 2019, Vol. 40
2019, Vol. 40



菜地是利用强度大、投入和产出高、受人类活动影响较大的一类农业土壤. Cd和Pb在土壤中移动性差、滞留时间长和不能被微生物降解等, 并可经过蔬菜等食物链进入人体, 严重损害人类健康, 因此必须采取有效措施阻控蔬菜对Cd和Pb的吸收[1~3].利用生物炭等化学钝化剂可以固定土壤中的重金属, 减少植物对重金属的吸收[4~6].但是长期使用化学钝化剂可能导致土壤理化性质的改变、土壤微生物数量和活性的下降和土壤肥力退化等问题[7].微生物固定修复技术是指利用土壤微生物对重金属的钝化作用可使重金属在土壤中沉淀或被吸附固定, 从而减少植物对重金属的吸收[8].重金属固定细菌不仅能够固定重金属, 改变土壤重金属赋存状态, 降低作物对重金属的吸收, 还能促进作物生长, 提高作物品质, 改善土壤质量, 环境友好, 有益于可持续发展等优点[9, 10].
长期高含量重金属污染会对土壤中的微生物有不同程度的毒性, 影响微生物种群的分布, 同时, 微生物为了应对重金属的胁迫, 它们也会进化出应对重金属毒性的机制, 包括金属还原、细胞通透性降低和细胞外隔离等, 因此从原位重金属污染蔬菜根际土壤中筛选得到的重金属固定细菌, 将具有更好的环境适应性和更高的阻控蔬菜吸收重金属的能力.虽然有部分学者采用高通量测序研究不同重金属污染条件下微生物种群的分布[11, 12], 但是长期高含量重金属污染条件下, 关于高含量重金属对生菜根际土壤中可培养细菌和重金属固定细菌的群落影响的研究较少.此外, 虽然有学者已报道使用重金属固定细菌降低向日葵、小麦和水稻等对重金属的吸收[13, 14], 但是目前关于具有阻控生菜吸收重金属的功能细菌仍较少.前期测定发现河南省新乡市郊区(35°36′N, 113°85′E)部分农田土壤中的Cd和Pb含量超过土壤安全标准几十到一百多倍[15].因此为了了解长期高含量重金属污染对生菜根际可培养细菌和重金属固定细菌群落的影响以及筛选得到高效阻控生菜吸收重金属的功能菌株, 本研究从河南省新乡市郊区农田采集两份意大利生菜根际土壤样品, 采用平板分离其可培养细菌, 采用摇瓶实验分析其吸附重金属能力, 比较不同样品可培养细菌和重金属固定细菌群落差异.采用水培实验筛选得到具有促进生菜生长和阻控生菜富集Cd和Pb的功能菌株, 以期为实施重金属污染农田生态整治与安全高效利用策略提供菌种资源和技术支撑.
1 材料与方法 1.1 样品采集及理化性质分析2018年5月, 在河南省新乡市郊区农田使用五点采样法采集两份意大利生菜根际土壤样品.第一份样品位于环宇集团有限公司厂区旁边(35°36′N, 113°85′E), 记为环宇样品(HY); 另一份样品位于环宇电池厂6 km外的大块镇附近农田(35°36′N, 113°80′E), 记为大块镇样品(DK), 位置如图 1.土壤pH、有机质、总N、总P和总K的测定参考文献[16].土壤样品中Cd和Pb的含量采用微波消解仪(SINEO MDS-6G, 中国)进行消解, 然后使用电感耦合等离子发射光谱仪(Inductively coupled plasma atomic emission spectrometry, ICP-OES, SHIMADZ ICPE-9820, 日本)进行测定[17].

|
图 1 生菜根际土采样地点示意 Fig. 1 Sampling sites of lettuce rhizosphere soil |
分别称取HY和DK样品各1.0 g放到含有50 mL无菌去离子水的三角瓶内, 150 r·min-1振荡30 min, 然后取1 mL土悬液梯度稀释成10-4、10-5和10-6, 涂布在LB固体平板上, 30℃培养3 d.对所有平板上长出的单菌落进行计数, 计算出HY和DK样品中可培养细菌的总数量.之后依据菌落大小、边缘形状和颜色等指标对HY和DK样品各随机挑取400个单菌落, 进行纯化和保藏.利用刘瑞等[18]的方法, 采用细菌基因组DNA提取试剂盒提取每个菌株的DNA, 采用细菌通用引物27F (5′-AGAGTTTGATCCTGCTCAG-3′)和1492R (5′-GGT TACCTTGTTACGACTT-3′)进行16S rRNA基因扩增, 扩增产物送到上海生工进行测序.
