 2020, Vol. 41
2020, Vol. 41

2. 西华师范大学环境科学与工程学院, 南充 637002;
3. 杭州市生态环境局淳安分局, 杭州 311700
 , ZHU Guang-wei1
, ZHU Guang-wei1 , LI Yun-xiang2 , WU Zhi-xu3 , ZHENG Wen-ting3 , LAN Jia3 , WANG Yu-cheng3 , XU Hai1 , ZHU Meng-yuan1
, LI Yun-xiang2 , WU Zhi-xu3 , ZHENG Wen-ting3 , LAN Jia3 , WANG Yu-cheng3 , XU Hai1 , ZHU Meng-yuan1
2. College of Environmental Science and Engineering, China West Normal University, Nanchong 637002, China;
3. Hangzhou Bureau of Ecology and Environment Chun'an Branch, Hangzhou 311700, China
水库是我国城市饮用水的重要来源之一, 在社会经济发展中发挥着越来越重要的作用.据水利部2016年发布的《全国重要饮用水水源地名录(2016)》, 全国618个纳入名录的水源地中, 涉及水库的重要水源地269处, 占水源地总数的43.5%[1].然而, 由于水库大多存在换水周期短、流域侵蚀强度大和水位反季节消涨等问题, 水库的水质具有较大的不稳定性, 水环境安全保障面临较大的挑战[2]. 2018年中国环境状况公报指出, 52个国家重要水库中, 6个富营养, 39个中营养, 仅7个为贫营养, 且一些重要城市的供水水库, 如于桥水库、高州水库、山仔水库及汤浦水库等均曾发生过蓝藻水华问题[3~7].水华发生一方面可能会造成取水点的滤水堵塞, 且蓝藻群落产生的毒素、污染和气味问题, 会对供水安全、休闲和旅游业产生重大影响;另一方面, 水华发生可能会导致水生物种组成发生变化, 严重威胁到水生态系统的安全及稳定[8].
水文、气象和水动力等气象水文要素及其动态综合变化对水库藻类水华的形成影响很大, 这使得水库的藻类水华等威胁供水安全的生态灾害的发生具有较大的不确定性.赵孟绪等[7]对汤溪水库蓝藻水华发生条件的研究发现, 营养盐不是汤溪水库浮游植物生长的控制条件, 水体稳定性是控制汤溪水库蓝藻水华发生时间的关键因素.乌拉圭Santa Grande水库的蓝藻及甲藻水华形成受水库入流量、水位、风速及温度等多种气象水文因素组合的综合作用控制[9];加拿大Yamaska河流水库水华的形成与极度干旱有关, 长的停留时间会增强有害藻类的优势度, 并加快藻类群落结构的演替速度[10];刘流等[11]和杨正健等[12]发现表层温跃层变薄是蓝藻水华出现的直接诱因;Yang等[13]发现厦门4个水库中水位陡降等水文条件突变促发了蓝藻优势的形成;吴卫菊等[14]发现水温季节变化是渔洞水库浮游植物结构演替的关键, 而水动力学过程则是浮游植物多样性变化和优势种形成的促发因素.稳定的水文气象条件则会延长水华过程, 如富春江水库和神农溪的蓝藻水华发生后, 稳定水文气象条件均导致了水华长时间地维持[15, 16].
水库的库尾河口区是水库承接大量上游外源污染的区域, 污染负荷较大, 加上水文过程变化大, 使其成为水库水质和水生态最不稳定的区域, 也是水库藻类异常增殖及蓝藻水华发生的敏感点[2].位于新安江水库库尾河口区的街口断面, 承接了60%的流域来水, 同时60%左右的外源颗粒物和营养盐经此入库[17], 水质变化大, 是新安江水库最容易出现富营养化与蓝藻水华的区域[18].近年发展起来的水生态高频监测是捕捉水体水质及藻类快速变化的一种有效手段.本研究以2017年杭州市生态环境局淳安分局设立在街口断面的水质高频监测浮标为基础, 于2017年5月至2018年12月在街口断面实施了3 d一次的高频水样及浮游植物样品的采集, 结合相应地水文气象数据(气温、入库流量及降雨量), 分析了水库入库河口水体的物理指标(温跃层、氧跃层及透明度等)、化学指标(碳、氮及磷等营养盐)及生物指标(浮游植物)的高频变化特征, 并探究了它们对气象水文过程(气温、降雨及入库流量)的响应机制, 以期为水库型水源地水质安全保障提供科学支撑.
1 材料与方法 1.1 研究对象概况新安江水库位于浙江省淳安县和建德市境内(29°22′~29°50′ N, 118°36′~119°14′ E), 是1957年为建设新安江水电站而筑坝形成的大型人工湖泊, 同时也是长三角最大的战略水源地及杭州市的饮用水源.新安江水库地处亚热带季风湿润性气候, 温暖湿润, 雨量充沛, 四季分明, 年均气温为17.8℃, 年均降水量为1 636.5 mm, 年日照时数达1 951 h[19].新安江水库水面面积为583 km2, 总库容1.784×1010 m3, 流域面积10 480 km2, 平均水深31 m, 最大水深100 m, 换水周期约2 a[20].水库水面狭长, 库湾众多, 南北纵长为150 km, 宽处达10 km, 包含1078个面积大于0.25 hm2的岛屿.
街口地处新安江水库库尾, 是新安江水库最大的入湖口[图 1 (a)], 也是新安江水库的水环境监测的国控断面及安徽与浙江的水域交界处.街口断面以上的集雨面积达6 000 km2, 街口处的年均入库水量超过67.77×108 m3, 占总入库径流60%以上[20].自2017年4月起, 杭州市生态环境局淳安分局在街口设置YSI高频监测剖面浮标[图 1 (b)], 每4 h监测一次, 其中水深介于0~10 m之间的上层水体每隔0.5 m测一次, 深度超过10 m后则每隔1 m测一次.浮标测定参数包括水深(WD)、水温(T)、浊度(Turb)、叶绿素a(CHL)、藻蓝素(PC)、电导率(Cond)、溶解氧(DO)及有色溶解性有机质(fDOM)等指标.

|
图 1 新安江水库监测点位及浮标示意 Fig. 1 Location of monitoring site and buoy in Xin'anjiang Reservoir |
2017年5月至2018年12月, 在街口开展了3 d一次的高频水质监测, 采样深度为水下0.5 m.其中2.5 L水样立即加入25 mL鲁戈试剂, 静置48 h后浓缩至30 mL, 用于水体浮游植物群落结构鉴定.另一部分水样立即冷冻, 带回实验室进行水质指标测定, 测定指标包括总氮(TN)、溶解性总氮(DTN)、总磷(TP)、溶解性总磷(DTP)、浮游植物叶绿素a(CHL)及溶解性有机碳(TOC)等, 其中TN与DTN、TP与DTP的区分采用Whatman GF/F玻璃纤维滤膜过滤的方法, 测定方法参照国家地表水环境质量标准[21], 但是测定水体TN和TP时, 未采用静置、分层的预处理, 而是将水样完全摇匀后直接分取25 mL水样, 加入25 mL碱性过硫酸钾消解液进行氧化分解处理;CHL采用热乙醇萃取分光光度法测定[22].透明度(SD)在现场使用塞氏盘测定.浮游植物群落结构数据来源于浮游植物样品的镜检结果, 镜检参考文献[23].
