 2020, Vol. 41
2020, Vol. 41


2. 合肥工业大学纳米矿物与污染控制安徽普通高校重点实验室, 合肥 230009
2. Key Laboratory of Nanominerals and Pollution Control of Anhui Higher Education Institutes, Hefei University of Technology, Hefei 230009, China
氮是重要的生源要素, 因此氮循环在整个地球环境系统中扮演中极为重要的角色, 维系着人类及其它地球生命系统的生存和发展.由于近些年人类活动的不断加剧, 导致全球新增“活性”氮量大幅增加, 进而导致全球氮循环产生严重失调的倾向.众所周知, 过量的氮输入到生态系统可能产生严重的生态危害性, 如农作物氮的富集、水体富营养化和水生态退化等.因此, 氮素污染问题引起人们越来越多的关注[1, 2].反硝化过程可以将NO3-从生态系统中完全去除, 因此反硝化技术被认为是解决水体氮素污染的重要途径和机制[3, 4].水塘洼地是河湖水系统的重要组成部分, 也是非点源氮磷滞留和转化的重要场所.目前, 针对水体沉积物反硝化问题的研究, 主要集中在大型河口滩涂湿地、湖泊、水库等大尺度缓流水体, 以及不同尺度河流沟渠水体沉积物等[5~7], 而对小型水塘洼地湿地类型的重视程度相对不足.从水力特征看, 由于空间尺度小及水体深度不大, 水塘洼地常因缺乏有效的水源补给而表现出一定的干湿交替现象, 这与湖库、河流等存在很大差别.水塘洼地是河湖水系常见的水体类型, 由于数量较大, 对调控陆地径流非点源污染负荷发挥着重要作用.因此, 开展水塘洼地沉积物反硝化潜力及其影响因素分析, 对推动和促进流域水环境管理具有重要意义.
巢湖是我国五大淡水湖泊之一, 湖泊富营养化颇为严重, 高负荷的氮素污染状况成为困扰巢湖水体富营养化治理的难题之一[8].据报道, 巢湖流域非点源氮磷污染负荷已超过点源污染贡献水平, 因此如何有效削减非点源氮磷入湖负荷, 成为巢湖水环境治理的一项颇为棘手的难题.巢湖流域大多为低矮丘陵岗地, 河流水系发达, 水塘洼地数量众多.随着巢湖富营养化治理及环湖景观带建设的不断推进, 环湖周边退耕还湖、退耕还湿初见成效, 沿湖村落的搬迁使得滨岸地带水塘洼地的数量显著增加, 如何有效发挥这种水体类型的氮磷滞留功效值得探究.本研究拟以巢湖西半湖北侧滨岸带中村落搬迁后的三处典型水塘洼地为对象, 通过对表层沉积物碳氮源添加的实验室培养分析, 解析沉积物反硝化潜力及碳氮的限制性, 并识别主要环境影响因素, 以期为巢湖周边水塘洼地沉积物反硝化潜力提升提供依据.
1 材料与方法 1.1 研究区概况巢湖位于安徽省合肥市境内, 是我国五大淡水湖泊之一.巢湖流域属于北亚热带湿润季风气候区, 年均气温约16℃, 年均降水量约1 100 mm, 其中5~8月降水量约占全年总降水量的55%.近年来, 巢湖水体富营养化日趋严重, 水华暴发频频发生.巢湖流域河流水系发达, 水塘洼地数量众多.在近10多年巢湖环湖景观带建设, 特别是西半湖景观休闲区建设过程中, 沿湖村落搬迁后遗留的水塘、洼地也都经水景观改造逐步转变为景观湿地的重要组成.
本研究选择的三处水塘洼地位于巢湖西半湖北岸大堤外侧, 介于塘西河和十五里河入湖河口之间, 北邻巢湖景观大道环湖北路(见图 1).该区域原为塘西村所在地, 目前已改造建设成巢湖西半湖景观休闲带.这里, 水塘1近似于圆形, 长宽约60 m, 滨水带芦苇和蒲草生长茂盛;水塘2长约800 m, 最宽约80 m, 两端为莲藕种植区, 周边浅水区芦苇、蒲草生长茂盛, 岸上滨水带大型水生/湿地植物生长浓密;洼地近似椭圆形, 长约90 m, 最宽处约60 m, 底质主要为黏质土壤, 水深较浅, 冬季常处于干涸状态, 大型植物种类单一, 基本都为浓密的蒲草丛覆盖.

