 2019, Vol. 40
2019, Vol. 40



2. 重庆大学三峡库区环境与生态部重点实验室, 重庆 400045
2. Key Laboratory of Three Gorges Reservoir Region's Eco-Environment, Ministry of Education, Chongqing University, Chongqing 400045, China
厌氧氨氧化(anaerobic ammonium oxidation, ANAMMOX)是一种新型脱氮工艺, 因其具有脱氮效率高、曝气量少、无需额外碳源等优点成为近年来研究热点.目前厌氧氨氧化工艺已被用于处理各种废水[1~4].根据厌氧氨氧化反应方程式[5]可知, 经过厌氧氨氧化反应后仍然有10%以上的NO3--N无法去除, 此外单一的厌氧氨氧化工艺对COD没有去除效果, 但废水中往往存在较高的COD.为了去除厌氧氨氧化产生的NO3--N和废水中的COD, 有必要将反硝化工艺与厌氧氨氧化工艺结合, 厌氧氨氧化耦合反硝化(simultaneous ANAMMOX and denitrification, SAD)能够实现同步脱氮除碳的目的.
Li等[6]的研究表明, 厌氧氨氧化和反硝化反应可在同一反应器中发生, 并具有良好的脱氮除碳效果.然而有研究表明, 高COD浓度或高碳氮比对厌氧氨氧化反应具有抑制作用[7], 因此启动SAD工艺需要将碳氮比控制在合理范围内.张诗颖等[8]发现采用ABR反应器时厌氧氨氧化耦合反硝化最佳协同条件为COD/TN=1, TN平均去除率为93%, 比单一厌氧氨氧化理论TN去除率高4%.张美雪等[9]的研究表明在SAD系统稳定期NH4+-N、NO2--N和TN的去除率依次为93.84%、94.62%和86.46%.
以上研究均在低氮进水浓度下(NH4+-N浓度30~80 mg ·L-1、NO2--N浓度45~160 mg ·L-1)进行, 而采用SAD处理高氮浓度废水的研究鲜有报道, 另外现有关于SAD的研究大多注重碳氮去除效果的分析, 缺乏对微生物群落变化的分析, 同时缺少对反应器内部不同部位的菌群变化分析.因此, 本实验采用高氮低碳废水(NH4+-N浓度190.63~246.39 mg ·L-1, NO2--N浓度216.28~267.92 mg ·L-1, COD浓度108~333 mg ·L-1)启动SAD工艺, 探究SAD启动过程中碳氮去除机制与微生物群落的关系, 以期为SAD应用于处理实际废水提供技术支持.
1 材料与方法 1.1 SAD工艺装置与运行实验采用膨胀颗粒污泥床(EGSB), 主要结构见图 1, 反应器高1.3 m, 反应区内径80 mm, 有效容积9 L.该厌氧氨氧化反应器已稳定运行2年, TN去除率85%以上.反应器主体采用水浴加热, 温度35~37℃, 反应器进水DO控制在(0.2±0.02) mg ·L-1, HRT为9 h, 实验阶段进水TN负荷为1.13~1.44 kg ·(m3 ·d)-1.

|
图 1 EGSB反应器运行装置示意 Fig. 1 Schematic diagram of EGSB reactor operation |
本实验采用人工配水, 进水pH均控制在7.8~8.2, 水质指标如表 1所示.
|
|
表 1 EGSB反应器进水水质 Table 1 Influent water quality of EGSB reactor |
DO、pH通过溶解氧仪(520M-01A, Thermo, USA)和pH计(8107UWMMD, Thermo, USA)测量.水质测定均采用国家标准方法[10]:NH4+-N采用纳氏试剂分光光度法测定, NO2--N采用N-(1-萘基)-乙二胺光度法测定, NO3--N采用紫外分光光度法测定, COD采用快速消解分光光度法测定, MLVSS采用恒重法测定.污泥的厌氧氨氧化活性和反硝化活性参考黄佳璐等[11]和徐峥勇[12]的测定方法并作适当调整, 具体步骤如下:采用100 mL原水和3 g(湿重)污泥置于150 mL血清瓶中, 测定厌氧氨氧化活性时控制NH4+-N和NO2--N的初始浓度分别为100 mg ·L-1和120 mg ·L-1左右, 测定反硝化活性时控制NO3--N和葡萄糖初始浓度分别为100 mg ·L-1和500 mg ·L-1左右, pH值均为7.8~8.1, 实验通过高纯氮气曝气30 min保证厌氧条件, 然后将血清瓶避光置于37℃、150 r · min-1的水浴恒温振荡器中, 采用注射器每1 h取1次水样, 测定NH4+-N、NO2--N和NO3--N浓度.根据基质降解曲线计算出污泥的厌氧氨氧化活性[以NH4+-N/VSS计, g ·(g ·d)-1]和反硝化活性[以NO3--N/VSS计, g ·(g ·d)-1].