1.3 菌株吸附固定Cd和Pb能力测定配制含有Cd质量浓度为3 mg·L-1, Pb质量浓度为5 mg·L-1的LB液体培养基, 分装到三角瓶中, 每瓶50 mL.将培养18 h左右的供试菌株制备成菌悬液, 按1%的比例接入上述培养基中, 30℃, 180 r·min-1振荡培养24 h.培养结束后, 取5 mL发酵液, 5 000 r·min-1离心5 min, 采用ICP-OES测定上清液中Cd和Pb的质量浓度.通过计算得出各个菌株对Cd和Pb的吸附率(%), 计算公式如下:
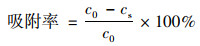
|
式中, c0表示溶液中重金属的初始质量浓度, mg·L-1; cs表示供试菌株上清液中重金属质量浓度, mg·L-1.
1.4 功能菌株对重金属的抗性和促生指标测定采用连续梯度平板法测定菌株对Cd和Pb的抗性和致死浓度(minimal inhibitory concentrations, MICs).即用接种环挑取供试菌株接种到含不同质量浓度Cd(100~1 000 mg·L-1, 每隔50 mg·L-1一个浓度梯度)和Pb(1 000~2 500 mg·L-1, 每隔100 mg·L-1一个浓度梯度)的LB固体培养基平板上, 30℃培养3 d, 若能长出菌落, 则重新转到含有相应重金属浓度的LB平板上, 30℃培养3 d.若两次均能长出菌落, 说明该菌株具有相应重金属浓度的抗性.菌株产吲哚乙酸(indole-3-acetic acid, IAA)的测定参考Jiang等[19]的方法, 产铁载体能力测定参考Rajkumar等[20]的方法, 产1-氨基环丙烷羧酸(1-Amino-1-cyclopropanecarboxylic acid, ACC)脱氨酶的能力测定方法参考Belimov等[21]的方法.
1.5 生菜水培实验选取若干籽粒饱满的两种生菜种子(意大利生菜和红花生菜, Lactuca sativa L.), 用75%乙醇进行消毒3 min, 再用无菌水漂洗3次, 然后置于水琼脂(0.4%)平板上, 30℃恒温培养, 待到种子长出四片小叶后, 挑取大小一致的生菜苗移栽于装有Hoagland营养液的盆卜中进行培养, 盆卜上层装有200 g石英砂, 起固定作用, 下层为Hoagland营养液.当生菜幼苗长到5 cm高后, 添加重金属, 使培养液中Cd终质量浓度为1 mg·L-1, Pb终质量浓度为3 mg·L-1.选取1.3节和1.4节实验中筛选到的高效固定Cd和Pb的菌株作为供试菌株进行接种.采用菌悬液浸根2 h的方式对生菜进行接种, 同时以不接种处理作为对照.接菌完成后继续培养30 d.将培养完成后的生菜从盆卜中取出, 根部用0.01 mol·L-1EDTA-2Na溶液浸泡10 min, 以除去生菜根部表面吸附的重金属, 然后用剪刀将生菜分为地上部和根部, 测量其鲜重, 然后置于烘箱80℃烘干, 测量生菜各组织干重.将烘干的生菜根和叶用粉碎机磨碎, 准确称取0.5 g进行微波消解, 采用ICP-OES测定消解液中Pb和Cd质量浓度.
1.6 数据处理采用SPSS 17.0软件进行数据统计分析, 用Microsoft Excel 2010软件绘图.
2 结果与分析 2.1 土壤理化性质HY和DK两个土样的基本理化性质如表 1所示. HY样品中的Cd (24.2 mg·kg-1)和Pb (503 mg·kg-1)的含量均显著高于DK样品中的Cd (0.8 mg·kg-1)和Pb (214 mg·kg-1)的含量.根据文献[22], 农田土壤重金属污染程度被分为5个等级(Ⅰ、Ⅱ、Ⅲ、Ⅳ和Ⅴ, 分别代表十分清洁、清洁、轻度污染、中度污染和重度污染), HY中的Cd和Pb污染均处于重度污染程度, 而DK中的Cd和Pb污染均处于中度污染程度. HY样品中的有机质(20.4 g·kg-1)、TP (2.5 g·kg-1)、TN (5.3 g·kg-1)和TK (5.46 g·kg-1)含量显著低于DK样品中的有机质(25.4 g·kg-1)、TP (6.5 g·kg-1)、TN (7.5 g·kg-1)和TK (12.5 g·kg-1)含量, 而HY样品的pH (6.67)显著高于DK样品(6.02)的pH.