|
图 1 采样区域示意 Fig. 1 Sketch-map of the distribution of the sampling areas |
表 1为整个采样期间水塘洼地水质数据的统计平均值.可以看出, 3个水塘洼地水体水质状况基本接近.TN平均浓度高达14.43mg·L-1, 超过《地表水环境质量标准》(GB 3838-2002)Ⅴ类水标准阈值6.22倍, NO3--N浓度更是显著高于NH4+-N;TP平均浓度为0.54mg·L-1, 略高于V类水标准, 且SRP占TP百分比平均为55.6%.
|
|
表 1 河流水质统计结果 Table 1 Mean value of water quality indices in the sampling areas |
1.2 样品采集与分析测试
根据3个水体植被类型及组成分布情况, 将其划分为水塘芦苇、水塘蒲草、洼地蒲草等3种类型植物丛, 并设置9个采样点(表 1).为增强采样点位的代表性, 每个采样点均布置在芦苇或蒲草占绝对优势且密集程度相对较高的区域, 相应地覆盖面积基本都在8 m2以上. 2018年10月、12月及2019年2月、4月和6月, 采集表层2~5 cm沉积物样及上覆水样.采样期间水塘采样点处水深基本维持在30~50 cm, 洼地水深仅为10~20 cm, 且在12月和翌年2月出现干涸情况.每个采样点采集沉积物约1 000 g, 现场测定pH、水温(T)、总溶解固体(TDS)、电导率(EC)和氧化还原电位(ORP).一部分新鲜沉积物用于反硝化速率测定, 其余部分放置于阴凉通风处自然风干, 经研磨过100目尼龙筛, 密封保存以备沉积物理化性质分析.
沉积物分析项目包括TN、TP、有机质(OM)、NH4+-N、NO3--N和pH等, 水样包括TN、TP、OM、NH4+-N、NO3--N、pH、溶解反应性磷(SRP)和COD等指标.具体测定方法参见文献[9].
1.3 实验方案设计称取相当于1 g烘干样的新鲜沉积物样于50 mL离心管中, 采用硝酸盐剩余量法[10~12]测定沉积物反硝化速率.针对碳、氮外源添加设置3种情形, 即单独添加碳、氮以及两者共同添加, 每种情形均设置两个梯度(见表 2).这里, 2018年10月未就单独添加碳情形开展培养实验.为便于比较, 针对上述3种情形设置一组空白对照样(添加同体积20 mL上覆水).此外, 对于上述每种处理方式均设置3个平行, 并以平均值代表测试结果.
|
|
表 2 反硝化实验方案设计1) Table 2 Specific treatments for denitrification |
将装有新鲜沉积物的离心管加塞密封, 放置在恒温培养箱30℃环境条件下培养48 h;然后采用3 500 r·min-1离心10 min, 分离过滤后测定NO3--N浓度, 并由滤液体积, 估算剩余的NO3--N量.为确定新鲜沉积物的NO3--N含量, 称取相同质量的新鲜样, 加入20 mL 4 mol·L-1的氯化钾溶液, 采用3 500 r·min-1转速离心10 min, 过滤后测定滤液中NO3--N浓度, 再由滤液体积, 计算单位质量沉积物的NO3--N量, 并将该测定值加上添加NO3--N量作为初始值, 采用式(1)计算反硝化速率:
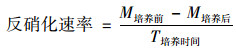
|
(1) |
式中, M培养前表示培养前单位质量沉积物的NO3--N量(mg·kg-1);M培养后表示培养后单位质量沉积物的NO3--N量(mg·kg-1);T培养时间表示沉积物在恒温培养箱的培养时间(h).
1.4 数据处理采用单因素方差分析(one-way ANOVA), 解析3种植物丛沉积物反硝化速率的差异性;采用Spearman系数表征反硝化速率与上覆水水质指标、沉积物理化指标的相关性.相关数据处理及图形绘制由Excel软件完成.