1.3 数据处理为了获得水质剖面分层的季节变化规律, 对YSI浮标的剖面数据进行了分层平均合并.为获得代表性的分层数据, 挑选夏季降雨量及入库流量较小期间的浮标数据, 获得相对稳定水动力状态下水体指标的分层状况, 确定上层、中层和下层的分层深度.对年度变化分析中, 以每日中午12:00的浮标分层数据为基础, 进行全年日变化对比分析.此外, 由于仪器探头漂移问题, 2017年7月5日至2018年2月7日期间的PC值基线偏高, 在数据处理中进行了基线纠偏处理.
浮游植物优势属的高频变化通过相隔两次采样的优势度变化进行刻画.浮游植物优势度(Y)的计算公式为:
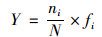
|
式中, ni为两次采样各属的生物量均值, N为两次采样所有属的生物量均值之和, fi为该属的出现频率.其中fi=镜检检出次数/总调查次数, 优势属是指优势度(Y)大于0.02的浮游植物属.
浮游植物群落结构变化与环境因子之间的关系采用RDA分析判别.其中, 浮游植物数据为监测期间出现过的所有优势属的生物量数据, 解释因子为同期的气象数据、水文数据和水质数据, 包括累积气温、累积入库流量、累积降雨、平均水位、平均风速、平均日照时数及氮、磷营养盐(累积值和平均值均为采样间隔期间的数据计算).因千岛湖的气温和水温的相关性很高(R2=0.8), 考虑到共线性问题, 最后RDA分析中采用了对群落结构变化解释率更高的累积气温数据进行分析讨论.采用Hellinger方法对浮游植物数据进行转换, 而环境因子数据通过lg(x+1)进行转换.
降雨量及气温数据来源于国家气象科学数据共享服务平台, 入库流量数据来源于太湖流域片水文信息服务系统中的鱼梁和屯溪两个流量监测站的累加数据[图 1 (a)].
数据处理及图形绘制使用Excel 2010、Origin 8.5、ArcGis 10.2及R 3.5.2等数据分析软件完成.
2 结果与分析 2.1 夏季水动力稳定期各指标的分层状况湖泊和水库的温度分层对水体水质、藻类、浮游动物、水下光环境等水质和生态特征具有重要塑造作用.为确定街口夏季的水质分层状态, 选取2017年7月27日、2017年8月8日、2018年7月29日及2018年8月8日中午12:00的测定数据作图(图 2).不同的水环境参数在垂向分层上有所差异.水温在0~4 m基本无较大变化, 在4~13 m温度大幅降低.溶解氧在水下0~5 m一直处于降低趋势, 在5~10 m出现最低值, 当水深超过10 m后, 溶解氧就不再变化.浊度表现为表层和底层高, 中间低, 且在水深为0~5 m时基本呈线性降低趋势, 在5~10 m浊度基本不变, 当水深超过12 m, 浊度又呈小幅增加趋势.叶绿素在0~5 m值最高, 在5~10 m有小幅度的变化, 当水深超过10 m时, 叶绿素基本为零.fDOM的变化幅度较大, 但在0~5 m呈下降趋势, 在5~10 m波动较大, 且2017年得波动幅度大于2018年, 并在5~10 m处出现极低值.fDOM在水深10~17 m基本无变化, 当水深超过18 m时, 又开始降低.

|
QSU为浮标自动监测系统的动态光学单位 图 2 街口监测断面夏季水质参数的分层特征 Fig. 2 Stratification of environmental water parameters in summer obtained at Jiekou monitoring site |
综合上述指标变化, 笔者将0~5 m定义为层Ⅰ, 5~10 m定义为层Ⅱ、10~20 m则定义为层Ⅲ.在进行年度变化分析时, 将各层的YSI测定指标进行数学平均, 然后按照3层进行趋势分析, 以剔除部分异常波动带来的影响, 获得相对稳定的水质指标随时间变化分层变化特征.
2.2 水质及气象水文参数的周年高频变化特征2017年5月至2018年12月的降雨量、气温、入库断面流量和街口YSI浮标监测各参数逐日变化及3 d一次的表层水体TN、DTN、TP、DTP、CHL浓度和SD深度等变化如图 3所示.

|
红线为参考线 图 3 2017~2018年街口监测断面主要水质指标的高频变化 Fig. 3 Variations of water environment indicators at Jiekou monitoring site from 2017 to 2018 |
从图 3可知, 日均气温在-0.8~33.8℃之间波动, 平均值为(18.2±8.6)℃, 其中超过25℃的天数占27%, 15~25℃的天数占35%, 低于15℃的天数占38%. 6~9月为气温高值期(5月中旬气温开始超过25℃, 9月下旬至10月上旬气温开始低于25℃), 12月~次年2月则是气温的低值期, 最高值出现在2017年7月28日, 最低值出现在2018年1月27日.日降雨量在0.0~83.3 mm之间波动, 最大值出现在2017年6月13日, 平均值为(4.0±9.4)mm, 2017年降雨量总计为1510.7 mm, 2018年为1385.4 mm.日入库断面流量(渔梁水文站与屯溪水文站之和)在7.0×105~2.6×108 m3·d-1之间波动, 最小值出现在2017年11月7日, 最大值出现在2017年6月24日, 平均值为(1.0×107±1.9×107)m3·d-1.入库流量的峰值与降雨量峰值一致, 但是, 其小峰变化明显少于降雨量(图 3), 这表明小雨和中雨不一定会形成入库流.此外, 街口断面上游45 km处的水坝对小、中等规模的降雨形成的入流具有一定的拦截作用. 2018年3月初的第一个流量小峰值对水体透明度及营养盐浓度造成了巨大的改变, 这表明每年的第一个较大入库流量对街口水环境的影响巨大.
水体浊度总体呈现表、中、底3层逐渐升高的特征, 且随入库流量的增大而增大, 呈现脉冲式变化的特点(图 3). 0~5 m浊度在0.24~111.12 NTU之间, 平均值为(5.12±8.30)NTU, 5~10 m浊度在0.01~104.39 NTU之间, 平均值为(5.66±10.02)NTU, 10~20 m浊度在0.01~210.14 NTU之间, 平均值为(6.65±12.78)NTU.浊度的时间变化趋势基本与入库流量一致, 在4月开始出现较大峰值, 最大峰值出现在6月底或7月初.水体SD受汛期及浮游植物季节性增殖影响明显, 日均值介于12~465 cm之间, 平均值为(214±96)cm, 3~9月的透明度明显低于10月~次年2月.
水温分层出现在3月中旬至10月中旬, 3层水体的平均温度随水深增加而降低, 且分层受持续降雨和暴雨的影响明显, 暴雨期水温分层消失, 且总体水温明显下降(图 3). 0~5 m水温在9.82~32.93℃之间, 平均值为(22.41±7.62)℃. 5~10 m的水温在9.68~30.49℃之间, 均值为(21.15±6.13)℃. 10~20 m的水温在9.30~25.3℃之间, 均值为(18.24±4.52)℃.此外, 水温分层的起始温度明显低于结束温度, 水温在14℃左右开始分层, 低于24℃开始分层消失.分层开始与消失的温度差异巨大, 主要与水体的下层温度不同有关.春季水温分层形成时, 整个水柱温度都比较低, 表层水温稍有增高, 就可能导致温度分层;而秋季水温分层消失时, 整个水体的温度处于较高水平, 下层水温较高, 此时, 表层水温在较高的情况下, 已经与底层水温没有明显差别了, 所以水温分层消失时的温度较高.