1.3 脱氮贡献率计算方法厌氧氨氧化脱氮贡献率A和反硝化脱氮贡献率D参考魏思佳等[13]的计算方法:
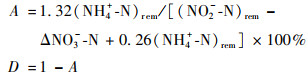
|
(1) |
式中, (NH4+-N)rem、(NO2--N)rem分别为NH4+-N、NO2--N去除量(mg ·L-1), ΔNO3--N为NO3--N变化量(mg ·L-1).
1.4 高通量测序分析方法在SAD启动实验前后分别提取反应器内上层和下层污泥样品中的微生物DNA, 其中Up1代表SAD启动前上层污泥, Down1代表SAD启动前下层污泥, Up2代表SAD启动后上层污泥, Down2代表SAD启动后下层污泥.采用高通量测序方法检测4组样品中的微生物[14], 测序由上海美吉生物提供(上海, 中国).
2 结果与讨论 2.1 SAD启动过程中脱氮除碳性能分析SAD启动采用逐步提高COD负荷的方式, 图 2表示启动过程中氮素去除特征.添加COD后NH4+-N出水浓度为0.15~11.64 mg ·L-1, 平均去除率为98.54%, 可见COD浓度的增加并没有使氨氮去除率下降. NO2--N出水浓度为0.52~7.96 mg ·L-1, 出水较稳定, 平均去除率为98.05%.从图 2(c)可知, NO3--N出水浓度随着COD的增加逐渐下降, 从启动前的48.01~52.04 mg ·L-1降至第3阶段的5.11~13.03 mg ·L-1.启动过程中TN出水浓度为16.56~48.85 mg ·L-1, 且随着COD的增加TN浓度逐渐降低, 第1阶段TN平均去除率为90.58%, 第2阶段为94.33%, 第3阶段为95.60%, 可见TN去除率随着COD增加而增加, 且增加速度趋缓.启动实验结果表明COD为306~333 mg ·L-1(COD/TN约0.75)时厌氧氨氧化耦合反硝化效果最好, TN最高去除率为95.60%, 比厌氧氨氧化理论TN去除率高了6.8%, 这在目前报道的SAD系统对TN去除率的研究中处于较高水平[8, 9], 表明本实验厌氧氨氧化与反硝化反应达到了良好地协同脱氮效果.

|
图 2 SAD启动过程中氮素去除特征 Fig. 2 Characteristics of nitrogen removal during SAD startup process |
图 3表示, SAD启动过程中三氮计量比关系及厌氧氨氧化和反硝化脱氮贡献率变化, 可见启动过程中ΔNO2--N/ΔNH4+-N并没有明显变化, 保持在1.05~1.21范围内, 略低于厌氧氨氧化反应的理论值, 但ΔNO3--N/ΔNH4+-N有明显下降的趋势, 第1阶段该比值范围为0.15~0.17, 第2阶段为0.07~0.10, 第3阶段为0.03~0.07.随着COD浓度的提高, NO2--N去除量没有增加, 但NO3--N消耗量明显增加, 这是由于有机物浓度增加后反应器内异养反硝化菌快速生长, 且优先利用NO3--N作为反硝化氮源, 这与刘晓萍[15]的研究结果相同.