2 结果与分析 2.1 沉积物氮磷污染特征表 3为整个采样期间3种植物丛沉积物基本理化指标的统计平均值.可以看出, 3个水体不同植物丛中, 除OM外, 其它各项指标相差不大.总体上, 洼地蒲草的OM含量明显低于水塘芦苇、水塘蒲草.另外, 洼地蒲草和水塘蒲草的最大变异系数分别为10.84%和12.38%, 相应指标都为NH4+-N;水塘芦苇中TP的变异系数最大, 为16.81%.因此, 3种植物丛沉积物基本理化性质的时空异质性不大.
|
|
表 3 沉积物基本理化指标统计结果 Table 3 Mean values of physiochemical indices in sediments |
3种植物丛沉积物的TN和TP均值分别为1 595.84 mg·kg-1和462.044 mg·kg-1, 均低于巢湖表层沉积物TN和TP含量(分别为1 737.00 mg·kg-1和691.00 mg·kg-1)[13], 显著低于邻近的十五里河沉积物(分别为1 884.23 mg·kg-1和1 068.07 mg·kg-1)[14].采样区沉积物NH4+-N含量均值为444.18 mg·kg-1, 明显低于十五里河氨氮含量(597.52mg·kg-1), 但高于巢湖表层沉积物中氨氮含量(春季蓝藻暴发前:305.52 mg·kg-1)[15]. 3种植物丛沉积物NO3--N含量均值为142.05mg·kg-1, 显著高于十五里河沉积物(58.38mg·kg-1).水塘芦苇、水塘蒲草OM均值分别为7.50%、7.40%, 显著高于巢湖表层沉积物中OM含量(5.86%)[13].
2.2 反硝化速率及对外源碳氮的响应 2.2.1 对照样反硝化速率与碳氮梯度情形相比, 不添加外源碳氮的沉积物对照样测得的反硝化速率可能更接近于采样点真实的反硝化潜力状况. 3种植物丛沉积物对照样反硝化速率的变化情况, 见图 2.显然, 整个采样期间, 3种植物丛的反硝化速率变化规律一致, 且都表现为2月最低, 6月最高.整个研究期间, 3种水生植物丛沉积物对照样反硝化速率总体上不存在显著的差异性(P>0.626), 但在不同月份情况则稍有不同.其中, 10月洼地蒲草和水塘芦苇沉积物对照样反硝化速率呈极显著差异性(P=0.001)、洼地蒲草和水塘蒲草之间也呈极显著差异性(P=0.008);4月洼地蒲草与水塘芦苇沉积物对照样反硝化速率呈现显著差异性(P=0.028), 其它月份不同类型沉积物对照样反硝化速率的差异性不明显.

|
图 2 对照样反硝化速率变化特征 Fig. 2 Variations in the denitrification rates in the controls during the sampling periods |
洼地蒲草对照样反硝化速率为(6.47±2.76)mg·(kg·h)-1, 水塘芦苇反硝化速率为(6.97±2.86)mg·(kg·h)-1, 水塘蒲草对照样反硝化速率值为(6.76±2.82)mg·(kg·h)-1.李如忠等[12]采用硝酸盐剩余量法分别对合肥城郊小溪流水质状况良好段和严重污染段沉积物反硝化速率进行了测算, 得到反硝化速率分别为2.53 mg·(kg·h)-1、3.32 mg·(kg·h)-1.秦如彬等[11]也以该方法测算了合肥城郊受生活污水严重污染的沟渠沉积物反硝化潜力, 得到反硝化速率为10.59 mg·(kg·h)-1;测算得到受污水厂尾水影响的溪流沉积物反硝化速率为6.05 mg·(kg·h)-1.牟晓杰等[10]在对闽江河口芦苇湿地的研究中, 得到污染严重滩涂湿地的反硝化速率约0.74~2.69mg·(kg·h)-1.不难推断, 巢湖滨岸水塘洼地沉积物反硝化潜力与这些溪流、湿地基本相当.