水体DO的分层情况与水温分层基本同步, 自3月中旬开始, 10月中旬结束.但溶解氧的波动较温度更大. 0~5 m溶解氧浓度在5.03~14.69 mg·L-1之间, 平均值为(8.72±1.94)mg·L-1, 5~10 m溶解氧浓度在1.36~11.76 mg·L-1之间, 平均值为(7.01±1.77)mg·L-1, 10~20 m溶解氧浓度在0.30~11.82 mg·L-1之间, 平均值为(5.99±2.76)mg·L-1.氧分层强度随温度升高而增大, 其中8月中旬~9月中下旬, 底层水体溶解氧会出现持续的缺氧状态(图 3).此外, 暴雨入流过程会破坏水体溶解氧分层.
水体浮游植物叶绿素a的分层期与水温分层期大致相同, 但是分层状况较水温分层更加不稳定, 分层与混合频繁交替(图 3).水体叶绿素a浓度变幅很大.其中, 0~5 m叶绿素在0.95~57.07 μg·L-1之间, 平均值为(6.55±6.02)μg·L-1, 5~10 m叶绿素在0.63~30.40 μg·L-1之间, 平均值为(2.57±2.53)μg·L-1, 10~20 m叶绿素在0.33~27.87 μg·L-1之间, 平均值为(1.72±2.49)μg·L-1. 0~5 m的叶绿素显著高于5~10 m及10~20 m, 平均值是10~20 m均值的3.8倍.叶绿素峰值一般出现在温度较高且降雨及入库流量均低的时期. 0~5 m藻蓝素在0.01~2.87 μg·L-1之间, 平均值为(0.33±0.40)μg·L-1, 5~10 m藻蓝素在0.01~2.64 μg·L-1之间, 平均值为(0.13±0.18)μg·L-1, 10~20 m藻蓝素在0.00~2.64 μg·L-1之间, 平均值为(0.15±0.19)μg·L-1. 2018年的藻蓝素显著高于2017年的同期水平, 这与浮游植物镜检所发现的2018年新安江水库蓝藻比例增高现象吻合.
水体总磷浓度受到降雨入流强度的影响, 但是变化强度与降雨强度变化不完全一致, 主要表现在波动的滞后性明显拉长, 峰谷切换平缓(图 3).TP浓度总体上介于0.011~0.188 mg·L-1之间波动, 平均值为(0.055±0.030)mg·L-1.DTP浓度介于0.003~0.122 mg·L-1之间波动, 平均值为(0.031±0.025)mg·L-1, DTP平均占TP的56%, 溶解态比例超过一半.高频监测表明水文过程驱动下水体TP短期超标是一种自然现象, 在进行国控断面的水质达标评估中, 应当考虑监测期间的水文过程背景.
水体总氮的时间变化与TP基本同步, 但是波动更小(图 3).TN浓度总体在0.75~2.76 mg·L-1之间波动, 平均值为(1.29±0.39)mg·L-1.DTN浓度在0.58~2.69 mg·L-1之间波动, 平均值为(1.14±0.37)mg·L-1.总氮中溶解态比例很高, DTN平均占TN的88%, 因此总氮浓度受水文冲击带来的浊度变化、高温晴热带来的藻类生物量变化影响相对较小, 因此, 总氮波动性比总磷小.
反映水体溶解性有机质浓度高低的fDOM指标也呈现明显的波动, 变化速度较叶绿素a的变化更快(图 3). 0~5 m的fDOM在1.38~21.77 QSU之间, 平均值为(8.91±3.25)QSU, 5~10 m的fDOM在1.44~20.63 QSU之间, 平均值为(7.65±3.29)QSU, 10~20 m的fDOM在1.01~19.42 QSU之间, 平均值为(8.28±2.47)QSU. 3~10月中旬是fDOM的分层期, 表层fDOM在7~9月会降至最低. 11月~次年2月底, 呈现混合均匀的特点.
总体来看, 营养盐的变化对降雨过程的响应尽管同步性不好, 但讯期(3~6月)的碳、氮、磷浓度是非汛期(其它月份)均值的2倍(P < 0.001).讯期与非汛期TN的浓度均值分别为(1.67±0.32)mg·L-1和(1.03±0.15)mg·L-1, DTN分别为(1.47±0.33)mg·L-1和(0.91±0.15)mg·L-1, TP分别为(0.080±0.029)mg·L-1和(0.038±0.015)mg·L-1, DTP分别为(0.046±0.032)mg·L-1和(0.021±0.011)mg·L-1, fDOM分别为(3.71±0.22)QSU和(1.64±0.09)QSU.这说明, 尽管不同营养盐对降雨过程的响应滞后程度不同, 但是降雨入流带来的综合影响是相似的.
2.3 浮游植物群落结构的高频变化特征 2.3.1 各门生物量占比及生物量变化高频监测结果表明, 街口的浮游植物主要为硅藻、隐藻、绿藻、蓝藻、甲藻、裸藻及金藻, 其中硅藻、隐藻、绿藻及蓝藻为主要门类.整个观测周期各门生物量平均占比分别为48%、15%、22%及12%[图 4 (d)].街口全年浮游植物以硅藻为主, 隐藻主要出现春季, 蓝藻及绿藻主要出现在夏季[图 4 (e)].

|
虚线为参考线 图 4 街口监测断面浮游植物变化特征 Fig. 4 High-frequency variation trends of phytoplankton at Jiekou monitoring site |
从生物量占比可知, 一年中的大部分时间均存在蓝藻, 监测期间共出现4次明显峰值过程, 分别为2017年8月14日至9月15日、2017年9月29日至10月30日、2018年2月2日至3月5日及2018年7月25日至10月26日[图 4 (d)].这4次过程, 蓝藻占比均超过了10%.但是, 不同年份间蓝藻峰值的出现时间大有不同. 2018年蓝藻夏季占比峰值较2017年提前26 d.此外, 在2018年2月底还出现一个蓝藻生物量占比小峰值, 而2017年由于该段时间还未开展监测, 并不清楚是否也存在冬季峰值.硅藻占比波动变化很大, 在0% ~100%之间, 但其在大多时间的占比均较高.与2017年相比, 2018年5~11月的平均占比下降8%.绿藻的阶段性生物量占比较大, 且主要出现在蓝藻生物量占优的前期. 2018年绿藻在6月1日的占比达到30%, 而2017年7月17日绿藻占比才首次突破30%. 2018年绿藻峰值出现的时间较2017年提前, 但5~11月的绿藻平均占比下降8%.隐藻占比峰值出现在4月, 占比最高值出现在2018年4月20日.除8、9月没有隐藻, 其它月份均有隐藻. 2018年5~11月的平均隐藻占比较2017年升高15%.隐藻占比峰值大致与蓝藻占比峰值相反, 蓝藻占比高的时候, 隐藻占比较低.甲藻门、裸藻门和金藻门的占比较低, 其中, 甲藻门主要在1~7月出现.