|
图 3 SAD启动过程中计量比关系及厌氧氨氧化和反硝化脱氮贡献率变化 Fig. 3 Characteristics of the measurement ratio relation ship and TN removal contribution of ANAMMOX and denitrification during SAD startup process |
由图 3可知, 随着COD的增加厌氧氨氧化脱氮贡献率不断下降, 启动前厌氧氨氧化平均脱氮贡献率为98.5%, 第1阶段为94.1%, 第2阶段为90.7%, 第3阶段为83.9%, 相反地, 反硝化脱氮贡献率从启动前的1.5%增至第3阶段的16.1%, 呈逐渐上升趋势, 这主要是因为异养反硝化菌在反应器内比例逐渐增加.由于COD/TN控制在0.75以下, 碳源不足导致反硝化菌无法正常进行反硝化反应, 其生长增加到一定程度后得到抑制, 增加的反硝化菌并没有和厌氧氨氧化菌抢夺基质NO2--N, 而是利用厌氧氨氧化反应产生的NO3--N, 因此反应器内厌氧氨氧化菌的脱氮性能并没有受到影响. Kumar等[16]指出SAD耦合成功的必要条件是反硝化反应不会和厌氧氨氧化反应争夺NO2--N, 本实验结果也证明了这一点.
图 4表示, SAD启动过程中COD去除特征, 可知第1阶段COD出水浓度为38~46 mg ·L-1, 第2阶段为48~68 mg ·L-1, 第3阶段为58~81 mg ·L-1, 可见, COD进水浓度的大幅增加并没有导致出水浓度大幅增加, 一方面是异养反硝化作用逐渐增强的影响, 另一方面可能是由于反应器内存在一定比例的厌氧消化菌, COD增加后反应器内厌氧消化菌得到生长, 异养反硝化和厌氧消化的共同作用使得COD去除率维持在较高水平, 最终稳定在75%~80%之间.

|
图 4 SAD启动过程中COD去除特征 Fig. 4 Characteristics of COD removal during SAD startup process |
通过对反应器内上层污泥和下层污泥取样分析, 计算污泥的厌氧氨氧化活性和反硝化活性, 见表 2.
|
|
表 2 反应器内污泥厌氧氨氧化活性和反硝化活性 Table 2 ANAMMOX and denitrification sludge activity in the reactor |
由表 2可知反应器下层污泥厌氧氨氧化活性明显优于上层, 主要是因为反应器由下部进水, 下部较高的基质浓度更有利于厌氧氨氧化菌的富集. SAD启动成功后, 反应器上层污泥厌氧氨氧化活性下降, 由0.08 g ·(g ·d)-1降至0.027 g ·(g ·d)-1,主要是因为有机物进入反应器后异养菌增长较快, 且异养菌多呈悬浮态, 在上层污泥中较多, 上层厌氧氨氧化菌变少, 厌氧氨氧化活性降低.启动成功后下层污泥厌氧氨氧化活性几乎不变, 由0.114 g ·(g ·d)-1变为0.11 g ·(g ·d)-1, 主要是因为下层颗粒状厌氧氨氧化污泥较多, 且具有一定的抗冲击性, 异养菌的生长对厌氧氨氧化菌影响较少.
从反硝化活性看, SAD启动前, 反应器内上层污泥反硝化活性略高于下层污泥, 启动后上下层污泥反硝化活性都有明显提高, 分别由0.057 g ·(g ·d)-1和0.049 g ·(g ·d)-1增加为0.241 g ·(g ·d)-1和0.213 g ·(g ·d)-1, 且上层污泥反硝化活性更高, 这可能是因为厌氧氨氧化颗粒污泥沉降性能好, 在反应器底部密度较大, 而悬浮态的异养反硝化菌密度较小, 在反应器上部生长更快, 因此上层污泥的反硝化活性优于下层污泥.
2.3 SAD典型周期内基质变化特征图 5表示典型周期(Phase3)内各基质变化曲线, 典型周期结束后测得MLVSS浓度为3156 mg ·L-1.由基质变化曲线可看出, NO2--N和NH4+-N降解可分为两个阶段:快速降解阶段(0~10 h)和稳定阶段(10~18 h).在快速降解阶段, NO2--N和NH4+-N降解过程接近零级反应.在稳定阶段, NO2--N和NH4+-N降解缓慢并趋于稳定. NO3--N浓度在反应中先增后减, 且在10 h时浓度达到最高, 安芳娇等[17]的研究也发现相同的现象, 这可能是由于在反应前期NO2--N浓度较高, 反硝化菌在利用氮源时优先利用NO2--N, 厌氧氨氧化反应产生的NO3--N逐步累积, 后期NO2--N浓度减小, 更多的NO3--N被反硝化菌利用, NO3--N浓度又逐渐降低.由图 5可知, 当NO2--N和NO3--N浓度稳定后, COD浓度依然在降低, 这可能是反应器中厌氧消化菌和微生物繁殖消耗碳源的共同作用.