2.2.2 对外源氮的响应在单独添加外源氮情形下, 针对氮浓度梯度N1和N2, 不同采样时间3种植物丛沉积物反硝化速率变化情况, 见图 3.与图 2中相应的空白对照样相比, 随着外源氮浓度的增加, 3种植物丛沉积物反硝化速率总体上表现出增大的趋势, 但也出现了异常情况.具体地说, 10月洼地蒲草沉积物反硝化速率N1梯度时不仅没有增加反而下降, N2梯度时下降幅度更显著; 6月洼地蒲草植物丛依旧延续这一特点.而且, 6月其它2种植物丛N2梯度相应的反硝化速率也远低于对照样和N1梯度.由于大部分情况下3种植物丛沉积物反硝化速率都表现出随着氮的投加量增加而快速增大的变化趋势, 表明硝态氮是植物丛沉积物反硝化过程的限制因素[16].但上述异常情况也意味这种限制性不是固定不变的, 而是具有一定的时间(或季节)变化性, 尤以洼地蒲草植物丛沉积物表现更为明显.

|
图 3 不同氮梯度下沉积物反硝化速率变化情况 Fig. 3 Variations in the denitrification rates along the nitrogen gradient |
总体上, N1梯度下洼地蒲草与水塘芦苇沉积物反硝化速率存在极显著差异性(P=0.007), 洼地蒲草与水塘蒲草之间也表现出一定的差异性(P=0.074), 而水塘芦苇与水塘蒲草的差异性不显著(P=0.234);N2梯度下3种植物丛反硝化速率没有表现出明显的差异性(P>0.261).
2.2.3 对外源碳的响应图 4为单独添加外源碳情形下3种植物丛沉积物反硝化速率变化情况.与图 2的对照样反硝化速率相比, 单独增加外源碳时, 整个采样期间3种植物丛沉积物反硝化速率均有大幅度下降, 表明3种植物丛沉积物反硝化进程受到碳的抑制.

|
图 4 不同碳梯度下沉积物反硝化速率变化情况 Fig. 4 Variations in the denitrification rates along the carbon gradient |
洼地蒲草在2018年2月和4月出现负值, 水塘芦苇和水塘蒲草在12月反硝化速率值也出现了负值.差异性分析结果表明, 外加碳源情形下3种植物类型沉积物反硝化速率之间不存在显著差异性(P>0.267).
2.2.4 对外加碳氮源的响应图 5为外源碳氮同时添加时, 不同碳氮梯度下3种植物丛沉积物反硝化速率值随采样时间而变化的情况.与图 2的对照样反硝化速率相比, N1+C1梯度的沉积物反硝化速率都有了较为明显的提高;与N1+C1梯度相比, 总体上N2+C2梯度的沉积物反硝化速率继续表现出较快的增长态势, 表明外源碳氮同时添加可以促进3种植物丛沉积物的反硝化作用.但在N2+C2梯度也出现了明显的异常情况, 即10月洼地蒲草、6月水塘蒲草植物丛沉积物反硝化速率反而低于N1+C1梯度, 但却依旧高于对照样反硝化速率值.这里, 由于实验中滴入的碳和氮溶液体积相同, 因此N1+C1和N2+C2梯度的C:N比值分别为15:1和25:1.换言之, 大部分情形下碳氮比为25:1时的反硝化速率较15:1时更高.

|
图 5 不同碳氮梯度下沉积物反硝化速率变化情况 Fig. 5 Variations in the denitrification rates along the dual nutrient gradient |
与N1+C1梯度相比, N2+C2梯度下各采样时间3种植物丛沉积物反硝化速率的差异性相对更为明显一些, 但无论是N1+C1梯度还是N2+C2梯度, 3种植物丛沉积物反硝化速率之间的差异性均达不到显著性水平(P>0.417).
与单添加氮(图 3), 特别是单添加碳(图 4)相比, 同时添加碳氮时各植物丛沉积物反硝化速率提高更显著, 表明碳氮同时添加更有利于巢湖滨岸水塘洼地植物丛沉积物反硝化进程.