|
|
表 1 土壤样品主要理化参数1) Table 1 Main physical and chemical properties of soil samples |
2.2 不同含量重金属对生菜根际土壤可培养细菌群落的影响
分别称取HY和DK样品0.5 g放入含有50 mL无菌去离子水的三角瓶内, 经过振荡和培养, 然后稀释涂布在LB固体平板上, 培养3 d后依据菌落大小、边缘形状和颜色等指标挑选单菌落, 最终HY和DK的平板上各挑取400个单菌落, 然后进行菌种鉴定, 结果如图 1所示. HY样品中的400个细菌隶属于3门14属.其中β-Proteobacteria是优势门, 占到51%, 而Actinobacteria占32%, Firmicutes占17%. Acinetobacter、Brevundimonas、Serratia、Arthrobacter和Pseudarthrobacter是优势属, 菌株的数量都超过40个.而DK样品中的400个菌隶属于4门30属, Firmicutes是优势门, 占到45%, 而β-Proteobacteria占31%, Actinobacteria占15%, Bacteroidetes占9%. Bacillus是绝对的优势属, 菌株数量达到102个, 这说明高含量的重金属降低了生菜根际土壤细菌在门和属水平上的分类单元, 降低了细菌的多样性.此外, 由表 1可知, DK样品每克土壤中的细菌数量为1.65×104个, 而HY样品每克土壤中的细菌数量为6.4×103个, 说明土壤中高含量的重金属抑制了细菌的生长和繁殖.
2.3 菌株吸附Cd和Pb能力比较对HY和DK样品各自分离得到的400株菌做了摇瓶吸附实验, 通过测定上清液中Cd和Pb的质量浓度, 得出菌株对溶液中重金属的吸附率.以重金属吸附率大于80%为分界线, 吸附率大于80%, 说明该菌株具有很强的重金属固定吸附能力; 吸附率小于80%, 说明该菌株固定吸附Cd和Pb的能力较弱, 结果如图 3所示. HY样品中的400株细菌, 对Cd和Pb吸附率均大于80%的菌株共有146株(82%~95%), 占比为36.5%, 其中Brevundimonas、Serratia、Arthrobacter和Pseudarthrobacter属具有固定重金属能力的菌株较多, 均大于10个菌株.而DK样品中的400株细菌, 对Cd和Pb吸附率均大于80%的菌株共有44株菌(81%~90%), 占比为11%, Bacillus是固定重金属能力的主要属, 含有26株细菌.这说明土壤中的细菌为了应对高含量重金属的胁迫, 它们也会进化出应对重金属毒性的机制, 进行固定钝化重金属, 从而得以繁殖下去.

|
图 3 HY和DK样品中具有高效重金属固定能力的菌株在属水平上的差异 Fig. 3 Genus-level differences in the heavy metal-immobilizing bacteria in the HY and DK samples |
对HY和DK样品中具有高Cd和Pb固定能力的190株细菌测定了产IAA、铁载体和ACC脱氨酶的能力以及对Cd和Pb的致死浓度测定.最终筛选到17株高产IAA、铁载体和ACC脱氨酶以及耐受高浓度Cd和Pb的菌株, 结果如表 2所示.这17株菌株均能分泌IAA, 介于20.35~79.63 mg·L-1之间, 其中Serratia liquefaciens HY-22、Bacillus thuringensis HY-53和Acinetobacter lwoffii HY-157产的IAA质量浓度最高, 分别为65.34、71.65和79.63 mg·L-1.这17株菌株均能耐受高浓度的Cd (250~750 mg·L-1)和Pb (1 200~2 300 mg·L-1), 其中菌株Serratia liquefaciens HY-22的Cd致死浓度为800 mg·L-1, Pb致死浓度为2 400 mg·L-1; 菌株Bacillus thuringensis HY-53的Cd致死浓度为750 mg·L-1, Pb致死浓度为2 300 mg·L-1; 菌株Acinetobacter lwoffii HY-157的Cd致死浓度为800 mg·L-1, Pb致死浓度为2 400 mg·L-1.
|
|
表 2 重金属固定细菌的促生特性和重金属抗性1) Table 2 Growth promotion characteristics and heavy metal resistance of the heavy metal-immobilizing bacteria |
2.5 功能菌株对生菜生长和富集Cd和Pb的影响
对HY样品中高产IAA、铁载体和ACC脱氨酶的3株重金属固定细菌Serratia liquefaciens HY-22、Bacillus thuringensis HY-53和Acinetobacter lwoffii HY-157进行生菜水培实验, 研究他们在不同重金属浓度下对生菜生长和富集Cd和Pb的影响, 结果如图 4和表 3所示.由图 4(a)可知, 在不添加重金属条件下, 与不接菌对照相比, 接种这3株细菌均能显著提高意大利生菜和花叶生菜根(47.5%~175%)和地上部(26.4%~53.1%)的干重.其中菌株Bacillus thuringensis HY-53的促生效果最好, 使意大利生菜根部干重提高了105%, 地上部干重提高了53.1%;使花叶生菜根部干重提高了175%, 地上部干重提高了52.8%.由图 4(b)可知, 在Cd为1 mg·L-1和Pb为3 mg·L-1条件下, 与不接菌对照相比, 接种这3株细菌均能显著提高意大利生菜和花叶生菜根(7.5%~77.6%)和地上部(15.4%~67.2%)的干重.其中菌株Serratia liquefaciens HY-22使意大利生菜根部干重提高了23.8%, 地上部干重提高了22.8%;使花叶生菜根部干重提高了7.5%, 地上部干重提高了36.2%.菌株Bacillus thuringensis HY-53使意大利生菜根部干重提高了77.6%, 地上部干重提高了51.2%;使花叶生菜根部干重提高了49.1%, 地上部干重提高了67.3%.菌株Acinetobacter lwoffii HY-157使意大利生菜根部干重提高了38.8%, 地上部干重提高了15.4%;使花叶生菜根部干重提高了15.1%, 地上部干重提高了38.2%.这说明这3株重金属固定细菌均具有促进生菜生长的能力.此外, 综合图 4来看, 没有接种菌的条件下, 与没有重金属胁迫相比, 在Cd为1 mg·L-1和Pb为3 mg·L-1条件下, 意大利生菜和花叶生菜根部干重降低了30.9%~36.1%, 地上部干重降低了41.4%~42.5%.

