 2020, Vol. 41
2020, Vol. 41

2. 江苏省大气环境监测与污染控制高技术研究重点实验室, 南京 210044;
3. 大气环境与装备技术协同创新中心, 南京 210044;
4. 南京信息工程大学数理与统计学院, 南京 210044;
5. 拉鲁湿地国家级自然保护区管理局, 拉萨 850000
 , TIAN Han-xin1,2,3 , ZHOU Lei1,2,3 , XU De-fu1,2,3
, TIAN Han-xin1,2,3 , ZHOU Lei1,2,3 , XU De-fu1,2,3 , ZHANG Jian-wei4 , PEN CUO Ci-ren5
, ZHANG Jian-wei4 , PEN CUO Ci-ren5
2. Jiangsu Key Laboratory of Atmospheric Environment Monitoring and Pollution Control, Nanjing 210044, China;
3. Collaborative Innovation Center of Atmospheric Environment and Equipment Technology, Nanjing 210044, China;
4. School of Mathematics and Statistics, Nanjing University of Information Science & Technology, Nanjing 210044, China;
5. Lhalu Wetland National Nature Reserve, Lhasa 850000, China
西藏作为中国国土海拔最高的区域, 其土地面积约为1.2亿hm2, 约占国土总面积的八分之一, 其省会拉萨市位于中国西南边陲.西藏既是南亚、东南亚地区的“江河源”与“生态源”, 也是中国乃至东半球气候的“启动与调节区”, 作为青藏高原的主要城市, 西藏所包含的地貌种类较多, 地貌大致可分为喜马拉雅山区, 藏南谷地, 藏北高原和藏东高山峡谷区.湿地被誉为“地球之肾”, 其具有调节生态多样性、改善水质的功能, 地处于西藏拉萨市的拉鲁湿地是整个青藏高原地区生态环境维护重要区域[1~4].拉鲁湿地是世界上海拔最高、我国面积最大的城市天然沼泽地, 其在生态平衡、增加空气湿度、改善拉萨市的城市气候和净化拉萨水环境等方面都起着极其重要的作用[5].
水生植物群落与环境因子之间是相互作用的, 环境因子的变化将影响植物群落的变化.为此, 很多学者都开展环境因素变化对植物群落的影响研究.如赵敏等[6]研究了基于盐分梯度的黑河中游湿地植物多样性及其与土壤因子的关系.王东波等[7]报道了冰封期呼伦湖浮游藻类群落结构及其与水环境因子的关系.刘晓玲等[8]分析了氮磷供应条件对黄河三角洲滨海湿地植物群落结构的影响.在拉鲁湿地植物群落研究方面, 李春[9]认为拉鲁湿地中新的植物群落的入侵和定居已使拉鲁湿地植物群落由早期的单一化向多样化变化发展, 早期以芦苇群落和苔草群落占绝对优势的特征已经不复存在, 拉多等[5]则针对拉鲁湿地植物物种组成与环境因子的关系做了相关研究, 王丽等[10]研究了拉鲁湿地不同水位梯度下植物的多样性特征.上述研究多集中在10年前的研究或通过控制水位梯度研究植物组成的变化, 然而有关拉鲁湿地植物多样性变化及其与水环境因子之间的相互关系研究鲜见报道.为此, 本研究调查了拉鲁湿地水生植物群落和水环境因子, 采用冗余分析(redundancy analysis, RDA)方法, 分析了水生植物多样性指数、丰富度指数和均匀度指数等变化及其与水环境因子之间的关系, 旨在为未来该区域的生态修复与环境污染治理提供理论依据.
1 材料与方法 1.1 研究区域概括拉鲁湿地国家级自然保护区位于西藏自治区首府拉萨市市区西北, 呈东西带状分布.地理坐标介于E91°03′41″~91°06′48″, N29°39′25″~29°42′08″之间.拉鲁湿地北面为高山;东面与拉萨市城关区拉鲁乡居民区接壤;东北面与流沙河相接, 流沙河发源于拉萨市西北部中低山区, 由夺底和娘热两条山沟支流汇合而成, 是一条典型的季节性河流;南面紧邻拉萨市城区, 以拉萨河引水的中干渠和当热路为界, 西面至加油站.中干渠补给水引自献多水电站尾水, 引用流量为55.44m3·s-1.西面则以当热路和与北面高山的交接处为终点.拉鲁湿地被通往西藏自治区党校的鲁定北路分为东西两片湿地, 两片湿地之间通过涵洞相连.