2.3 反硝化速率与环境因素的关系对照样反硝化速率与上覆水水质指标的Spearman相关系数见表 4.这里, 3种植物丛的对照样反硝化速率均与T和NH4+-N浓度呈极显著相关关系, 水塘蒲草沉积物还与NO3--N表现出极显著相关性, 水塘芦苇和水塘蒲草都与TN表现出极显著相关性.虽然洼地蒲草对照样反硝化速率与SRP呈现显著相关, 但总体而言与磷素关系并不密切, 与COD浓度关系似乎更弱.
|
|
表 4 对照样反硝化速率与水质指标Spearman相关系数1) Table 4 Spearman correlation coefficients between denitrification rates and water quality indexes |
对照样反硝化速率与沉积物理化指标的Spearman相关系数, 见表 5.显然, 与上覆水水质指标相比, 沉积物理化指标与对照样反硝化速率的相关关系较弱, 甚至不存在显著相关性.
|
|
表 5 对照样反硝化速率与沉积物理化指标Spearman相关系数 Table 5 Spearman correlation coefficients between denitrification rates and sediment physiochemical indexes |
3 讨论
根据外源氮梯度下沉积物反硝化速率的变化情况, 初步判定巢湖滨岸3个水塘洼地沉积物反硝化存在氮限制性.但由图 3也可以看出, 并不是所有植物丛沉积物的高氮浓度反硝化速率都较低氮浓度情形的反硝化速率更高, 尤其是2019年6月高氮浓度时3种植物丛沉积物反硝化速率较低浓度下降幅度在90%以上, 洼地蒲草甚至还出现了负值.笔者以为, 这可能是由于水分、碳源或其它因素转变成为反硝化限制性因素的缘故.实际上, Mulholland等[17]在对美国72条河流沉积物反硝化潜力的研究中, 也发现当硝酸盐含量过高时, 反硝化速率反而随着硝酸盐浓度的增加而下降.本研究中, 单独添加碳源, 反硝化受到了明显的抑制作用, 这可能是一方面采样区域为芦苇/蒲草浓密植物区, 沉积物中经年存在大量的腐殖质植物残体, 有机质含量较为丰富;另一方面, 氮限制性的存在表明反硝化底物硝态氮的不足, 此时进一步投入有机碳就会加剧硝态氮的不足, 进而导致碳限制性的发生.当然, 有机碳对反硝化过程的影响也存在很大的不确定性和动态变化性, 所得结论往往也不一致.如Herrman等[18]在对美国东北部农业源头溪流沉积物反硝化作用影响因素的研究中发现, 碳浓度的改变对沉积物反硝化速率没有影响, 作者认为这可能与研究溪流有着丰富的DOC和沉积碳有关.高雪梅等[19]在对不同量的碳输入梯度下果园排水沟底泥氮素反硝化速率的研究中发现, 加入碳能使反硝化速率有一定的增加.从生态化学计量学观点来看, 生物体对碳氮等生源物质的利用往往遵从一定的化学计量关系, 也就是说, 服从一定的碳氮比.因此, 碳氮比往往被视为控制反硝化脱氮速率的重要因素.本研究中, 根据外源碳情形的分析, 有机碳(葡萄糖)不是限制性因素而是抑制因素, 但当碳氮同时添加时情况明显发生了转变, 大部分情形下碳氮比为25:1时的反硝化速率较15:1时更高, 表明碳氮比为25:1可能更适合3个水塘洼地沉积物反硝化[20].因此, 在设计反硝化速率的影响因素时, 不能将碳和氮的反硝化效应生硬割裂开来, 需要从整体上综合考虑两者的影响, 即从碳氮比角度权衡碳氮的添加.
图 4的外加碳情形下部分反硝化速率值出现了负值, 尤其是洼地蒲草.笔者以为, 这可能与洼地蒲草处于干湿交替状态有很大关系.根据采样记录, 2018年12月和翌年2月, 洼地蒲草采样区处于干涸状态, 塘底底质直接暴露在空气中, 导致表层土壤的含氧量高, 而水塘中采样点所在的芦苇、蒲草经年被30 cm以上水体覆盖, 沉积物的含氧量相对不足.由于直接挖取表层沉积物且密封装袋, 当天返回实验室后立即投入培养实验, 使得洼地蒲草底质中残存的O2含量相对较高.本研究在培养实验中未排空离心管上部的空气, 加之底质中残存O2, 使得培养过程中可能发生一定的硝化作用.而高浓度碳的抑制作用本身导致通过反硝化途径损失的硝态氮量明显降低, 有时甚至低于残存O2经硝化作用生成的硝态氮量, 从而使得部分反硝化速率测定结果出现了负值.牟晓杰等[10]在以硝酸盐剩余量法对人为干扰活动(养殖塘、污染物排放和农业活动)对湿地土壤硝化-反硝化潜力影响的研究中, 测得水稻田、养殖场土壤反硝化速率最低范围也出现了负值情况.当前国内外普遍采用乙炔抑制法, 利用乙炔抑制N2O向N2转化, 将反硝化产物控制为N2O, 测定N2O的产量来推算反硝化速率[21~23].但硝酸盐剩余量法操作简单, 安全方便, 在对沉积物反硝化潜力及影响因素的初步分析中, 仍具有显著的优越性.