|
不同小写字母表示处理之间有显著差异(P < 0.05) 图 4 不同重金属浓度下重金属固定细菌对生菜各组织干重的影响 Fig. 4 Effects of heavy metal-immobilizing bacteria on the tissue dry weight of lettuce on the different levels of Cd and Pb-contaminated solution |
|
|
表 3 在Cd为1 mg·L-1和Pb为3 mg·L-1条件下重金属固定细菌对生菜各组织Cd和Pb含量的影响1) Table 3 Effects of heavy metal-immobilizing bacteria on the Cd and Pb concentration in lettuce in Cd (1 mg·L-1) and Pb (3 mg·L-1) |
由表 3可知, 在Cd为1 mg·L-1和Pb为3 mg·L-1条件下, 与不接菌对照相比, 接菌Serratia liquefaciens HY-22、Bacillus thuringensis HY-53和Acinetobacter lwoffii HY-157均能显著降低意大利生菜根部和地上部Cd(38.7%~66.6%)和Pb(34.7%~62.5%)的含量.不接菌处理组中意大利生菜地上部Cd含量为0.62 mg·kg-1, Pb含量为0.95 mg·kg-1; 而菌株Serratia liquefaciens HY-22使意大利生菜地上部Cd的含量下降到0.26 mg·kg-1, Pb的含量下降到0.44 mg·kg-1; 菌株Bacillus thuringensis HY-53使意大利生菜地上部Cd的含量下降到0.25 mg·kg-1, Pb的含量下降到0.42 mg·kg-1; 菌株Acinetobacter lwoffii HY-157使生菜地上部Cd的含量下降到0.38 mg·kg-1, Pb的含量下降到0.62 mg·kg-1.中华人民共和国食品安全国家标准-食物中污染物限量(GB 2762-2017)[23]中规定新鲜叶菜蔬菜Cd的限量标准是0.2 mg·kg-1, Pb的限量标准是0.3 mg·kg-1, 而菌株Serratia liquefaciens HY-22和Bacillus thuringensis HY-53均能使意大利生菜鲜叶中Cd(0.12~0.15 mg·kg-1)和Pb(0.18~0.21 mg·kg-1)的含量达到食品安全国家标准.与不接菌对照相比, 菌株Serratia liquefaciens HY-22和Acinetobacter lwoffii HY-157对花叶生菜根部和地上部Cd和Pb的含量无显著影响, 但菌株Bacillus thuringensis HY-53能够显著降低花叶生菜根部和地上部Cd(57.1%~70.1%)和Pb(58.1%~65.2%)的含量, 且花叶生菜鲜叶中Cd和Pb的含量达到食品安全国家标准.
3 讨论 3.1 高含量重金属提高生菜根际土壤重金属固定细菌的比例重金属对土壤微生物的影响, 主要表现在改变微生物群落结构, 降低生物量, 影响其生物活性等方面[24, 25]. Singh等[26]发现在不同浓度重金属污染下土壤中微生物的生物量也不相同, 在土壤中加入一定量的Cd, 能使土壌细菌数目由每克土壤4.8×107个减少为2 000个. Kalndeler等的[27]研究表明在污染矿区土壤中, 靠近矿区土壤的微生物量明显低于远离矿区的土壤, 并且距离矿区越近, 生物量的下降幅度越明显.在本研究中, HY样品(高浓度Cd和Pb)中每g土壤可培养细菌的数量为6.4×104个, 而DH样品(低浓度Cd和Pb)中每克土壤可培养细菌的数量为1.2×105个, 说明高浓度重金属抑制了部分细菌的生长繁殖, 减少了土壤中细菌的生物量.