就总生物量而言, 浮游植物总生物量介于2.6×10-3~12.2 mg·L-1之间, 其中3~9的生物量显著高于其它月份(P < 0.001), 最高值出现在2017年8月25日[图 4 (e)].总生物量存在一定年际差异, 2017年的峰值出现在8月15日, 2018年的峰值出现在7月31日, 且2018年的峰值比2017年低32%[图 4 (e)].各门生物量的高频变化可知.研究期间, 硅藻生物量在0~10.0 mg·L-1之间波动, 除了7~9月的部分时间硅藻被蓝藻和绿藻取代优势地位之外, 大多数时间, 硅藻生物量都是所有门中生物量最大的, 主导了整个浮游植物生物量的波动. 2018年硅藻的生物量明显低于2017年.蓝藻门与硅藻门的生物量变化具有一定的同步性, 均表现出夏秋季节升高的特征.蓝藻生物量在0~2.62 mg·L-1之间, 最大值出现在2018年8月21日, 且2018年2月3日至4月2日蓝藻生物量有一段小峰值. 2018年的蓝藻较2017年出现时间较早且生物量较高, 2018年蓝藻在6月13日便持续出现, 比2017年提前27 d, 且2018年的峰值为2.62 mg·L-1, 比2017年高0.54 mg·L-1.此外, 绿藻主要出现在7月中旬~9月初, 呈现双峰特征, 且绿藻峰值及其出现时间存在较大年际差异. 2018年的生物量第一个小峰值出现在6月22日, 为1.34 mg·L-1, 最高峰值出现在7月28日, 为3.77 mg·L-1;与2017年相比, 绿藻小峰值出现的时间提前22 d, 但大峰值远低于2017年.隐藻的生物量峰值主要出现在3~6月期间, 生物量最高值出现在2018年4月2日, 为2.11 mg·L-1.
总的来讲, 高频监测抓住了各门浮游植物的长期演替规律及短期波动的变化特征, 全年基本呈现为硅藻/绿藻→硅藻/隐藻→绿藻/蓝藻→硅藻/绿藻的演替模式.降雨过程及入库流量过程对藻类影响较大, 藻类生物量峰值均出现在流量及降雨量较小, 气温较高的情况下, 且暴雨及大的入库流量会造成浮游植物的生物量迅速大幅下降(图 4).
2.3.2 优势属的高频变化高频监测表明, 河口区水体浮游植物种属结构多样, 且呈高度动态变化的特征. 2017年5月3日至2018年12月1日先后出现的主要优势属有24个, 包括蓝藻门的5个属, 分别为颤藻属(Oscillatoria spp.)、微囊藻属(Microcystis spp.)、鱼腥藻属(Dolichospermun spp.)、席藻属(Phormidium spp.)和束丝藻属(Aphanizomenon spp.), 绿藻门的5个属, 分别为纤维藻属(Ankistrodesmus spp.)、角星鼓藻属(Staurastrum spp.)、栅藻属(Scenedesmus spp.)、盘星藻属(Pediastrum spp.)和新月藻属(Closterium spp.), 硅藻门的9个属, 分别为舟形藻属(Navicula spp.)、脆杆藻属(Fragilaria spp.)、星杆藻属(Asterionella spp.)、异极藻属(Gomphonema spp.)、曲壳藻属(Achnanthes spp.)、桥弯藻属(Cymbella spp.)、小环藻属(Cyclotella spp.)、针杆藻属(Synedra spp.)和直链藻属(Melosira spp.), 隐藻门的2个属, 分别为蓝隐藻属(Chroomonas spp.)和隐藻属(Cryptomonas spp.), 甲藻门2个属, 分别为多甲藻属(Peridinium spp.)和角甲藻属(Ceratium spp.), 以及裸藻门的1个属, 为裸藻属(Euglena spp.).各优势属快速演替, 少则持续3 d至1周, 而多则持续半年, 呈现高度的动态变化(图 5).

|
“*”表示其为优势属 图 5 街口监测断面浮游植物优势属的高频变化 Fig. 5 High-frequency variations of phytoplankton dominant genus at Jiekou monitoring site |
蓝藻门的束丝藻适宜生长的季节比较宽泛, 在2017年7月初~11月中旬、2018年的2~3月和6~12月均表现出较高的优势度.而适宜蓝藻门中的微囊藻和颤藻生长的季节相对更短, 主要出现在温度较高的时期(2017年7月下旬~8月和2018年8~9月), 且具有较高的生物量.鱼腥藻的生长条件比较宽泛, 但其优势度不大.蓝藻门的优势属演替模式大致可分为5个阶段:1~3月为鱼腥藻属到束丝藻属的转变, 4~5月蓝藻门无优势属, 6月束丝藻属又开始持续占优, 8~9月微囊藻属和颤藻属生物量增加, 与束丝藻属共同占优, 9月之后束丝藻属仍占优, 但生物量降低.
绿藻门最主要的优势属为盘星藻属, 2017年7~9月、2018年的6月和8~9月, 均呈现明显的优势度.新月藻则与之互补, 主要在冬季等温度较低的季节出现.绿藻门的优势属演替主要分为两个阶段:6~9月盘星藻属占优, 10月~次年2月新月藻属占优.
硅藻门中最大量出现的是针杆藻和直链藻, 除了8月及2018年的11~12月, 其余月份大多都是优势属.此外, 小环藻则在温度较低的季节成为优势属.脆杆藻则是在秋季成为优势.硅藻门的优势属演替主要分为3~7月直链藻属和针杆藻属占优, 7~10月脆杆藻属和针杆藻属占优, 11月~次年2月小环藻属、针杆藻属及直链藻属共同占优.
隐藻门中的主要优势属是隐藻属, 2018年3~6月呈现了较高的优势度.甲藻门的多甲藻属在2018年3~5月出现连续占优的情况, 而角多甲属在监测期间也出现占优情况, 但存在明显的偶发性.金藻门在监测期间未检出优势属, 而裸藻门裸藻属占优为偶发性.
3 讨论 3.1 水库河口区的热分层与氧分层特征热分层是湖泊和水库中极其重要的物理过程[24, 25].热分层将导致水体上下物质交换大大受阻, 进而对水体化学指标和生物指标产生关键性的影响.热分层的形成与气温关系密切, 但水动力、水体透明度及透明度变化伴随的光能收支也会对热分层造成影响[26].相关研究表明, 在水动力相对稳定的坝前区域, 千岛湖的热分层主要受气温和透明度的影响, 热分层一旦形成, 相当稳定, 直至秋季稳定下降到一定水平才消失[27~29].与新安江水库之前坝前水域的研究结果不同, 本次高频监测发现水库河口区的温度分层有明显区别:热分层消失季节偏早, 且温度分层频繁受到降雨入流破坏.从图 3可以看出, 街口的热分层从3月中旬水温达到14℃左右开始形成, 一般到10月中旬水温24℃以后逐渐消失, 但是分层期间3层水体多次发生不同程度的混合.比较大的混合发生在2017年的4月2日、4月10日、6月14日和6月25日, 以及2018年的4月13日、4月25日、5月28日和7月2日, 其中2017年4次混合所对应的前5 d累积降雨量分别为21.2、111.7、207.1和187.5 mm, 而2018年4次混合所对应的前5 d累积降雨量分别为43.0、65.3、37.8和84.8 mm.可见, 超过20 mm以上的大雨事件即能对河口区的热分层产生明显影响.这与文献[28, 29]监测的坝前水域温跃层稳定存在的情况不同.据Liu等[28]的坝前高频监测数据分析, 该水域热分层从3月初表层水温达到14℃左右开始形成, 直到11月中旬水温下降到20℃左右才开始消失, 期间温跃层十分稳定.这表明河口区的温跃层稳定性明显弱于坝前区.热分层的频繁变化导致氧分层和营养盐上下交换等水质的相应变化, 为浮游植物群落结构变化和藻类优势属演替带来了契机.