对照样反硝化速率与上覆水水质指标T、NH4+-N存在显著的相关关系, 这与大多数研究结论一致[5].但对照样反硝化速率与水质指标中NO3--N含量呈负相关, 似乎与人们的主观认知存在出入.事实上, 沉积物反硝化速率与水相中NO3--N浓度关系也很复杂, 存在很多不确定性.如汪银龙等[24]从细菌角度研究汾河下游水体nirS型反硝化细菌群落组成与无机氮的关系, 结果发现反硝化细菌的优势菌属红细菌属Rhodobacter、陶厄氏菌属Thauera与水质指标中的NO3--N、NO2--N含量呈负相关.本研究中, 对照样反硝化速率与沉积物化学性质相关性较弱.有学者指出, 反硝化速率与沉积物NO3--N、NH4+-N含量存在正相关关系[25, 26].但也有学者发现反硝化速率不受硝态氮、有机质或者磷的影响, 如李飞跃等[27]的研究发现反硝化速率与土壤硝态氮、氨氮浓度不具有显著的相关关系, 牟晓杰等[10]针对闽江河口湿地的研究也发现, 土壤有机质、TN、TP和可溶性碳含量以及电导率等与土壤硝化-反硝化作用不具有显著相关关系.此外, 王静等[5]在对三峡库区小江支流蓄水和泄水期沉积物硝化反硝化特征的研究中发现, 沉积物反硝化速率与沉积物理化指标(TN、NH4+-N、NO3--N)呈显著负相关.
4 结论(1) 洼地蒲草、水塘芦苇、水塘蒲草3种植物丛沉积物对照样反硝化速率随时间变化较为明显, 且彼此间总体上不存在显著差异性.
(2) 外源氮的添加促进了3种植物丛沉积物反硝化进程, 但随着氮浓度的增加, 少数情况下沉积物反硝化速率反而下降.
(3) 外源碳的添加对3种植物丛沉积物反硝化速率都起到明显的抑制作用, 尤以洼地蒲草沉积物的抑制作用更为明显.
(4) 碳氮同时添加较单独添加碳或氮情形更有利于沉积物反硝化进程, 且碳氮比为25:1时的反硝化速率较15:1时更高.
| [1] | Racchetti E, Bartoli M, Soana E, et al. Influence of hydrological connectivity of riverine wetlands on nitrogen removal via denitrification[J]. Biogeochemistry, 2011, 103(1-3): 335-354. DOI:10.1007/s10533-010-9477-7 |
| [2] | Soana E, Balestrini R, Vincenzi F, et al. Mitigation of nitrogen pollution in vegetated ditches fed by nitrate-rich spring waters[J]. Agriculture, Ecosystems & Environment, 2017, 243: 74-82. |
| [3] | Roley S S, Tank J L, Grace M R, et al. The influence of an invasive plant on denitrification in an urban wetland[J]. Freshwater Biology, 2018, 63(4): 353-365. DOI:10.1111/fwb.13073 |
| [4] | Unda-Calvo J, Martínez-Santos M, Ruiz-Romera E, et al. Implications of denitrification in the ecological status of an urban river using enzymatic activities in sediments as an indicator[J]. Journal of Environmental Sciences, 2019, 75: 255-268. DOI:10.1016/j.jes.2018.03.037 |
| [5] |
王静, 刘洪杰, 雷禹, 等. 三峡库区小江支流沉积物硝化反硝化速率在蓄水期和泄水期的特征[J]. 环境科学, 2107, 38(3): 946-953. Wang J, Liu H J, Lei Y, et al. Nitrification and denitrification rates in a small tributary, Nanhe River, of three gorge dam reservoir during water collection and release events[J]. Environmental Science, 2107, 38(3): 946-953. |
| [6] |
李家兵, 张党玉, 吴春山, 等. pH对闽江河口湿地沉积物氮素转化关键过程的影响[J]. 水土保持学报, 2017, 31(1): 272-278. Li J B, Zhang D Y, Wu C S, et al. Effects of pH on the key nitrogen transformation processes of the wetland sediment in the min river estuary[J]. Journal of Soil and Water Conservation, 2017, 31(1): 272-278. |
| [7] | Bellinger B J, Jicha T M, Lehto L P, et al. Sediment nitrification and denitrification in a lake superior estuary[J]. Journal of Great Lakes Research, 2014, 40(2): 392-403. DOI:10.1016/j.jglr.2014.03.012 |
| [8] |