1.2 取样方法拉鲁湿地总体上从东北向西南略为倾斜, 从东至西地面坡度为3‰~5‰, 水从东部流入东部湿地, 然后一部分从鲁定公路下的涵洞流入西部湿地, 并从西部湿地西南角流出, 另一部分从鲁定公路右侧流出湿地.因此, 根据拉鲁湿地的地形特征和水流状况, 将拉鲁湿地分为5个区域, 分别为东部(E)、南部(S)、北部(N)、中西部(M)和西部(W).东部靠近拉萨市城关区拉鲁乡居民区;北部为流沙河补给水流经的湿地区域;南部为中干渠进水流经湿地区域;中西部为鲁定公路两侧, 且由东部流经西部的涵洞区;西部为西部湿地.
2019年5月进行了植物调查, 具体方法为, 沿湿地边缘且有水域地带由西到东布置采样样方.西部设11个样方, 包括采样点1~11号;中西部设6个样方, 包括采样点12~16号和47号;南部设12个样方, 包括采样点36~46号和48号;北部设10个样方, 分别是17~26号采样点;东部设9个样方, 分别是27~35号采样点, 具体的采样点见图 1.样方大小均为1 m×1 m, 调查并记录每个样方的植物的种类、株数、株高、冠幅、盖度和生物量(干重), 然后计算出各植物种类的重要值和物种多样性指数, 同时测定每个样方的水质.

|
图 1 拉鲁湿地采样点分布示意 Fig. 1 Sampling sites in the Lhalu Wetland |
采用便携式多参数水质仪(美国YSI ProDSS)现场监测了拉鲁湿地各采样点的DO、pH、温度、电导率和浊度.每个采样点测3次, 并取其平均值, 代表该样点测量值.采用文献[11]的方法分析水样的总氮、氨氮和总磷.
1.4 数据处理每个样方分别统计其植物种类、所属的科与属, 并统计每个种在样方中的数量.相对密度、相对频度和相对盖度计算方法如下:
(1) 密度和相对密度按下列公式计算[12]

|
(2) 频度和相对频度按下列公式计算[12]

|
(3) 盖度和相对盖度按下列公式计算[12]

|
(4) 物种重要值和多样性指数按下列公式计算[12]
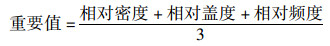
|
Margalef指数(R):
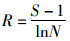
|
Shannon-Wiener多样性指数(H):
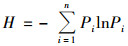
|
Pielou均匀度指数(J):
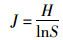
|
Simpson指数(D):
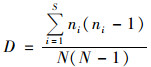
|
式中, S为样方内所有物种数, N为个体总数, Pi是第i种的个体数, ni占总个体数N的比例, 即Pi=ni/N.
1.5 数据分析应用EXCEL 2007对所取样方内植物种类、株数、生物量、株高和盖度等进行统计并计算出相应物种多样性指数, 并采用Origin 9.0绘制水质箱线图.采用SPSS 20分析了拉鲁湿地不同区域水质的变化, 另外, 采用Canoco 4.5对5个区域的植物因子与环境因子进行DCA分析, 并通过选择线性模型冗余分析(RDA)[13]方法, 分析植物多样性与水环境因子间的关系, 且采用蒙特卡洛(Monte Carlo)进行显著性检验[14].