重金属污染不仅能够降低土壤微生物的生物量, 同时也能影响土壤微生物群落结构, 即土壤微生物多样性, 包括物种多样性、功能多样性、遗传多样性和生态特征多样性[28, 29].在本研究中, Cd和Pb污染水平的提高改变了生菜根际土壤可培养细菌群落结构(图 2), HY根际土中400个细菌隶属于3门14属.其中β-Proteobacteria是优势门, 占到51%; Acinetobacter、Brevundimonas、Serratia、Arthrobacter和Pseudarthrobacter是优势属.而DK样品中的400个细菌隶属于4门30属, Firmicutes是优势门, 占到45%, 而β-Proteobacteria占31%, Actinobacteria占15%, Bacteroidetes占9%; Bacillus是绝对的优势属.这表明高浓度的重金属减少了生菜根际可培养细菌在门和属上的分类单位, 降低了生菜根际可培养细菌多样性.一般来说, 重金属毒性会作用于一部分抗性较低的微生物, 减少它们的丰度, 如Firmicutes和Bacteroidetes门, 研究表明它们的丰度与镉浓度呈负相关[30], 另一方面, 重金属会增加一些金属抗性细菌的丰度[31].本研究中发现HY样品中的400株细菌, 吸附率大于80%的菌株共有146株, 占比为36.5%, 其中Brevundimonas、Serratia、Arthrobacter和Pseudarthrobacter属具有固定重金属能力的菌株较多.而DK样品中的400株细菌, 吸附率大于80%的菌株共有44株菌, 占比为11%, Bacillus是固定重金属能力的主要属.这说明高浓度重金属显著提高了生菜根际土壤中重金属固定细菌的比例.在高镉铅污染下, Acinetobacter、Serratia和Bacillus属的一些细菌, 已被鉴定为重金属污染土壤中的优势属, 在抵抗和固定重金属作用中发挥某些特定作用[32]. β-Proteobacteria是高镉铅污染水平下生菜根际土壤最为重要的优势菌门, 它在之前的研究中已被认定为重金属污染土壤中最为重要的一类微生物[33]. Wei等[34]的研究发现土壤Mn浓度对超累积植物商陆(Phytolacca americana)叶片组织的内生细菌多样性和群落结构有显著影响, 随着土壤Mn浓度的增加, 其Shannon指数显著下降.重金属抗性菌随重金属浓度增加而增加, 而敏感菌则减少[35].导致本研究中HY和DK样品的意大利生菜根际土壤可培养细菌群落结构变化的原因可能为:某些重金属抗性低的根际细菌因不能适应高浓度的Cd和Pb而减少或消失, 如Mucilaginibacter、Chryseobacterium、Terribacillus和Staphylococcus等, 而那些具有重金属抗性和固定能力的根际细菌则具有竞争优势而大量增殖, 比如具有重金属抗性的Acinetobacter、Arthrobacter和Pseudarthrobacter, 同时作物为了抵抗Cd和Pb的毒害作用, 也会选择让对其有利的根际和内生细菌大量增殖.

|
图 2 HY和DK样品中可培养细菌在门和属上的群落结构 Fig. 2 Community structure of culturable bacteria by phyla and genera in the HY and DK samples |
重金属固定细菌对重金属的钝化作用可使重金属在土壤中沉淀或被吸附固定, 改变重金属赋存状态, “钝化”其生物有效性, 从而降低作物对重金属的吸收[8, 9, 24].林晓燕等[36]从重金属镉污染稻田土壤中筛选到两株对镉具有较强耐性的细菌菌株LCd1和LCd2, 并研究了两株菌株对7种重金属离子的吸附情况, 结果显示两株菌都对铅和镉有较高的吸附率. Chen等[17]从南京市栖霞山铅锌矿区中分离到1株重金属固定细菌Neorhizobium huautlense T1-17, 能够使辣椒果实中Cd和Pb含量比不接菌对照分别降低39.1%和81.3%.本研究从重金属污染农田生菜根际土壤中筛选到3株重金属固定细菌Serratia liquefaciens HY-22, Bacillus thuringensis HY-53和Acinetobacter lwoffii HY-157, 具有很强的固定钝化Cd和Pb的能力.与不接菌对照相比, 这3株菌均能显著降低意大利生菜根和叶中Cd(38.7%~66.6%)和Pb(34.7%~62.5%)的含量, 且菌株Serratia liquefaciens HY-22和Bacillus thuringensis HY-53能够使意大利生菜地上部Cd和Pb的含量达到国家食品安全标准.重金属固定细菌可以通过自身细胞壁的吸附、与磷酸盐络合、或通过细菌代谢产生的其他阴离子与重金属发生沉淀或吸附反应, 从而达到重金属固定的作用[37, 38].因此可以通过测定供试菌株对重金属离子的吸附能力判定供试菌株钝化重金属的能力.在本研究中, 从HY样品中分离出146株对Cd和Pb吸附率大于80%的菌株, 其中Serratia liquefaciens HY-22、Bacillus thuringensis HY-53和Acinetobacter lwoffii HY-157均具有很强的吸附固定Cd和Pb的能力.这说明菌株通过吸附固定溶液中的Cd和Pb, 降低溶液中Cd和Pb的质量浓度, 从而降低了生菜对Cd和Pb的吸收.但是微生物-植物相互作用存在着一个匹配的过程, 同一株菌对不同品种作物作用不尽相同.所以, 开展重金属固定细菌与蔬菜的匹配性研究, 具有重要的应用价值[39].本研究中Serratia liquefaciens HY-22、Bacillus thuringensis HY-53和Acinetobacter lwoffii HY-157均能显著降低意大利生菜根部和地上部Cd和Pb的含量, 但只有Bacillus thuringensis HY-53能显著降低花叶生菜根部和地上部Cd和Pb的含量, 这说明菌株Bacillus thuringensis HY-53具有更广的适应性和更强的阻控蔬菜吸收重金属的能力, 具有广阔的应用前景.