|
图 5 SAD典型周期内基质变化特征 Fig. 5 Characteristics of matrix changes during a typical cycle of SAD |
微生物多样性和丰富度如表 3所示, 本实验中4组微生物测序样本覆盖率(coverage)均大于0.99, 能保证测序结果的准确性. Shannon和Simpson代表样品多样性, Sobs、Ace和Chao代表样品丰富度.由表 3可知SAD启动前上层污泥Shannon和Sobs指数均高于下层污泥, 因此反应器上层污泥微生物多样性和丰富度均高于下层污泥, 这是由于下层污泥富集的厌氧氨氧化菌较多, 微生物种类相对较少. SAD启动后上层污泥Shannon和Sobs指数均有所下降, 这表明SAD启动后反应器上下层污泥微生物多样性和丰富度有所上升, 主要是因为有机物加入后异养菌得到生长, 反应器内微生物种类增多.
|
|
表 3 反应器内微生物多样性和丰富度 Table 3 Microbial diversity and abundance in the reactor |
2.4.2 微生物门分类水平组成分析
将微生物检测相对丰度>1%的菌门作为主要菌门, SAD启动前后反应器上层污泥和下层污泥微生物门水平组成如图 6所示. 4组微生物群落中共发现9个主要菌门, 分别为绿弯菌门(Chloroflexi)、浮霉菌门(Planctomycetes)、厚壁菌门(Firmicutes)、装甲菌门(Armatimonadetes)、变形菌门(Proteobacteria)、放线菌门(Actinobacteria)、拟杆菌门(Bacteroidetes)、绿菌门(Chlorobi)和Patescibacteria菌门.其中绿菌门仅是SAD启动前的主要菌门, 而Patescibacteria仅是SAD启动后的主要菌门, 二者相对丰度较小(1.22%~1.98%), 其它7个菌门在SAD启动前后均为反应器内主要菌门.

|
图 6 反应器内微生物门水平组成 Fig. 6 Composition of microorganisms at the phylum level in the reactor |
绿弯菌门是厌氧氨氧化系统中常见的菌门, 汪瑶琪等[18]在厌氧氨氧化系统中也发现较大丰度的绿弯菌门. Chen等[19]的研究发现, 绿弯菌门是兼性厌氧菌, 有利于厌氧氨氧化菌形成颗粒污泥. Kindaichi等[20]的研究表明, 绿弯菌门分解死亡的厌氧氨氧化菌, 且它的丝状结构有利于生物膜的形成, 可见绿弯菌门对于厌氧氨氧化系统具有重要作用.由图 6可知, SAD启动后绿弯菌门相对丰度有所增加, 反应器上下部分别由35.87%和31.79%升至40.94%和38.92%, 这是因为反应器内属于绿弯菌门的菌种主要为厌氧发酵菌, COD增加后部分厌氧发酵菌得到生长, 绿弯菌门丰度增加.浮霉菌门是厌氧氨氧化菌所在的主要菌门, 在上下层污泥中相对丰度分别为20.16%和32.29%, 可见下层污泥厌氧氨氧化菌富集效果优于上部, 这与污泥活性的分析结果相同. SAD启动后上下层污泥浮霉菌门相对丰度均有所下降, 表明COD增加后异养菌的生长抑制了厌氧氨氧化菌的生长, 但是这种抑制效果并没有影响厌氧氨氧化脱氮效果, 这可从较高的NH4+-N去除率得到验证.相关研究表明变形菌门和厚壁菌门对反硝化有重要作用[21], 启动前后上下层污泥变形菌门相对丰度分别由8.15%和8.45%增至13.84%和14.07%, 这主要是异养反硝化菌生长增殖的作用.装甲菌门是厌氧氨氧化系统中常见的菌门, 其相对丰度在SAD启动后略微下降. Zhang等[22]的研究发现, 厌氧氨氧化系统中装甲菌门的存在会导致NH4+-N过量消耗, 这可能是本实验中ΔNO2--N/ΔNH4+-N低于理论值的原因.