2 结果与分析 2.1 拉鲁湿地不同区域水环境因子特征拉鲁湿地水质变化如图 2所示, 从中可以看出, 对于DO含量, 东部(平均为9.06 mg·L-1)和北部(平均为7.81 mg·L-1)比较高, 中西部(平均为2.46 mg·L-1)比较低, 且东部显著高于中西部(P<0.05).对于电导率, 西部较高(平均为325.46μS·cm-1), 中西部(平均为288.60 μS·cm-1)其次, 南部(平均为266.46 μS·cm-1)、北部(平均为259.32 μS·cm-1)和东部(平均为234.29 μS·cm-1)均比较低, 且西部显著高于东部(P<0.05).对于pH, 北部(平均为8.41)和东部(平均为8.63)较高, 中西部(平均为7.29)较低, 其东部显著高于中西部(P<0.05).对于水温, 西部(平均为18.02℃)较高, 南部(平均为13.11℃)较低;中西部(平均为15.52℃)、北部(平均为16.70℃)和东部(平均为15.85℃)差异性不显著, 但其西部显著高于南部(P<0.05).对于浊度, 东部(平均为3.97 NTU)和北部(平均为3.74 NTU)显著高于中西部(平均为1.24 NTU)(P<0.05), 西部(平均为2.77 NTU)和南部(平均为3.08 NTU)差异性不显著.对于水中氨氮, 西部(平均为0.029 mg·L-1)则较高, 东部(平均为0.017 mg·L-1)则较低, 各个区域之间差异性不大.对于水中总氮, 南部(平均为1.306 mg·L-1)与北部(平均为1.384 mg·L-1)较高, 中西部(平均为0.453 mg·L-1)较低, 南部与北部显著高于中西部和东部(P<0.05);对于水中总磷, 北部(平均为0.052 mg·L-1)较高, 南部(平均为0.015 mg·L-1)较低, 各区域之间差异性不大.

|
不同者小写英文字母表示数据间差异显著(P<0.05);箱体代表上四分位数到下四分位数, 两端须线代表最值, 箱体内部实线代表中位数, 虚线代表平均值 图 2 拉鲁湿地不同区域之间的水因子特征 Fig. 2 Characteristics of water factors in different regions of the Lhalu Wetland |
本次调查, 拉鲁湿地调查区域内有17科17属共18种水生植物, 包含草本17种, 灌木1种.从表 1可以看出, 湿地西部地区据统计共有11种, 其中杉叶藻和水葱为优势种, 其物种重要值由高到低依次为杉叶藻、水葱、菖蒲、空心莲子草、香蒲、灯芯草、芦苇、金鱼藻、萍蓬草、眼子菜和狐尾藻.湿地中西部共有11种, 其中金鱼藻为优势种, 其物种重要值由高到低依次为金鱼藻、水蓼、狐尾藻、菖蒲、水葱、芦苇、空心莲子草、萍蓬草、灯芯草、沿阶草和菹草.湿地南部地区共有6种, 其中水蓼为优势种, 其按物种重要值由高到低依次为水蓼、菖蒲、水葱、芦苇和杉叶藻和水苦荬;湿地北部地区共有8种, 其中水蓼为优势种, 其按物种重要值由高到低依次为水蓼、灯芯草、水葱、香附草、绿萍、眼子菜、酸木和芦苇;湿地东部地区共有7种, 其中水葱为优势种, 其按物种重要值由高到低依次为水葱、灯芯草、菖蒲、香蒲、眼子菜、水蓼和芦苇.
|
|
表 1 拉鲁湿地物种组成与重要值 Table 1 Species composition and important values in the Lhalu Wetland |
2.3 拉鲁湿地不同区域植物多样性
拉鲁湿地不同区域植物丰富度如表 2所示.从中可以看出, 各区域按物种丰富度由大到小排序为(W、M)>N>E>S, Margalef物种丰富度指数依次M>W>E>N>S, 说明湿地中西部的植物种类最多;按Shannon-Wiener指数依次排序为M>W>S>E>N, 说明中西部的植物群落所包含的植物信息量大, 群落的复杂程度较其他地区高;按Simpson指数依次排序为N>E>M>S>W, 说明西部的植物群落中以水葱、杉叶藻为主的优势度最大;按Pielou指数依次排序为S>M>W>E>N, 说明南部植物群落分布最均匀.
|
|
表 2 拉鲁湿地物种多样性指数 Table 2 Species diversity indices in the Lhalu Wetland |
2.4 拉鲁湿地水环境因子与植物多样性之间的关系
本研究的5个区域的4个轴中梯度最大值分别为1.450、0.791、0.868、1.214和0.264, 且均小于3, 符合线性模型, 因此选用RDA方法探究植物多样性与水环境因子间的关系.RDA分析结果如表 3所示, 从中可以看出, 在西部、中西部、南部、北部和东部地区的第一排序轴和第二排序轴分别解释了植物与环境间关系的81.4%和18.3%、96.1%和3.4%、88.3%和10.9%、96.7%和3%及95.2%和4.6%, 表明第一排序轴和第二排序轴均包含了绝大部分排序信息, 为此, 可用前两轴的数据对水环境因子与植物多样性间的关系进行解释.