氧分层的形成对水质产生重要的影响, 底层缺氧会促进可溶性活性磷的内源性释放, 是水库营养盐的重要补给途径[30].此外, 底层缺氧会导致水体下层及表层底泥厌氧分解, 产生危害水质的异味物质, 是湖库水源地水质安全监控的重要过程.已有研究表明, 水库的氧分层主要受热分层变化的影响[31, 32], 但同时还受水体初级生产力变化的影响, 以及表层水体的水动力复氧过程的影响[33].因此, 氧分层较热分层变化更为剧烈.据Zhang等[32]前期对新安江水库主要湖泊区的氧分层研究表明, 新安江水库氧跃层厚度与温跃层厚度显著正相关(R2=0.99, P < 0.001).但是从图 3可见, 河口区的氧分层变化明显多于热分层, 除了降雨入流等水动力扰动外, 水动力稳定期适宜条件下的表层浮游植物生长产氧过程、下层微生物耗氧过程等对水体氧分层同样产生影响.
图 6显示了气温、入流流量及降雨量与水库河口区不同层溶解氧误差值(CVDO)和水温偏差值(CVWT)的关系.从中可以得出如下定量关系:气温高于14℃时, 河口区氧分层逐步形成;温跃层则是随着气温的升高, 分层概率逐步加大.降雨入流的扰动与气温的改变共同控制了河口区的氧跃层和温跃层, 温度与3层水体溶解氧及温度的变异系数均表现为显著正相关关系(P < 0.001), R2分别达到0.58及0.54(图 6);随着降雨量增大, 3层水体的水温及溶解氧的变异系数明显降低, 且4~9月的降雨量与溶氧分层及温度分层存在显著负相关关系, 相关系数分别为-0.19(P < 0.001)和-0.13(P < 0.050), 降雨量越大, 水相混合度越高, 当日降雨量超过60 mm, 热分层、氧分层基本消失;入库流量增加, 水相混合作用加强, 入库流量与溶氧及水温的变异系数表现为极显著负相关(P < 0.001), R2分别为0.51及0.25, 热分层和氧分层迅速减小, 当入库流量大于5×107 m3·d-1, 热分层和氧分层基本消失.

|
CVDO代表溶解氧变异系数; CVWT代表水温变异系数 图 6 温跃层、氧跃层与气温、入库流量及降雨量关系 Fig. 6 Relationship between the thermocline, oxygen layer, temperature, inflow and rainfall |
高频观测更加直观地揭示了气象水文过程对水库河口区营养盐的决定性影响.前期的研究表明, 新安江水库水营养盐浓度在季节上受降雨量的影响[34].然而, 前期基于月尺度的雨量与营养盐关系并不好, 比如兰佳等[35]基于2008~2009年的监测数据认为新安江水库的TN、TP浓度与上月降雨量之间无显著相关关系.盛海燕等[36]基于2003~2012年的调查数据发现新安江水库逐月降雨量及入库流量与下月水体中氮、磷含量不存在显著关系.笪文怡等[37]分析了2016~2017年的氮、磷浓度与不同时间尺度的累积降雨量关系, 结果发现降雨量仅与磷营养盐浓度存在显著关系.这些研究缺乏高频的监测, 加上河口区多变的水环境与浮游植物群落结构, 导致二者的关系不好.基于本调查, 可以发现采样前3 d累积降雨量、累积入库流量与营养盐指标之间极显著正相关(表 1, P < 0.001), 其中, 入库流量与碳、氮和磷的相关性要明显大于降雨量.然而, 气温与营养盐的关系较差, 仅TP与气温显著相关.这表明水库河口区的营养盐主要受外源入库影响, 是一个物理作用为主的过程.由于TP中有相当大比例是以颗粒态存在, 特别是藻类生物体中含有大量的磷, 因此, 气温与TP之间有较好的相关性, 而与DTP相关性差.
|
|
表 1 营养盐与前三日累积水文气象条件的皮尔逊相关性分析1) Table 1 Pearson's correlation analysis among hydro-meteorological factors and nutrients |
降雨强度与河口区营养盐浓度之间的关系并非简单的线性关系, 如2018年3月2日以来的降雨形成的较大入库流量直接造成营养盐浓度增加1倍(图 2).不同季节的降雨与营养盐的关系不同, 面源污染的输出强度随降雨过程会快速下降.汛期(3~6月)和非汛期相比, 采样前3 d的累积降雨量及累积入库流量与营养盐浓度的关系存在显著差异(图 7).

|
图 7 不同时期的营养盐与气象水文关系 Fig. 7 Relationship among nutrients and hydro-meteorological factors in different periods |
降雨强度方面, 当前3 d的累积降雨量小于10 mm时, 降雨主要起到稀释的作用, 水中的TN和TP浓度均呈小幅降低趋势.前3 d的累积降雨量在10~20 mm的时候, 汛期水中的TN和TP浓度均呈小幅上升趋势, 非汛期营养盐浓度变化不大.前3 d的累积降雨量超过50 mm之后, 汛期的TN浓度在出现暴雨的情况下, 浓度会增加.其原因是因为暴雨对地表的冲击能力大, 可将土壤中的氮洗脱到水体中[38, 39].罗献宝等研究表明洪灾会对新安江水库的水质造成巨大的影响[40]. 3~6月降雨量较大季节, 当前3 d的累积入库流量小于1.0×107 m3时, TN、TP及fDOM的浓度随降雨量增加呈显著增加趋势, 而当前3 d的累积入库流量超过1.0×108 m3时, TP浓度随入库流量增加还有所降低, 但TN浓度在累积入库流量超过3.0×108 m3后显著增高.当前3 d的累积入库流量超过2.5×108 m3后, 水体中fDOM浓度明显降低, 呈现明显的稀释作用.
3.3 水库河口区浮游植物群落结构变化及影响因素中营养水库的浮游植物生长, 既受气温、降雨和水位等气象水文条件的控制, 又受氮、磷和硅等营养盐供给的影响[41].一般认为, 浮游植物的生长与组成的最关键因子为营养盐浓度, 异常增殖主要归因于高营养盐浓度, 特别是磷的浓度.然而越来愈多的研究表明, 物理因素在控制浮游植物的动态变化中也发挥着巨大的作用.已有研究表明河流流量、水力滞留时间和死水区的存在, 对浮游植物生物量具有重大影响[42], 且水华的发生是由多种非生物组合共同控制, 如流量和水温[43], 流量、温度和养分浓度[44], 流量、温度和辐射[45].在环境因子、水文条件多变的水库河口区, 成为多种浮游植物优势属竞争的热点区域, 也必将成为整个水库浮游植物群落结构突变的策源地.Bowes等[8]的相关研究也指出, 要了解控制藻华动力学的因素, 就需要在高频时间尺度上获取物理、化学及生物监测据.因此对于河口区这种水动力扰动强烈的不稳定生态系统而言, 高频监测应该能够得出更加准确的因果关系.