2.4.3 功能微生物群属分析将微生物检测相对丰度>0.5%的菌属作为主要分析对象, 4组微生物样品共检测出27个属, 微生物属水平组成如图 7所示. SAD启动前反应器内部优势菌属为Candidatus Brocadia(18.24%~30.84%)、norank_f_Anaerolineaceae(13.33%~14.42%)、norank_o_SBR1031 (6.71%~7.22%)、norank_o_Fimbriimonadales(5.03%~5.62%)、Limnobacter(4.32%~4.52%)和Clostridium_sensu_stricto_ 1 (3.82%~4.64%)等. SAD启动后反应器内部优势菌属为Candidatus Brocadia(15.91%~25.69%)、OLB 13 (11.09%~23.98%)、norank_o_SBR1031 (5.63%~14.81%)、norank_o_Fimbriimonadales(4.84%~4.99%)、norank_f_A4b(4.25%~6.37%)和Limnobacter(3.53%~4.73%)等. Candidatus Brocadia为反应器内检测出的唯一厌氧氨氧化菌, 其相对丰度和浮霉菌门相当, 表明本实验Candidatus Brocadia为厌氧氨氧化菌优势菌种.文献[23]的研究发现Candidatus Kuenenia适合存在于低基质浓度废水中, 而Candidatus Brocadia主要存在于高基质浓度废水中, 本实验进水NH4+-N平均浓度在200mg ·L-1以上, 属于高基质浓度废水, 更适合Candidatus Brocadia生长. Limnobacter是一种异养菌, 属于变形菌门, SAD启动前后Limnobacter丰度没有较大变化.有研究表明Limnobacter能和厌氧氨氧化菌共生, 保护厌氧氨氧化菌免受外界恶劣环境的影响[24], Candidatus Brocadia相对丰度在SAD启动后有所下降, 但下降不明显, 仍然表现出较高的厌氧氨氧化活性, 这可能与Limnobacter的保护作用有关.

|
图 7 反应器内微生物属水平组成 Fig. 7 Composition of microorganisms at the genus level in the reactor |
norank_f_Anaerolineaceae、norank_o_SBR1031、OLB13 和norank_f_A4b均属于绿弯菌门厌氧绳菌纲(Anaerolineae), 有研究表明该类微生物为厌氧消化核心微生物种群[25], SAD启动后该类厌氧消化菌丰度有所增加, 由于实验采用葡萄糖作为碳源, 可能是葡萄糖的添加促进了厌氧消化作用.另有研究表明隶属于厚壁菌门的土孢杆菌(Terrisporobacter)和毛球菌(Trichococcus)也属于厌氧消化菌[26, 27], 在本实验中也检测到.有研究表明, 氢噬菌(Hydrogenophaga)、索氏菌(Thauera)、假单胞菌(Pseudomonas)、副球菌(Paracoccus)和Denitratisoma菌都具有反硝化功能, 其中索氏菌、副球菌和Denitratisoma主要发生异养反硝化[28~30], 假单胞菌既可以发生自养反硝化[31], 也可以发生异养反硝化, 而氢噬菌主要发生自养反硝化[32].另外有研究发现属于厚壁菌门的芽孢杆菌(Bacillus)也具有反硝化功能[33].
图 8反映了SAD启动前后上下层污泥不同功能菌属相对丰度及其变化.可见SAD启动后厌氧消化菌相对丰度有较大增加, 反应器上下部厌氧消化菌相对丰度分别由29.87%和27.14%增至38.56%和35.82%.厌氧氨氧化菌在SAD启动后相对丰度有所下降, 上下层污泥厌氧氨氧化菌相对丰度分别由18.24%和30.84%降至15.91%和25.69%. SAD启动后反硝化菌相对丰度增幅较大, 反应器上下部变形菌门相对丰度分别由2.76%和2.79%增至9.10%和10.88%.菌群变化的主要原因在于有机物进入反应器后, 异养反硝化菌在有外部碳源情况下快速生长, 外加碳源和异养菌的增加抑制了厌氧氨氧化菌的生长, 部分厌氧氨氧化菌死亡, 厌氧氨氧化菌相对丰度下降, 死亡的厌氧氨氧化菌细胞和外加碳源为厌氧消化菌提供了营养物质, 厌氧消化菌得到生长, 相对丰度增加.