|
|
表 3 不同区域植物与水环境因子的RDA结果 Table 3 Results of redundancy analysis between plants and water environmental factors in different regions |
植物多样性与水环境因子RDA分析结果表明(表 4), 水环境因子与坐标轴的相关系数越高则水环境因子对植物多样性的相关性越强, 影响越大.如西部第一排序轴反映了浊度的变化, 该因子与第一排序轴的相关系数为0.674 6, 从左到右逐渐增加;第二排序轴反映了水温(0.643 7)的变化, 从上到下逐渐减小.中西部第一排序轴反映了总氮(-0.945 7)的变化, 从左到右逐渐减小, 第二排序轴反映了溶解氧(0.947 4)、电导率(0.751 7)、氨氮(0.671 5)、总磷(0.776 1)和浊度(0.861 2)的变化, 溶解氧、电导率、氨氮、总磷和浊度从上到下均逐渐减小.因此, 在本研究中, 拉鲁湿地不同区域植物多样性受水环境因素的影响.西部为水温和浊度;中部为电导率、溶解氧、浊度、总氮、总磷和氨氮;南部为pH、电导率、水温和溶解氧;北部为溶解氧、电导率、pH、水温、氨氮、总氮和总磷;东部为溶解氧、电导率、pH、水温和浊度、总氮、总磷和氨氮.
|
|
表 4 RDA前两轴与水环境因子之间的相关系数 Table 4 Correlation coefficient between the first two axes of RDA and the water environment factor |
拉鲁湿地不同区域的植物多样性指数与水环境因子的相关性如图 3所示, 从中可以看出, 北部区域溶解氧和pH分别与D呈正相关, 与H、J和R呈负相关(蒙特卡洛检验:P<0.05);该区域水温与H、J和R呈正相关, 与呈D负相关(P<0.05).在拉鲁湿地西部, 浊度与R、H和J呈显著正相关, 与J呈显著负相关(P<0.05).在中西部, 总氮与D呈显著正相关, 与R、H和J呈显著负相关(P<0.05).在东部, 总氮和氨氮分别与D呈弱正相关, 与R、H和J呈弱负相关.在南部, pH和溶解氧分别与J呈弱正相关, 总氮与D呈弱正相关.上述结果说明拉鲁湿地溶解氧、pH、水温、总氮和浊度等是影响植物的多样性的重要因素.

|
红色箭头代表水环境因子, 蓝色箭头代表植物的多样性 图 3 RDA分析排序 Fig. 3 Ordination diagram of RDA |
丰富度指数反映物种的数目.Margalef物种丰富度指数的数值越大, 反映出该群落或生境的物种丰富度越高, 物种数目越多[14, 15].在拉鲁湿地中, 不同区域的Margalef物种丰富度指数依次: M>W>E>N>S, 说明拉鲁湿地中西部区域物种丰富度高, 而南部区域物种丰富度较低.Shannon-Wiener多样性指数是用来描述物种个体出现的紊乱和不确定性的, 指数的值越大, 不确定性越大, 物种分配越平均[15].拉鲁湿地不同区域的Shannon-Wiener指数依次排序为M≈W>S>E>N, 说明中西部和西部区域的植物群落不确定性高, 物种个体分配的均匀程度高, 物种多样性高.Simpson指数又称优势度指数, 反映物种种类出现的概率程度[16].拉鲁湿地不同区域的Simpson指数依次排序为N>E>M>S>W, 说明拉鲁湿地北部区域的植物群落内物种分布的均匀程度最高, 而西部区域植物群落内优势种明显, 优势种的分布集中, 以水葱、杉叶藻为优势种.Pielou指数是物种均匀度的指标, 一般用来表征群落中物种空间分布的均匀程度, 指数的计算值越大, 说明植物分布越均匀[17].拉鲁湿地不同区域的Pielou指数依次排序为S>M>W>E>N, 说明拉鲁湿地南部区域植物群落中的各物种在空间分布的均匀程度最高, 北部区域最低.彭映辉等[18]的研究认为, 水生植物群落多样性的丰富程度与所受干扰强度呈负相关, 而群落多样性的丧失程度与所受干扰强度呈正相关.这说明拉鲁湿地北部、东部和南部区域所受的干扰强度高于西部区域.