王菁晗, 何吕奇姝, 杨成, 等. 太湖、巢湖、滇池水华与相关气象、水质因子及其响应的比较(1981-2015年)[J]. 湖泊科学, 2018, 30(4): 897-906. Wang J H, He L Q S, Yang C, et al. Comparison of algal bloom related meteorological and water quality factors and algal bloom conditions among Lakes Taihu, Chaohu, and Dianchi (1981-2015)[J]. Journal of Lake Sciences, 2018, 30(4): 897-906. |
| [9] | 国家环境保护总局. 水和废水监测分析方法[M]. (第四版). 北京: 中国环境科学出版社, 2002. |
| [10] |
牟晓杰, 刘兴土, 仝川, 等. 人为干扰对闽江河口湿地土壤硝化-反硝化潜力的影响[J]. 中国环境科学, 2013, 33(8): 1413-1419. Mou X J, Liu X T, Tong C, et al. Effects of human disturbance on nitrification and denitrification potential in the min river estuarine wetland[J]. China Environmental Science, 2013, 33(8): 1413-1419. |
| [11] |
秦如彬, 李如忠, 高苏蒂, 等. 城乡交错带典型溪流沟渠沉积物氮污染特征及硝化-反硝化潜力[J]. 环境科学, 2017, 38(3): 936-945. Qin R B, Li R Z, Gao S D, et al. Pollution characteristics and nitrification and denitrification potential of superficial sediments from streams in an urban-rural fringe[J]. Environmental Science, 2017, 38(3): 936-945. |
| [12] |
李如忠, 郑侠, 高苏蒂, 等. 人为扰动背景下城市边缘溪流底质硝化-反硝化潜力分析[J]. 环境科学, 2017, 38(11): 4598-4606. Li R Z, Zheng X, Gao S D, et al. Nitrification and denitrification potential of benthic sediments in a suburban stream under intense human disturbance scenarios[J]. Environmental Science, 2017, 38(11): 4598-4606. |
| [13] |
苗慧, 沈峥, 蒋豫, 等. 巢湖表层沉积物氮、磷、有机质的分布及污染评价[J]. 生态环境学报, 2017, 26(12): 2120-2125. Miao H, Shen Z, Jiang Y, et al. Distribution characteristics and pollution assessment of nitrogen, phosphorus and organic matter in surface sediments of Chaohu Lake[J]. Ecology and Environment Sciences, 2017, 26(12): 2120-2125. |
| [14] |
李如忠, 戴源, 刘晓薇, 等. 巢湖十五里河沉积物硝化速率的城乡梯度变化及相关性[J]. 环境科学学报, 2019, 39(3): 688-695. Li R Z, Dai Y, Liu X W, et al. Gradient variation and correlation of sediment nitrification rates in urban and rural areas of Shiwuli River, Chaohu Lake Basin[J]. Acta Scientiae Circumstantiae, 2019, 39(3): 688-695. |
| [15] |
孔明, 张路, 尹洪斌, 等. 蓝藻暴发对巢湖表层沉积物氮磷及形态分布的影响[J]. 中国环境科学, 2014, 34(5): 1285-1292. Kong M, Zhang L, Yin H B, et al. Influence of algae bloom on distribution of total and speciation of nitrogen and phosphorus in the surface sediments from Lake Chaohu[J]. China Environmental Science, 2014, 34(5): 1285-1292. |
| [16] | Tuttle A K, McMillan S K, Gardner A, et al. Channel complexity and nitrate concentrations drive denitrification rates in urban restored and unrestored streams[J]. Ecological Engineering, 2014, 73: 770-777. DOI:10.1016/j.ecoleng.2014.09.066 |
| [17] | Mulholland P J, Helton A M, Poole G C, et al. Stream denitrification across biomes and its response to anthropogenic nitrate loading[J]. Nature, 2008, 452(7184): 202-205. DOI:10.1038/nature06686 |