4 结论(1) 与低含量重金属污染(DK)生菜根际土壤相比, 高含量重金属污染(HY)生菜根际土壤中可培养细菌的生物量显著降低, 重金属固定细菌的比例显著提高.其中Brevundimonas、Serratia、Arthrobacter和Pseudarthrobacter为主要的固定重金属的细菌种属.
(2) 重金属固定细菌Serratia liquefaciens HY-22, Bacillus thuringensis HY-53和Acinetobacter lwoffii HY-157均能显著降低意大利生菜和花叶生菜根和地上部中Cd和Pb的含量.
(3) Bacillus thuringensis HY-53能够使意大利生菜和花叶生菜新鲜叶片中Cd和Pb的含量达到食品安全国家标准.
| [1] | Franca F C S S, Albuuerque A M A, Almeida A C, et al. Heavy metals deposited in the culture of lettuce (Lactuca sativa L.) by the influence of vehicular traffic in Pernambuco, Brazil[J]. Food Chemistry, 2017, 215: 171-176. DOI:10.1016/j.foodchem.2016.07.168 |
| [2] | León-Cañedo J A, Alarcón-Silvas S G, Fierro-Sañudo J F, et al. Mercury and other trace metals in lettuce (Lactuca sativa) grown with two low-salinity shrimp effluents:accumulation and human health risk assessment[J]. Science of the Total Environment, 2019, 650: 2535-2544. DOI:10.1016/j.scitotenv.2018.10.003 |
| [3] |
靳琪, 高红, 岳波, 等. 村镇生活垃圾重金属含量及其来源分析[J]. 环境科学, 2018, 39(9): 4385-4392. Jin Q, Gao H, Yue B, et al. Heavy metal content of rural living solid waste and related source and distribution analysis[J]. Environmental Science, 2018, 39(9): 4385-4392. |
| [4] | Wang D F, Zhang G L, Zhou L L, et al. Immobilizing arsenic and copper ions in manure using a nanocomposite[J]. Journal of Agricultural and Food Chemistry, 2017, 65(41): 8999-9005. DOI:10.1021/acs.jafc.7b02370 |
| [5] |
孙延琛, 曾祥峰, 杨立琼, 等. 强还原过程对设施菜地土壤重金属形态转化的影响[J]. 应用生态学报, 2017, 28(11): 3759-3766. Sun Y C, Zeng X F, Yang L Q, et al. Effects of strong reductive process on transformation of heavy metals in protected vegetable soil[J]. Chinese Journal of Applied Ecology, 2017, 28(11): 3759-3766. |
| [6] |
赵军超, 王权, 任秀娜, 等. 钙基膨润土辅助对堆肥及土壤Cu、Zn形态转化和白菜吸收的影响[J]. 环境科学, 2018, 39(4): 1926-1933. Zhao J C, Wang Q, Ren X N, et al. Effect of Ca-bentonite on Cu and Zn forms in compost and soil, and their absorption by Chinese cabbage[J]. Environmental Science, 2018, 39(4): 1926-1933. |
| [7] | Udeigwe T K, Eze P N, Teboh J M, et al. Application, chemistry, and environmental implications of contaminant-immobilization amendments on agricultural soil and water quality[J]. Environment International, 2011, 37(1): 258-267. DOI:10.1016/j.envint.2010.08.008 |
| [8] | Lin X Y, Mou R X, Cao Z Y, et al. Characterization of cadmium-resistant bacteria and their potential for reducing accumulation of cadmium in rice grains[J]. Science of the Total Environment, 2016, 569-570: 97-104. DOI:10.1016/j.scitotenv.2016.06.121 |
| [9] | Etesami H. Bacterial mediated alleviation of heavy metal stress and decreased accumulation of metals in plant tissues:Mechanisms and future prospects[J]. Ecotoxicology and Environmental Safety, 2018, 147: 175-191. DOI:10.1016/j.ecoenv.2017.08.032 |
| [10] | Wang Q, Chen L, He L Y, et al. Increased biomass and reduced heavy metal accumulation of edible tissues of vegetable crops in the presence of plant growth-promoting Neorhizobium huautlense T1-17 and biochar[J]. Agriculture, Ecosystems & Environment, 2016, 228: 9-18. |
| [11] | Zheng X, Chen L, Chen M M, et al. Functional metagenomics to mine soil microbiome for novel cadmium resisitance genetic determinants[J]. Pedosphere, 2019, 29: 298-310. DOI:10.1016/S1002-0160(19)60804-0 |
| [12] |