3.2 拉鲁湿地植物群落多样性与水环境因子间的关系水分影响了拉鲁湿地植物群落.拉鲁湿地从东北向西南倾斜, 从东至西地面坡度为3‰~5‰, 因此, 拉鲁湿地西部地势较低, 水位较高, 能形成沉水植物(金鱼藻、狐尾藻)-漂浮植物(萍蓬草)和挺水植物(水葱、菖蒲)群落.拉鲁湿地中西部是东部湿地和西部湿地之间的涵洞区域, 水经涵洞从东部湿地进入西部湿地, 涵洞出水口常年积水, 水位较深, 容易形成沉水植物(金鱼藻、狐尾藻、菹草)-漂浮植物(萍蓬草)和挺水植物(菖蒲)群落.拉鲁湿地北部和东部地势较高, 受流沙河季节性进水的影响, 属于间歇性水域, 形成漂浮植物(眼子菜)-挺水植物(灯芯草)群落.在拉鲁湿地南部, 紧靠拉萨市中干渠, 以挺水植物(水蓼、菖蒲)群落为主.因此, 拉鲁湿地北部和东部区域的植物多样性低于西部和中西部区域, 可能与其水分含量较低有关, 这与田应兵等的研究结果相似, 即湿地水分的减少, 植物群落类型发生演替, 水生植物群落逐渐减少[19].
水质影响水生植物群落的变化.吴翠等[20]认为湖水氮磷含量过高, 水体富营养化, 浮游藻类大量繁殖, 造成水生植被覆盖率和生物量都大幅下降, 植物群落结构简单化.郝孟曦等[21]报道水体中TN和TP浓度越高, 水生植物种类和分布越少.从图 1可以看出, 在本次调查中, 拉鲁湿地东部、北部和南部区域未发现沉水植物, 且植物多样性较低, 可能与该区域氮、磷浓度和pH比较高有关.于丹[22]的研究发现水体透明度严重影响沉水植物分布.从图 2和表 1可以看出, 拉鲁湿地中西部和西部沉水植物(金鱼藻、狐尾藻)的生长良好, 可能与该区域水体浊度低, 透明度高有关.金鱼藻、菹草和狐尾藻为耐污的为优势种[5, 23], 大茨藻和小茨藻一般生长在比较洁净的水环境中[23], 水芹和茭白为不耐污植物种[20], 杉叶藻具有良好的耐水淹耐旱能力[17].在本次调查中, 未发现茭白, 而金鱼藻和杉叶藻等优势种, 说明拉鲁湿地植物群落中, 部分耐污种取代非耐污种;植物群落中优势种由耐污种为主, 这在一定程度上说明了水环境的变化, 将引发植被衰退.
4 结论(1) 拉鲁湿地不同区域植物群落变化大, 靠近出水口的西部与中西部区域植物丰富度大且分布不均匀, 以杉叶藻和金鱼藻为优势种的群落;位于入水口的南部与北部区域植物丰富度较低, 南和北部区域以水蓼为优势种, 且植物群落分布不均匀;东部区域为以水葱为优势种, 植物丰富度较小的群落.
(2) 拉鲁湿地各区域之间溶解氧、电导率、pH、水温和浊度的差异性较大, 由西部到东部区域随着海拔高度的升高, 水环境因子的大致变化为溶解氧含量逐渐增大, 电导率逐渐减小, pH逐渐增大, 水温逐渐降低, 浊度逐渐增大.
(3) 溶解氧、pH、水温、总氮和浊度等影响了拉鲁湿地的植物群落, 较高的氮、磷浓度和pH引起植物多样性减少, 呈现耐污性物种逐渐成为拉鲁湿地优势种的趋势.