| [18] | Herrman K S, Bouchard V, Moore R H. Factors affecting denitrification in agricultural headwater streams in Northeast Ohio, USA[J]. Hydrobiologia, 2008, 598(1): 305-314. DOI:10.1007/s10750-007-9164-4 |
| [19] |
高雪梅, 佘冬立, 颜晓元, 等. 不同量碳源输入梯度下果园排水沟底泥氮素反硝化与N2O排放研究[J]. 环境科学, 2016, 37(7): 2731-2737. Gao X M, She D L, Yan X Y, et al. Denitrification loss and N2O emission from different carbon inputs in orchard drains sediments[J]. Environmental Science, 2016, 37(7): 2731-2737. |
| [20] | Waters E R, Morse J L, Bettez N D, et al. Differential carbon and nitrogen controls of denitrification in riparian zones and streams along an urban to exurban gradient[J]. Journal of Environmental Quality, 2014, 43(3): 955-963. |
| [21] | Malghani S, Kim J, Lee S H, et al. Application of two contrasting rice-residue-based biochars triggered gaseous loss of nitrogen under denitrification-favoring conditions:a short-term study based on acetylene inhibition technique[J]. Applied Soil Ecology, 2018, 127: 112-119. DOI:10.1016/j.apsoil.2018.03.011 |
| [22] |
马培, 李新艳, 王华新, 等. 河流反硝化过程及其在河流氮循环与氮去除中的作用[J]. 农业环境科学学报, 2014, 33(4): 623-633. Ma P, Li X Y, Wang H X, et al. Denitrification and its role in cycling and removal of nitrogen in river[J]. Journal of Agro-Environment Science, 2014, 33(4): 623-633. |
| [23] | Yuan H J, Zhang Z J, Qin S P, et al. Effects of nitrate and water content on acetylene inhibition technique bias when analysing soil denitrification rates under an aerobic atmosphere[J]. Geoderma, 2019, 334: 33-36. DOI:10.1016/j.geoderma.2018.07.039 |
| [24] |
汪银龙, 冯民权, 董向前. 汾河下游水体nirS型反硝化细菌群落组成与无机氮关系[J]. 环境科学, 2019, 40(8): 3596-3603. Wang Y L, Feng M Q, Dong X Q. Community composition of nirS-type denitrifying bacteria in the waters of the lower reaches of the Fenhe river and its relationship with inorganic nitrogen[J]. Environmental Science, 2019, 40(8): 3596-3603. |
| [25] |
邓焕广, 张智博, 张菊, 等. 东鱼河春季沉积物反硝化脱氮作用与N2O排放研究[J]. 水土保持学报, 2019, 33(1): 283-287. Deng H G, Zhang Z B, Zhang J, et al. Denitrification and N2O emission in sediments of Dongyu River in spring[J]. Journal of Soil and Water Conservation, 2019, 33(1): 283-287. |
| [26] | Salahudeen J H, Reshmi R R, Krishnan K A, et al. Denitrification rates in estuarine sediments of Ashtamudi, Kerala, India[J]. Environmental Monitoring and Assessment, 2018, 190(6): 323. DOI:10.1007/s10661-018-6698-z |
| [27] |
李飞跃, 汪建飞, 谢越, 等. 江苏句容水库农业流域农田土壤反硝化作用的研究[J]. 农业环境科学学报, 2012, 31(3): 638-643. Li F Y, Wang J F, Xie Y, et al. Denitrification of farmland soils in the agricultural watershed of Jurong reservoir, Jiangsu province, China[J]. Journal of Agro-Environment Science, 2012, 31(3): 638-643. |