李毳, 景炬辉, 刘晋仙, 等. 十八河铜尾矿库坝面细菌群落时空动态及其驱动力[J]. 应用生态学报, 2018, 29(6): 1975-1982. Li C, Jing J H, Liu J X, et al. Spatiotemporal dynamics and driving forces of soil bacterial communities on the dam of Shibahe copper mine tailings in Shanxi, China[J]. Chinese Journal of Applied Ecology, 2018, 29(6): 1975-1982. |
| [13] | Marques A P G C, Moreira H, Franco A R, et al. Inoculating Helianthus annuus (sunflower) grown in zinc and cadmium contaminated soils with plant growth promoting bacteria-Effects on phytoremediation strategies[J]. Chemosphere, 2013, 92(1): 74-83. DOI:10.1016/j.chemosphere.2013.02.055 |
| [14] | Li Y, Pang H D, He L Y, et al. Cd immobilization and reduced tissue Cd accumulation of rice (Oryza sativa wuyun-23) in the presence of heavy metal-resistant bacteria[J]. Ecotoxicology and Environmental Safety, 2017, 138: 56-63. DOI:10.1016/j.ecoenv.2016.12.024 |
| [15] |
韩辉, 王晓宇, 蔡红, 等. 重金属固定植物促生细菌的筛选及其阻控小麦富集重金属效应[J]. 环境科学, 2019, 40(7): 3339-3346. Han H, Wang X Y, Cai H, et al. Isolation of heavy metal immobilized and plant growth-promoting bacteria and its effects on reducing heavy metal accumulation in wheat[J]. Environmental Science, 2019, 40(7): 3339-3346. |
| [16] | 鲍士旦. 土壤农化分析[M]. (第三版).北京: 中国农业出版社, 2000: 86-115. |
| [17] | Chen L, He L Y, Wang Q, et al. Synergistic effects of plant growth-promoting Neorhizobium huautlense T1-17 and immobilizers on the growth and heavy metal accumulation of edible tissues of hot pepper[J]. Journal of Hazardous Materials, 2016, 312: 123-131. DOI:10.1016/j.jhazmat.2016.03.042 |
| [18] |
刘瑞, 于章龙, 薛冲, 等. 市售豆芽携带细菌种属鉴定及酸性电解水的杀菌效果[J]. 食品科学, 2017, 38(17): 168-173. Liu R, Yu Z L, Xue C, et al. Identification of bacterial species and microbial inactivation by acidic electrolyzed water on commercial bean sprouts[J]. Food Science, 2017, 38(17): 168-173. DOI:10.7506/spkx1002-6630-201717028 |
| [19] | Jiang C Y, Sheng X F, Qian M, et al. Isolation and characterization of a heavy metal-resistant Burkholderia sp. from heavy metal-contaminated paddy field soil and its potential in promoting plant growth and heavy metal accumulation in metal-polluted soil[J]. Chemosphere, 2008, 72(2): 157-164. DOI:10.1016/j.chemosphere.2008.02.006 |
| [20] | Rajkumar M, Nagendran R, Lee K J, et al. Influence of plant growth promoting bacteria and Cr6+ on the growth of Indian mustard[J]. Chemosphere, 2006, 62(5): 741-748. DOI:10.1016/j.chemosphere.2005.04.117 |
| [21] | Belimov A A, Hontzeas N, Safronova V I, et al. Cadmium-tolerant plant growth-promoting bacteria associated with the roots of Indian mustard (Brassica juncea L. Czern.)[J]. Soil Biology and Biochemistry, 2005, 37(2): 241-250. DOI:10.1016/j.soilbio.2004.07.033 |
| [22] | Yang Z F, Yu T, Hou Q Y, et al. Geochemical evaluation of land quality in China and its applications[J]. Journal of Geochemical Exploration, 2014, 139: 122-135. DOI:10.1016/j.gexplo.2013.07.014 |
| [23] | GB 2762-2017, 食品安全国家标准食品中污染物限量[S]. |
| [24] | Han H, Wang Q, He L Y, et al. Increased biomass and reduced rapeseed Cd accumulation of oilseed rape in the presence of Cd-immobilizing and polyamine-producing bacteria[J]. Journal of Hazardous Materials, 2018, 353: 280-289. DOI:10.1016/j.jhazmat.2018.04.024 |
| [25] |
金笑, 寇文伯, 于昊天, 等. 鄱阳湖不同区域沉积物细菌群落结构、功能变化及其与环境因子的关系[J]. 环境科学研究, 2017, 30(4): 529-536. Jin X, Kou W H, Yu H T, et al. Environmental factors influencing the spatial distribution of sediment bacterial community structure and function in Poyang Lake[J]. Research of Environmental Sciences, 2017, 30(4): 529-536. |