致谢: 现场调查中得到了拉鲁湿地国家自然保护区管理局梁剑豫和尼琼的支持, 在此一并致谢.
| [1] | Wu J, Lu J, Luo Y M, et al. An overview on the organic pollution around the Qinghai-Tibet plateau:the thought-provoking situation[J]. Environment International, 2016, 97: 264-272. |
| [2] | Yuan G L, Sun Y, Li J, et al. Polychlorinated biphenyls in surface soils of the central Tibetan Plateau:altitudinal and chiral signatures[J]. Environmental Pollution, 2015, 196: 134-140. |
| [3] | Wang Y G, Deng C Y, Liu Y, et al. Identifying change in spatial accumulation of soil salinity in an inland river watershed, China[J]. Science of the Total Environment, 2018, 621: 177-185. |
| [4] | Siles G, Voirin Y, Bénié G B. Open-source based geo-platform to support management of wetlands and biodiversity in Quebec[J]. Ecological Informatics, 2018, 43: 84-95. |
| [5] |
拉多, Birks J, 普布, 等. 拉萨市拉鲁湿地植物群落与环境因素之间的关系及其分布格局[J]. 西藏大学学报(自然科学版), 2009, 24(1): 15-20. Laduo, Birks J, Pubu, et al. The relationship between composition and distribution of vegetation and environmental factors in Lhalu Dhamra, Lhasa[J]. Journal of Tibet University, 2009, 24(1): 15-20. |
| [6] |
赵敏, 赵锐锋, 张丽华, 等. 基于盐分梯度的黑河中游湿地植物多样性及其与土壤因子的关系[J]. 生态学报, 2019, 39(11): 4116-4126. Zhao M, Zhao R F, Zhang L H, et al. Plant diversity and its relationship with soil factors in the middle reaches of the Heihe River based on the soil salinity gradient[J]. Acta Ecologica Sinica, 2019, 39(11): 4116-4126. |
| [7] |
王东波, 君珊, 陈丽, 等. 冰封期呼伦湖浮游藻类群落结构及其与水环境因子的关系[J]. 中国环境监测, 2019, 35(4): 59-66. Wang D B, Jun S, Chen L, et al. Community structures of phytoplankton in the Hulun Lake during icebound season and its relation with environmental factors[J]. Environmental Monitoring in China, 2019, 35(4): 59-66. |
| [8] |
刘晓玲, 王光美, 于君宝, 等. 氮磷供应条件对黄河三角洲滨海湿地植物群落结构的影响[J]. 生态学杂志, 2018, 37(3): 801-809. Liu X L, Wang G M, Yu J B, et al. Effects of nitrogen and phosphorus supply on plant community structure of coastal wetland in the Yellow river Delta[J]. Chinese Journal of Ecology, 2018, 37(3): 801-809. |
| [9] |
李春. 拉鲁湿地生态环境及动植物物种资源变化特征研究[J]. 自然资源学报, 2005, 20(1): 145-151. Li C. Research on the variation characteristics of eco-environment and propagation resources in Lalu Wetland[J]. Journal of Natural Resources, 2005, 20(1): 145-151. |
| [10] |
王丽, 杨涛, 杜红霞, 等. 不同水位梯度下拉鲁湿地典型湿草甸植物群落特征[J]. 植物资源与环境学报, 2018, 27(1): 11-16. Wang L, Yang T, Du H X, et al. Plant community characteristics of typical wet meadow in Lalu wetland under different water level gradients[J]. Journal of Plant Resources and Environment, 2018, 27(1): 11-16. |
| [11] | 国家环境保护总局. 水和废水监测分析方法[M]. (第四版). 北京: 中国环境科学出版社, 2002: 200-284. |
| [12] |
段遍红, 滕玉风, 占玉芳, 等. 黑河桥湿地植物群落组成与多样性研究[J]. 林业科技通讯, 2016(12): 27-30. Duan B H, Teng Y F, Zhan Y F, et al. Research on the plant community composition and diversity of Heihe Bridge wetland[J]. Forest Science and Technology, 2016(12): 27-30. |
| [13] |
