineering Journal, 2018, 353: 254-263. DOI:10.1016/j.cej.2018.07.136 |
| [22] | Tian T, Qiao S, Yu C, et al. Effects of nano-sized MnO2 on methanogenic propionate and butyrate degradation in anaerobic digestion[J]. Journal of Hazardous Materials, 2019, 364: 11-18. DOI:10.1016/j.jhazmat.2018.09.081 |
| [23] | Li L, Pang H L, He J G, et al. Characterization of phosphorus species distribution in waste activated sludge after anaerobic digestion and chemical precipitation with Fe3+ and Mg2+[J]. Chemical Engineering Journal, 2019, 373: 1279-1285. DOI:10.1016/j.cej.2019.05.146 |
| [24] |
万厚钊, 缪灵, 徐葵, 等. MnO2基超级电容器电极材料[J]. 化工学报, 2013, 64(3): 801-813. Wan H Z, Miao L, Xu K, et al. Manganese oxide-based electrode behavior as materials for electrochemical supercapacitors[J]. CIESC Journal, 2013, 64(3): 801-813. DOI:10.3969/j.issn.0438-1157.2013.03.004 |
| [25] | Lei Y Q, Sun D Z, Dang Y, et al. Metagenomic analysis reveals that activated carbon aids anaerobic digestion of raw incineration leachate by promoting direct interspecies electron transfer[J]. Water Research, 2019, 161: 570-580. DOI:10.1016/j.watres.2019.06.038 |
| [26] | Li Y, Ren C Y, Zhao Z S, et al. Enhancing anaerobic degradation of phenol to methane via solubilizing Fe(Ⅲ) oxides for dissimilatory iron reduction with organic chelates[J]. Bioresource Technology, 2019, 291: 121858. DOI:10.1016/j.biortech.2019.121858 |
| [27] | Zhang Z H, Gao P, Cheng J Q, et al. Enhancing anaerobic digestion and methane production of tetracycline wastewater in EGSB reactor with GAC/NZVI mediator[J]. Water Research, 2018, 136: 54-63. DOI:10.1016/j.watres.2018.02.025 |
| [28] |
杨波, 徐辉, 冯修平, 等. SCAR处理城市生活污水的效能及其微生物群落动态分析[J]. 环境科学, 2017, 38(5): 2021-2029. Yang B, Xu H, Feng X P, et al. Analysis on performance and microbial community dynamics of a strengthen circulation anaerobic reactor treating municipal wastewater[J]. Environmental Science, 2017, 38(5): 2021-2029. |
| [29] | Mei R, Nobu M K, Narihiro T, et al. Novel Geobacter species and diverse methanogens contribute to enhanced methane production in media-added methanogenic reactors[J]. Water Research, 2018, 147: 403-412. DOI:10.1016/j.watres.2018.10.026 |
| [30] |
周婷, 余林鹏, 符力, 等. 微生物直接电子传递:甲烷代谢古菌研究进展[J]. 应用与环境生物学报, 2018, 24(5): 1032-1040. Zhou T, Yu L P, Fu L, et al. Microbial direct electron transfer:advances in its study in the metabolism of methane by archaea[J]. Chinese Journal of Applied & Environmental Biology, 2018, 24(5): 1032-1040. |
| [31] | Yang Y F, Zhang Y B, Li Z Y, et al. Adding granular activated carbon into anaerobic sludge digestion to promote methane production and sludge decomposition[J]. Journal of Cleaner Production, 2017, 149: 1101-1108. DOI:10.1016/j.jclepro.2017.02.156 |