|
图 8 功能微生物相对丰度变化 Fig. 8 Changes of relative abundance of functional microorganisms |
(1) 在高氮进水基质下启动SAD反应, 随着进水COD浓度的增加, NH4+-N和NO2--N出水稳定, 平均去除率在98%以上; TN去除率逐渐升高, 第3阶段TN平均去除率为95.6%, 比厌氧氨氧化理论TN去除率高6.8%.随着进水COD浓度的增加, ΔNO3--N/ΔNH4+-N明显下降, 从0.15~0.17逐步降至0.03~0.07; COD浓度增加的过程中厌氧氨氧化脱氮贡献率逐渐下降, 反硝化脱氮贡献率逐渐上升, COD去除率逐步增加.
(2) 反应器内下层污泥厌氧氨氧化活性明显高于上层污泥, 下层污泥反硝化活性略高于上层污泥. SAD启动后厌氧氨氧化活性略微降低, 污泥反硝化活性明显增加, 上层污泥反硝化活性更高.
(3) 典型周期内, 即SAD脱氮效果最佳时, NO2--N和NH4+-N在快速降解阶段降解过程接近零级反应, NO3--N浓度在反应中先增后减, 且在10 h时浓度达到最高.
(4) 高通量测序分析结果表明反应器内微生物群落特征与SAD脱氮除碳性能密切相关.反应器内与脱氮除碳有关的功能微生物主要有厌氧氨氧化菌、反硝化菌和厌氧消化菌. SAD启动后反应器内厌氧氨氧化菌丰度相对减少, 厌氧消化菌和异养反硝化菌丰度明显增加.
| [1] | Akgul D, Aktan C K, Yapsakli K, et al. Treatment of landfill leachate using UASB-MBR-SHARON-Anammox configuration[J]. Biodegradation, 2013, 24(3): 399-412. DOI:10.1007/s10532-012-9597-y |
| [2] |
王欢, 裴伟征, 李旭东, 等. 低碳氮比猪场废水短程硝化反硝化-厌氧氨氧化脱氮[J]. 环境科学, 2009, 30(3): 815-821. Wang H, Pei W Z, Li X D, et al. Removing nitrogen from low-C/N-piggery-wastewater using shortcut nitrification/denitrification-ANAMMOX[J]. Environmental Science, 2009, 30(3): 815-821. DOI:10.3321/j.issn:0250-3301.2009.03.032 |
| [3] | Leal C D, Pereira A D, Nunes F T, et al. ANAMMOX for nitrogen removal from anaerobically pre-treated municipal wastewater:effect of COD/N ratios on process performance and bacterial community structure[J]. Bioresource Technology, 2016, 211: 257-266. DOI:10.1016/j.biortech.2016.03.107 |
| [4] |
杨振琳, 于德爽, 李津, 等. 海藻糖强化厌氧氨氧化耦合反硝化工艺处理高盐废水的脱氮除碳效能[J]. 环境科学, 2018, 39(10): 4612-4620. Yang Z L, Yu D S, Li J, et al. Enhanced nitrogen and carbon removal performance of simultaneous ANAMMOX and denitrification (SAD) with trehalose addition treating saline wastewater[J]. Environmental Science, 2018, 39(10): 4612-4620. |
| [5] | Strous M, Kuenen J G, Jetten M S M. Key physiology of anaerobic ammonium oxidation[J]. Applied and Environmental Microbiology, 1999, 65(7): 3248-3250. |
| [6] | Li J, Qiang Z M, Yu D S, et al. Performance and microbial community of simultaneous ANAMMOX and denitrification (SAD) process in a sequencing batch reactor[J]. Bioresource Technology, 2016, 218: 1064-1072. DOI:10.1016/j.biortech.2016.07.081 |
| [7] | Chamchoi N, Nitisoravut S, Schmidt J E. Inactivation of ANAMMOX communities under concurrent operation of anaerobic ammonium oxidation (ANAMMOX) and denitrification[J]. Bioresource Technology, 2008, 99(9): 3331-3336. DOI:10.1016/j.biortech.2007.08.029 |
| [8] |
张诗颖, 吴鹏, 宋吟玲, 等. 厌氧氨氧化与反硝化协同脱氮处理城市污水[J]. 环境科学, 2015, 36(11): 4174-4179. Zhang S Y, Wu P, Song Y L, et al. Nitrogen removal using ANAMMOX and denitrification for treatment of municipal sewage[J]. Environmental Science, 2015, 36(11): 4174-4179. |
| [9] |
张美雪, 李芸, 李军, 等. 低浓度乙酸盐诱导下厌氧氨氧化与异养反硝化高效耦合脱氮[J]. 环境工程学报, 2016, 10(11): 6127-6132. Zhang M X, Li Y, Li J, et al. Nitrogen removal performance of ANAMMOX coupling heterotrophic denitrification in low concentration of acetate[J]. Chinese Journal of Environmental Engineering, 2016, 10(11): 6127-6132. DOI:10.12030/j.cjee.201506020 |