| [26] | Singh S, Kayastha A M, Asthana R K, et al. Response of rhizobium leguminosarum to nickel stress[J]. World Journal of Microbiology and Biotechnology, 2001, 17(7): 667-672. DOI:10.1023/A:1012912700268 |
| [27] | Kandeler E, Kampichler C, Horak O. Influence of heavy metals on the functional diversity of soil microbial communities[J]. Biology and Fertility of Soils, 1997, 23(3): 299-306. |
| [28] |
刘沙沙, 付建平, 蔡信德, 等. 重金属污染对土壤微生物生态特征的影响研究进展[J]. 生态环境学报, 27(6): 1173-1178. Liu S S, Fu J P, Cai X D, et al. Effect of heavy metals pollution on ecological characteristics of soil microbes:a Review[J]. Ecology and Environmental Sciences, 27(6): 1173-1178. |
| [29] | Wang K, Zhang D M, Xiong J B, et al. Response of bacterioplankton communities to cadmium exposure in coastal water microcosms with high temporal variability[J]. Applied and Environmental Microbiology, 2015, 81(1): 231-240. DOI:10.1128/AEM.02562-14 |
| [30] | Pishchik V N, Vorob'ev N I, Provorov N A, et al. Mechanisms of plant and microbial adaptation to heavy metals in plant-microbial systems[J]. Microbiology, 2016, 85(3): 257-271. DOI:10.1134/S0026261716030097 |
| [31] | Wood J L, Zhang C, Mathews E R, et al. Microbial community dynamics in the rhizosphere of a cadmium hyper-accumulator[J]. Scientific Reports, 2016, 6: 36067. DOI:10.1038/srep36067 |
| [32] | Idris R, Trifonova R, Puschenreiter M, et al. Bacterial communities associated with flowering plants of the Ni hyperaccumulator Thlaspi goesingense[J]. Applied and Environmental Microbiology, 2004, 70(5): 2667-2677. DOI:10.1128/AEM.70.5.2667-2677.2004 |
| [33] | Chen Y, Jiang Y M, Huang H Y, et al. Long-term and high-concentration heavy-metal contamination strongly influences the microbiome and functional genes in Yellow River sediments[J]. Science of the Total Environment, 2018, 637-638: 1400-1412. DOI:10.1016/j.scitotenv.2018.05.109 |
| [34] | Wei Y, Hou H, Shangguan Y X, et al. Genetic diversity of endophytic bacteria of the manganese-hyperaccumulating plant Phytolacca americana growing at a manganese mine[J]. European Journal of Soil Biology, 2014, 62: 15-21. DOI:10.1016/j.ejsobi.2014.02.011 |
| [35] | Rajkumar M, Ae N, Freitas H. Endophytic bacteria and their potential to enhance heavy metal phytoextraction[J]. Chemosphere, 2009, 77(2): 153-160. DOI:10.1016/j.chemosphere.2009.06.047 |
| [36] |
林晓燕, 牟仁祥, 曹赵云, 等. 耐镉细菌菌株的分离及其吸附镉机理研究[J]. 农业环境科学学报, 2015, 34(9): 1700-1706. Lin X Y, Mou R X, Cao Z Y, et al. Isolation and cadmium adsorption mechanisms of cadmium-resistant bacteria strains[J]. Journal of Agro-Environment Science, 2015, 34(9): 1700-1706. |
| [37] | Huang J H, Yuan F, Zeng G M, et al. Influence of pH on heavy metal speciation and removal from wastewater using micellar-enhanced ultrafiltration[J]. Chemosphere, 2017, 173: 199-206. DOI:10.1016/j.chemosphere.2016.12.137 |
| [38] | Xu C R, He S B, Liu Y M, et al. Bioadsorption and biostabilization of cadmium by Enterobacter cloacae TU[J]. Chemosphere, 2017, 173: 622-629. DOI:10.1016/j.chemosphere.2017.01.005 |
| [39] |
伍惠, 钟喆栋, 樊伟, 等. 8株优良大豆根瘤菌与不同地区27个大豆主栽品种的匹配性研究[J]. 大豆科学, 2017, 36(3): 405-418. Wu H, Zhong Z D, Fan W, et al. Symbiotic compatibility among eight elite soybean rhizobia strains and twenty-seven soybean cultivars from different planting regions[J]. Soybean Science, 2017, 36(3): 405-418. |