侯志勇, 谢永宏, 高大立, 等. 洞庭湖湿地典型植物群落生活型构成及其环境影响因子[J]. 应用与环境生物学报, 2016, 22(6): 993-999. Hou Z Y, Xie Y H, Gao D L, et al. The life form and environment factors of typical plant communities in the Dongting Lake wetlands[J]. Chinese Journal of Applied & Environmental Biology, 2016, 22(6): 993-999. |
| [14] |
吕晓燕, 于鲁冀, 王莉, 等. 淮河流域河南段水生植物多样性评价及其影响因素分析[J]. 中国环境监测, 2015, 31(6): 83-90. Lyu X Y, Yu L J, Wang L, et al. The evaluation of the hydrophytes diversity and its influencing factors of the Huaihe River Basin Henan section[J]. Environmental Monitoring in China, 2015, 31(6): 83-90. |
| [15] |
李振基, 陈圣宾. 群落生态学[M]. 北京: 气象出版社, 2011: 27-32. Li Z J, Chen S B. Community ecology[M]. Beijing: Meteorological Press, 2011: 27-32. |
| [16] |
蔡在峰, 曲文馨, 洪宇薇, 等. 天津市北大港湿地自然保护区植物多样性特征分析[J]. 西北农业学报, 2019, 28(8): 1326-1334. Cai Z F, Qu W X, Hong Y W, et al. Analysis of plant diversity characteristics in Beidagang Wetland Nature Reserve of Tianjin[J]. Acta Agriculturae Boreali-occidentalis Sinica, 2019, 28(8): 1326-1334. |
| [17] |
何彤慧, 程志, 张玉峰, 等. 银川平原沟渠植物多样性特征及影响因素[J]. 湿地科学, 2013, 11(3): 352-358. He T H, Cheng Z, Zhang Y F, et al. Characteristics of plant diversity of canals and ditches and their influencing factors in Yinchuan Plain[J]. Wetland Science, 2013, 11(3): 352-358. |
| [18] |
彭映辉, 简永兴, 王建波, 等. 湖北省五大湖泊水生植物多样性的比较研究[J]. 水生生物学报, 2004, 28(5): 464-470. Peng Y H, Jian Y X, Wang J B, et al. A comparative study on aquatic plant diversity in five largest lakes of Hubei province in China[J]. Acta Hydrobiologica Sinica, 2004, 28(5): 464-470. |
| [19] |
田应兵, 熊明标, 宋光煜. 若尔盖高原湿地土壤的恢复演替及其水分与养分变化[J]. 生态学杂志, 2005, 24(1): 21-25. Tian Y B, Xiong M B, Song G Y. Restoration succession of wetland soils and their changes of water and nutrient in Ruoergai Plateau[J]. Chinese Journal of Ecology, 2005, 24(1): 21-25. |
| [20] |
吴翠, 唐万鹏, 史玉虎, 等. 长湖湿地水生植被演替研究[J]. 湿地科学, 2007, 5(2): 188-191. Wu C, Tang W P, Shi Y H, et al. The succession of aquatic vegetation on the Changhu Lake Wetland, Hubei Province[J]. Wetland Science, 2007, 5(2): 188-191. |
| [21] |
郝孟曦, 杨磊, 孔祥虹, 等. 湖北长湖水生植物多样性及群落演替[J]. 湖泊科学, 2015, 27(1): 94-102. Hao M X, Yang L, Kong X H, et al. Diversity and community succession of macrophytes in Lake Changhu, Hubei Province[J]. Journal of Lake Sciences, 2015, 27(1): 94-102. |
| [22] |
于丹. 水生植物群落动态与演替的研究[J]. 植物生态学报, 1994, 18(4): 372-378. Yu D. Study on the dynamics and succession of aquatic macrophyte communities in the Zhushun Lake, Harbin[J]. Acta Phytoecologica Sinica, 1994, 18(4): 372-378. |
| [23] |
祝国荣, 倪乐意, 方涛, 等. 巢湖东湖区入湖河流的水生植被群落结构和区系特征及其与环境因子的关系[J]. 应用与环境生物学报, 2012, 18(6): 889-896. Zhu G R, Ni L Y, Fang T, et al. Community structure and floristic compositions of aquatic plant communities in the inflowing rivers of East Chaohu Lake and their relationships to environmental factors[J]. Chinese Journal of Applied & Environmental Biology, 2012, 18(6): 889-896. |