| [10] | APHA. Standard methods for the examination of water and wastewater (21st ed.)[M]. Washington, DC: American Public Health Association, American Water Works Association, Water Environment Federation, 2005. |
| [11] |
黄佳路, 王小龙, 高大文. 常温下厌氧氨氧化污泥的储存及活性恢复[J]. 环境科学, 2017, 38(11): 4687-4695. Huang J L, Wang X L, Gao D W. Storage and reactivation of anaerobic ammonium oxidation (ANAMMOX) sludge at room temperature[J]. Environmental Science, 2017, 38(11): 4687-4695. |
| [12] |
徐峥勇.基于亚硝化、厌氧氨氧化与反硝化的脱氮耦合工艺及其控制策略研究[D].长沙: 湖南大学, 2011. Xu Z Y. Study on control strategies of partial nitrification, anaerobic ammonium oxidation and denitrification coupling process[D]. Changsha: Hunan University, 2011. http://www.wanfangdata.com.cn/details/detail.do?_type=degree&id=Y2065281 |
| [13] |
魏思佳, 于德爽, 李津, 等. 厌氧氨氧化与反硝化耦合脱氮除碳研究Ⅰ:COD/NH4+-N对耦合反应的影响[J]. 中国环境科学, 2016, 36(3): 759-767. Wei S J, Yu D S, Li J, et al. Simultaneous carbon and nitrogen removal by anaerobic ammonium oxidation and denitrification Ⅰ:effect of COD/NH4+-N on coupled reaction[J]. China Environmental Science, 2016, 36(3): 759-767. DOI:10.3969/j.issn.1000-6923.2016.03.019 |
| [14] |
陈重军, 张海芹, 汪瑶琪, 等. 基于高通量测序的ABR厌氧氨氧化反应器各隔室细菌群落特征分析[J]. 环境科学, 2016, 37(7): 2652-2658. Chen C J, Zhang H Q, Wang Y Q, et al. Characteristics of microbial community in each compartment of ABR ANAMMOX reactor based on high-throughput sequencing[J]. Environmental Science, 2016, 37(7): 2652-2658. |
| [15] |
刘晓萍.厌氧氨氧化菌的富集及其与反硝化菌协同脱氮研究[D].青岛: 青岛科技大学, 2016. Liu X P. Enrichment of anaerobic ammonium oxidation bacteria and coupled with denitrification bacteria for nitrogen removal[D]. Qingdao: Qingdao University of Science & Technology, 2016. http://cdmd.cnki.com.cn/Article/CDMD-10426-1016719892.htm |
| [16] | Kumar M, Lin J G. Co-existence of ANAMMOX and denitrification for simultaneous nitrogen and carbon removal-Strategies and issues[J]. Journal of Hazardous Materials, 2010, 178(1-3): 1-9. DOI:10.1016/j.jhazmat.2010.01.077 |
| [17] |
安芳娇, 黄剑明, 黄利, 等. 基质比对厌氧氨氧化耦合反硝化脱氮除碳的影响[J]. 环境科学, 2018, 39(11): 5058-5064. An F J, Huang J M, Huang L, et al. Effect of substrate ratio on removal of nitrogen and carbon using anaerobic ammonium oxidation and denitrification[J]. Environmental Science, 2018, 39(11): 5058-5064. |
| [18] |
汪瑶琪, 张敏, 姜滢, 等. 厌氧氨氧化启动过程及微生物群落结构特征[J]. 环境科学, 2017, 38(12): 5184-5191. Wang Y Q, Zhang M, Jiang Y, et al. Start-up and Characteristics of the microbial community structure of ANAMMOX[J]. Environmental Science, 2017, 38(12): 5184-5191. |
| [19] | Chen C J, Huang X X, Lei C X, et al. Effect of organic matter strength on ANAMMOX for modified greenhouse turtle breeding wastewater treatment[J]. Bioresource Technology, 2013, 148: 172-179. DOI:10.1016/j.biortech.2013.08.132 |
| [20] | Kindaichi T, Yuri S, Ozaki N, et al. Ecophysiological role and function of uncultured Chloroflexi in an ANAMMOX reactor[J]. Water Science & Technology, 2012, 66(12): 2556-2561. |
| [21] | Miao Y, Liao R H, Zhang X X, et al. Metagenomic insights into Cr(VI) effect on microbial communities and functional genes of an expanded granular sludge bed reactor treating high-nitrate wastewater[J]. Water Research, 2015, 76: 43-52. DOI:10.1016/j.watres.2015.02.042 |