根据浮游植物在监测期间是否占优, 共选取了24属浮游植物进行RDA分析, 这些浮游植物的生物量占整个水库浮游植物的99.6%.对本次高频监测数据中浮游植物优势属与环境因子的关系分析表明, 新安江水库河口区浮游植物群落结构变化有密切联系的环境因子包括累积气温、平均水位、累积入库流量、平均日照时数、DOC、TN、TP、TN :TP和SD(图 8), 这些环境因子共解释了42.45%浮游植物的变化.

|
图中标注的环境因子均为浮游植物群落变化的显著影响因子(P < 0.050) 图 8 街口监测断面水体浮游植物优势属与环境因子RDA分析 Fig. 8 RDA analysis of phytoplankton community and environmental factors at Jiekou monitoring site |
不同优势属生长的关键影响因子不同.全年基础上RDA分析表明, 蓝藻门的微囊藻属及颤藻属与累积气温、平均水位、DOC等因子正相关, 但蓝藻门所有属均与TN、TP、累积入库流量等因子负相关(图 8).这说明对千岛湖的蓝藻而言, 高温、高水位等物理条件对其生长、占优起决定性作用, 但暴雨过程不利于其生物量累积.此外, 该区域的营养盐浓度高低对蓝藻的生长及其占优不产生主要影响.硅藻不同优势属与环境因子的关系则呈现高度的多样性, 表明硅藻不同属生物学特征差异大, 分别适合不同季节、水文情势生长, 呈现较大的多样性, 这也是硅藻门浮游植物在大多数月份能够保持生物量占优的原因.绿藻门的新月藻属生物量与透明度显著相关, 而与入库流量和积温等负相关, 表现出对静水、冬季等条件的适应性, 这与相关文献揭示的特征一致.王轲等[46]的实验研究也提出新月藻个体较小, 低浓度氨氮即可满足其生长所需的氮源, Vuorio等[47]的研究同样表明低营养盐水平的水域更有利于粒径小的藻细胞生长.
不同季节、不同优势属与环境因子的关系不同(图 8), 这与不同季节环境因子对藻类生长的限制强度变化有关.春季至冬季解释率依次为30.35%、45.43%、44.54%及51.5%.春季的影响因子包括累积入库流量、平均水位和TP, 其中累积入库流量是最主要的影响因子, 体现了春季的水文条件对藻类优势属形成的决定作用, 如2018年春季的第一个较大入库流量, 直接破坏了浮游植物的群落结构, 大幅降低了蓝藻占比(图 4).此外, 春季一般营养盐充足, 对藻类生长的限制有限;夏季的影响因子包括累积入库流量、平均日照时数、平均水位、SD、DOC、TN和TP;秋季的影响因子为累积入库流量、累积气温、平均日照时数、平均水位、TN、DOC和SD, 其中累积气温、SD及平均水位为最主要的因子;冬季为累积气温、平均水位和DOC(图 8).这种季节性的主导因子差异, 一则是不同季节适合生长的藻类优势属不同, 二则是由于不同季节环境条件自身的变化所致.从高频浮游植物数据与环境因子的RDA分析中, 可以看出, 除了夏季TN是浮游植物群落结构变化的最重要因素, 其它季节最重要的影响因子均为气象水文因子, 且夏季的水文因子对群落结构的影响也非常巨大.可见, 对于千岛湖街口而言, 气象水文条件是浮游植物生长的绝对控制因素.此外, 从藻类异常增殖的敏感季节看, 夏季的营养盐均为影响藻类群落结构演替的关键因素, 这表明新安江水库的藻类异常增殖已经受到氮、磷的影响.在流域面源污染控制方面, 应当增大力度.
4 结论(1) 通过高频监测表明, 大型深水水库的库尾河口区水体热分层、氧分层、营养盐浓度和藻类生物量及优势属变化频繁, 降雨及其伴随的入流扰动是水库河口区水环境及浮游植物群落结构高度动态变化的重要驱动力.
(2) 春季气温达到14℃以上时, 新安江水库河口区热分层和氧分层开始形成, 秋季气温降至24℃时, 热分层和氧分层开始消失, 分层期间, 降雨量大于20 mm时, 热分层开始受到明显冲击, 大于80 mm时热分层基本全部破坏, 营养盐浓度受降雨量及入库流量的影响显著, 总磷还与气温显著正相关.
(3) 新安江水库河口区浮游植物群落结构变化多样, 其中硅藻、蓝藻、绿藻及隐藻是主要类群, 年均生物量占总浮游植物生物量的97%.优势属演替受降雨、入库流量等影响大, 夏季还受碳、氮和磷等营养盐的影响.监控与预警蓝藻等藻类异常增殖时间, 水文气象及营养盐是重要考量参数.
致谢: 感谢杭州市生态环境局淳安分局在样品采集及高频监测浮标数据支撑上的大力帮助, 感谢安徽师范大学硕士研究生孙祥和史鹏程等在采样过程中给予的帮助, 感谢陈晓霞对浮游植物样品的鉴定, 感谢中国科学院南京地理与湖泊研究所邹伟和国超旋博士研究生在数据分析方法上的协助.
| [1] | 李达.全国重要饮用水水源地名录(2016年)[EB/OL]. https://wenku.baidu.com/view/1c253560cec789eb172ded630b1c59eef8c79a21.html, 2018-08-11. |
| [2] |
王丽婧, 郑丙辉. 水库生态安全评估方法(Ⅰ):IROW框架[J]. 湖泊科学, 2010, 22(2): 169-175. Wang L J, Zheng B H. Method for ecological security assessment of reservoirs (Ⅰ):IROW framework[J]. Journal of Lake Sciences, 2010, 22(2): 169-175. |
| [3] |
房旭, 段洪涛, 曹志刚, 等. 基于多源卫星数据的小型水体蓝藻水华联合监测——以天津于桥水库为例[J]. 湖泊科学, 2018, 30(4): 967-978. Fang X, Duan H T, Cao Z G, et al. Remote monitoring of cyanobacterial blooms using multi-source satellite data:a case of Yuqiao Reservoir, Tianjin[J]. Journal of Lake Sciences, 2018, 30(4): 967-978. |
| [4] |
王菲凤, 仝川, 杨芳, 等. 福州山仔水库水华微囊藻毒素时空分布特征[J]. 环境科学学报, 2011, 31(3): 533-546. Wang F F, Tong C, Yang F, et al. Spatial and temporal distribution of microcystins in the Fuzhou Shanzai Reservoir[J]. Acta Scientiae Circumstantiae, 2011, 31(3): 533-546. |
| [5] |
姚玲爱, 赵学敏, 周广杰, 等. 广东省高州水库春季蓝藻水华成因初步探讨[J]. 湖泊科学, 2011, 23(4): 534-540. Yao L A, Zhao X M, Zhou G J, et al. Preliminary regulating factors of spring cyanobacteria bloom in Gaozhou Reservoir, Guangdong province[J]. Journal of Lake Sciences, 2011, 23(4): 534-540. |
| [6] |
施练东, 竺维佳, 张俊芳, 等. 亚热带水库浮游植物群落结构季节演替及其春季水华成因分析——以浙江汤浦水库为例[J]. 水生态学杂志, 2013, 34(2): 32-39. Shi L D, Zhu W J, Zhang J F, et al. Seasonal succession of phytoplankton community structures and analysis of spring water bloom in subtropical reservoir-as Tangpu Resevior in Zhejiang province[J]. Journal of Hydroecology, 2013, 34(2): 32-39. |
| [7] |
赵孟绪, 韩博平. 汤溪水库蓝藻水华发生的影响因子分析[J]. 生态学报, 2005, 25(7): 1554-1561. Zhao M X, Han B P. Analysis of factors affecting cyanobacteria bloom in a tropical reservoir (Tangxi Reservoir, China)[J]. Acta Ecologica Sinica, 2005, 25(7): 1554-1561. DOI:10.3321/j.issn:1000-0933.2005.07.004 |
| [8] | Bowes M J, Loewenthal M, Read D S, et al. Identifying multiple stressor controls on phytoplankton dynamics in the River Thames (UK) using high-frequency water quality data[J]. Science of the Total Environment, 2016, 569: 1489-1499. |
| [9] | González-Piana M, Fabián D, Piccardo A, et al. Dynamics of total microcystin LR concentration in three subtropical hydroelectric generation reservoirs in Uruguay, South America[J]. Bulletin of Environmental Contamination and Toxicology, 2017, 99(4): 488-492. DOI:10.1007/s00128-017-2158-7 |
| [10] | Remmal Y, Hudon C, Hamilton P B, et al. Forecasting the magnitude and composition of phytoplankton blooms in a eutrophic lowland river (Rivière Yamaska, Que., Canada)[J]. Canadian Journal of Fisheries and Aquatic Sciences, 2017, 74(8): 1298-1311. DOI:10.1139/cjfas-2016-0305 |
| [11] |
刘流, 刘德富, 肖尚斌, 等. 水温分层对三峡水库香溪河库湾春季水华的影响[J]. 环境科学, 2012, 33(9): 3046-3050. Liu L, Liu D F, Xiao S B, et al. Effects of thermal stratification on spring blooms in Xiangxi Bay of the Three Gorges reservoir[J]. Chinese Journal of Environmental Science, 2012, 33(9): 3046-3050. |
| [12] |
杨正健, 刘德富, 马骏, 等. 三峡水库香溪河库湾特殊水温分层对水华的影响[J]. 武汉大学学报(工学版), 2012, 45(1): 1-9, 15. Yang Z J, Liu D F, Ma J, et al. Effects of special vertical layered water temperatures on algal bloom in Xiangxi Bay of Three Gorges Reservoir[J]. Engineering Journal of Wuhan University, 2012, 45(1): 1-9, 15. |
| [13] | Yang J, Lv H, Yang J, et al. Decline in water level boosts cyanobacteria dominance in subtropical reservoirs[J]. Science of the Total Environment, 2016, 557-558: 445-452. DOI:10.1016/j.scitotenv.2016.03.094 |
| [14] |
吴卫菊, 杨凯, 汪志聪, 等. 云贵高原渔洞水库浮游植物群落结构及季节演替[J]. 水生态学杂志, 2012, 33(2): 69-75. Wu W J, Yang K, Wang Z C, et al. Community structure and seasonal succession of phytoplankton in Yudong Reservoir of Yungui-plateau[J]. Journal of Hydroecology, 2012, 33(2): 69-75. |
| [15] | Gou C X, Zhu G W, Paerl H W, et al. Extreme weather event may induce Microcystis blooms in the Qiantang River, Southeast China[J]. Environmental Science and Pollution Research, 2018, 25(22): 22273-22284. DOI:10.1007/s11356-018-2216-7 |
| [16] |
朱孔贤, 毕永红, 胡建林, 等. 三峡水库神农溪2008年夏季铜绿微囊藻(Microcystis aeruginosa)水华暴发特性[J]. 湖泊科学, 2012, 24(2): 220-226. Zhu K X, Bi Y H, Hu J L, et al. Characteristics of Microcystis aeruginosa bloom in summer 2008 in Shennong River of Three Gorges Reservoir[J]. Journal of Lake Sciences, 2012, 24(2): 220-226. DOI:10.3969/j.issn.1003-5427.2012.02.008 |
| [17] | Li Y, Zhang Y L, Shi K, et al. Monitoring spatiotemporal variations in nutrients in a large drinking water reservoir and their relationships with hydrological and meteorological conditions based on Landsat 8 imagery[J]. Science of the Total Environment, 2017, 599-600: 1705-1717. DOI:10.1016/j.scitotenv.2017.05.075 |
| [18] |
刘其根, 陈立侨, 陈勇. 千岛湖水华发生与主要环境因子的相关性分析[J]. 海洋湖沼通报, 2007(1): 117-124. Liu Q G, Chen L Q, Chen Y. Correlation between biomass reduction of silver carp and bighead carp and the occurrence of algal blooms in Lake Qiandaohu[J]. Transactions of Oceanology and Limnology, 2007(1): 117-124. DOI:10.3969/j.issn.1003-6482.2007.01.017 |
| [19] |
胡忠军, 孙月娟, 刘其根, 等. 浙江千岛湖深水区大型底栖动物时空变化格局[J]. 湖泊科学, 2010, 22(2): 265-271. Hu Z J, Sun Y J, Liu Q G, et al. Temporal and spatial distribution of profundal macrozoobenthic community in Lake Qiandao, Zhejiang, China[J]. Journal of Lake Sciences, 2010, 22(2): 265-271. |
| [20] | Li Y, Zhang Y L, Shi K, et al. Spatiotemporal dynamics of chlorophyll-a in a large reservoir as derived from Landsat 8 OLI data:understanding its driving and restrictive factors[J]. Environmental Science and Pollution Research, 2018, 25(2): 1359-1374. DOI:10.1007/s11356-017-0536-7 |
| [21] |
王明翠, 刘雪芹, 张建辉. 湖泊富营养化评价方法及分级标准[J]. 中国环境监测, 2002, 18(5): 47-49. Wang M C, Liu X Q, Zhang J H. Evaluate method and classification standard on lake eutrophication[J]. Environmental Monitoring in China, 2002, 18(5): 47-49. DOI:10.3969/j.issn.1002-6002.2002.05.018 |
| [22] |
陈宇炜, 陈开宁, 胡耀辉. 浮游植物叶绿素a测定的"热乙醇法"及其测定误差的探讨[J]. 湖泊科学, 2006, 18(5): 550-552. Chen Y W, Chen K N, Hu Y H. Discussion on possible error for phytoplankton chlorophyll-a concentration analysis using hot-ethanol extraction method[J]. Journal of Lake Sciences, 2006, 18(5): 550-552. DOI:10.3321/j.issn:1003-5427.2006.05.019 |
| [23] | 胡鸿钧, 李尧英, 魏印心, 等. 中国淡水藻类[M]. 上海: 上海科学技术出版社, 1980. |
| [24] | Boehrer B, Schultze M. Stratification of lakes[J]. Reviews of Geophysics, 2008, 46(2): RG2005. |
| [25] | Hambright K D, Gophen M, Serruya S. Influence of long-term climatic changes on the stratification of a subtropical, warm monomictic lake[J]. Limnology and Oceanography, 1994, 39(5): 1233-1242. DOI:10.4319/lo.1994.39.5.1233 |
| [26] | Dake J M K, Harleman D R F. Thermal stratification in lakes:analytical and laboratory studies[J]. Water Resources Research, 1969, 5(2): 484-495. DOI:10.1029/WR005i002p00484 |
| [27] | Rueda F, Schladow G. Mixing and stratification in lakes of varying horizontal length scales:scaling arguments and energy partitioning[J]. Limnology and Oceanography, 2009, 54(6): 2003-2017. DOI:10.4319/lo.2009.54.6.2003 |
| [28] | Liu M, Zhang Y L, Shi K, et al. Thermal stratification dynamics in a large and deep subtropical reservoir revealed by high-frequency buoy data[J]. Science of the Total Environment, 2019, 651: 614-624. DOI:10.1016/j.scitotenv.2018.09.215 |
| [29] | Zhang Y L, Wu Z X, Liu M L, et al. Thermal structure and response to long-term climatic changes in Lake Qiandaohu, a deep subtropical reservoir in China[J]. Limnology and Oceanography, 2014, 59(4): 1193-1202. DOI:10.4319/lo.2014.59.4.1193 |
| [30] | Nürnberg G K. Quantification of oxygen depletion in lakes and reservoirs with the hypoxic factor[J]. Lake and Reservoir Management, 2002, 18(4): 299-306. DOI:10.1080/07438140209353936 |