| [22] | Zhang Z, Liu S. Insight into the overconsumption of ammonium by ANAMMOX consortia under anaerobic conditions[J]. Journal of Applied Microbiology, 2014, 117(6): 1830-1838. DOI:10.1111/jam.12649 |
| [23] | Van Der Star W R L, Miclea A I, Van Dongen U G J M, et al. The membrane bioreactor:a novel tool to grow ANAMMOX bacteria as free cells[J]. Biotechnology and Bioengineering, 2008, 101(2): 286-294. DOI:10.1002/bit.21891 |
| [24] | Wang C, Liu S T, Xu X C, et al. Achieving mainstream nitrogen removal through simultaneous partial nitrification, ANAMMOX and denitrification process in an integrated fixed film activated sludge reactor[J]. Chemosphere, 2018, 203: 457-466. DOI:10.1016/j.chemosphere.2018.04.016 |
| [25] | Xia Y, Wang Y B, Wang Y, et al. Cellular adhesiveness and cellulolytic capacity in Anaerolineae revealed by omics-based genome interpretation[J]. Biotechnology for Biofuels, 2016, 9: 111. DOI:10.1186/s13068-016-0524-z |
| [26] | Deng Y, Guo X, Wang Y W, et al. Terrisporobacter petrolearius sp. nov., isolated from an oilfield petroleum reservoir[J]. International Journal of Systematic and Evolutionary Microbiology, 2015, 65(10): 3522-3526. DOI:10.1099/ijsem.0.000450 |
| [27] | Parshina S N, Strepis N, Aalvink S, et al. Trichococcus shcherbakoviae sp. nov., isolated from a laboratory-scale anaerobic EGSB bioreactor operated at low temperature[J]. International Journal of Systematic and Evolutionary Microbiology, 2019, 69(2): 529-534. DOI:10.1099/ijsem.0.003193 |
| [28] | Thomsen T R, Kong Y H, Nielsen P H. Ecophysiology of abundant denitrifying bacteria in activated sludge[J]. FEMS Microbiology Ecology, 2007, 60(3): 370-382. DOI:10.1111/j.1574-6941.2007.00309.x |
| [29] | Osaka T, Yoshie S, Tsuneda S, et al. Identification of acetate-or methanol-assimilating bacteria under nitrate-reducing conditions by stable-isotope probing[J]. Microbial Ecology, 2006, 52(2): 253-266. DOI:10.1007/s00248-006-9071-7 |
| [30] | Cao S B, Du R, Li B K, et al. High-throughput profiling of microbial community structures in an ANAMMOX-UASB reactor treating high-strength wastewater[J]. Applied Microbiology and Biotechnology, 2016, 100(14): 6457-6467. DOI:10.1007/s00253-016-7427-6 |
| [31] | Su J F, Ma M, Huang T L, et al. Characteristics of autotrophic and heterotrophic denitrification by the strain Pseudomonas sp. H117[J]. Geomicrobiology Journal, 2017, 34(1): 45-52. DOI:10.1080/01490451.2015.1137661 |
| [32] | Zhang Y H, Zhong F H, Xia S Q, et al. Autohydrogenotrophic denitrification of drinking water using a polyvinyl chloride hollow fiber membrane biofilm reactor[J]. Journal of Hazardous Materials, 2009, 170(1): 203-209. DOI:10.1016/j.jhazmat.2009.04.114 |
| [33] | Kong Q, He X, Feng Y, et al. Pollutant removal and microorganism evolution of activated sludge under ofloxacin selection pressure[J]. Bioresource Technology, 2017, 241: 849-856. DOI:10.1016/j.biortech.2017.06.019 |