| [31] | Stefan H G, Hondzo M, Fang X, et al. Simulated long term temperature and dissolved oxygen characteristics of lakes in the north-central United States and associated fish habitat limits[J]. Limnology and Oceanography, 1996, 41(5): 1124-1135. DOI:10.4319/lo.1996.41.5.1124 |
| [32] | Zhang Y L, Wu Z X, Liu M L, et al. Dissolved oxygen stratification and response to thermal structure and long-term climate change in a large and deep subtropical reservoir (Lake Qiandaohu, China)[J]. Water Research, 2015, 75: 249-258. DOI:10.1016/j.watres.2015.02.052 |
| [33] | North R P, North R L, Livingstone D M, et al. Long-term changes in hypoxia and soluble reactive phosphorus in the hypolimnion of a large temperate lake:consequences of a climate regime shift[J]. Global Change Biology, 2014, 20(3): 811-823. DOI:10.1111/gcb.12371 |
| [34] |
余员龙.千岛湖浮游植物群落结构时空分布格局及其与主要环境因子的关系[D].上海: 上海海洋大学, 2010. Yu Y L. Spatial and temporal distribution of the phytoplankton community of Lake Qiandaohu and its relationship with the main environmental factors[D]. Shanghai: Shanghai Ocean University, 2010. http://www.wanfangdata.com.cn/details/detail.do?_type=degree&id=Y1821646 |
| [35] |
兰佳, 吴志旭, 李俊, 等. 新安江水库浮游植物群落特征及影响因素分析[J]. 中国环境监测, 2014, 30(2): 40-46. Lan J, Wu Z X, Li J, et al. Analysis of phytoplankton community and its preliminary influenced factors in Xin'an Jiang Reservoir[J]. Environmental Monitoring in China, 2014, 30(2): 40-46. DOI:10.3969/j.issn.1002-6002.2014.02.008 |
| [36] |
盛海燕, 吴志旭, 刘明亮, 等. 新安江水库近10年水质演变趋势及与水文气象因子的相关分析[J]. 环境科学学报, 2015, 35(1): 118-127. Sheng H Y, Wu Z X, Liu M L, et al. Water quality trends in recent 10 years and correlation with hydro-meteorological factors in Xin'anjiang Reservoir[J]. Acta Scientiae Circumstantiae, 2015, 35(1): 118-127. |
| [37] |
笪文怡, 黎云祥, 朱广伟, 等. 水文气象过程对千岛湖氮磷变化的影响[J]. 水生态学杂志, 2019, 40(5): 9-19. Da W Y, Li Y X, Zhu G W, et al. Influence of hydrometeorological processes on Nutrient Dynamics in Qiandao Lake[J]. Journal of Hydroecology, 2019, 40(5): 9-19. |
| [38] |
袁燕, 韩龙喜, 孙杨, 等. 基于典型降雨TN入库过程的水库水质响应数值模拟分析[J]. 安全与环境学报, 2014, 14(3): 189-193. Yuan Y, Han X L, Sun Y, et al. Simulated research of reservoir water quality response under typical rainfall[J]. Journal of Safety and Environment, 2014, 14(3): 189-193. |
| [39] |
张瑞国, 王克勤, 陈奇伯, 等. 昆明市水源区不同利用类型坡地径流氮和磷的输出特征[J]. 环境科学研究, 2009, 22(5): 607-611. Zhang R G, Wang K Q, Chen Q B, et al. Nitrogen and phosphorus loss characterstics in runoff from slopes with different land uses in Songhua Dam Reservoir of Kunming[J]. Research of Environmental Sciences, 2009, 22(5): 607-611. |
| [40] |
罗献宝, 文军, 骆东奇, 等. 洪水胁迫因子对千岛湖水质的影响与风险评价研究[J]. 中国生态农业学报, 2006, 14(3): 118-121. Luo X B, Wen J, Luo D Q, et al. Effect and risk assessment of flood stress on the water quality of Qian-dao Lake[J]. Chinese Journal of Eco-Agriculture, 2006, 14(3): 118-121. |
| [41] |
朱广伟, 金颖薇, 任杰, 等. 太湖流域水库型水源地硅藻水华发生特征及对策分析[J]. 湖泊科学, 2016, 28(1): 9-21. Zhu G W, Jin Y W, Ren J, et al. Characteristics of diatom blooms in a reservoir-water supply area and the countermeasures in Taihu Basin, China[J]. Journal of Lake Sciences, 2016, 28(1): 9-21. |
| [42] | Bowes M J, Gozzard E, Johnson A C, et al. Spatial and temporal changes in chlorophyll-a concentrations in the River Thames basin, UK:are phosphorus concentrations beginning to limit phytoplankton biomass?[J]. Science of the Total Environment, 2012, 426(2): 45-55. |
| [43] | Desortová B, Punčochář P. Variability of phytoplankton biomass in a lowland river:response to climate conditions[J]. Limnologica, 2011, 41(3): 160-166. DOI:10.1016/j.limno.2010.08.002 |
| [44] | Larroudé S, Massei N, Reyes-Marchant P, et al. Dramatic changes in a phytoplankton community in response to local and global pressures:a 24-year survey of the River Loire (France)[J]. Global Change Biology, 2013, 19(5): 1620-1631. DOI:10.1111/gcb.12139 |
| [45] | Waylett A J, Hutchins M G, Johnson A C, et al. Physico-chemical factors alone cannot simulate phytoplankton behaviour in a lowland river[J]. Journal of Hydrology, 2013, 497: 223-233. DOI:10.1016/j.jhydrol.2013.05.027 |
| [46] |
王轲, 王林同, 牛海凤, 等. 低温下氨氮对淡水浮游藻生长及群落结构影响的生态模拟研究[J]. 环境科学学报, 2012, 32(3): 731-738. Wang K, Wang L T, Niu H F, et al. Simulation of the effect of ammonia nitrogen on the growth and community structure of freshwater phytoplankton under low temperature[J]. Acta Scientiae Circumstantiae, 2012, 32(3): 731-738. |
| [47] | Vuorio K, Lagus A, Lehtimäki J M, et al. Phytoplankton community responses to nutrient and iron enrichment under different nitrogen to phosphorus ratios in the northern Baltic Sea[J]. Journal of Experimental Marine Biology and Ecology, 2005, 322(1): 39-52. DOI:10.1016/j.jembe.2005.02.006 |