 2019, Vol. 40
2019, Vol. 40


2. 哈尔滨工业大学城市水资源与水环境国家重点实验室, 哈尔滨 150090
2. State Key Laboratory of Urban Water Resource and Environment, Harbin Institute of Technology, Harbin 150090, China
短程硝化可以节约25%的能源与40%的碳源, 是低碳氮比废水脱氮除磷工艺中重要的工艺.目前短程硝化工艺已经成功应用于污泥消化液、垃圾渗滤液等高氨氮污废水的处理, 但是低氨氮生活污水短程硝化的运行尚存在问题, 如何取得稳定的亚硝酸盐积累仍是实验中的难点.强化生物除磷工艺与亚硝化工艺具有一定协同性, A/O不断交替地运行方式以及较长的厌氧段不利于NOB的增殖, 所以在同一个反应器内实现除磷与亚硝化是技术可行的, 值得研究与探索.已有研究大多针对絮状污泥[1, 2], 如果将颗粒污泥工艺与除磷亚硝化结合起来, 则系统具有微生物含量高, 抗冲击能力强, 避免二次释磷的优点, 对处理实际污水有极大意义.
泥龄是除磷亚硝化颗粒污泥系统的重要参数, 既要通过排出富磷污泥达到除磷目的, 又要考虑到硝化菌较长的世代时间.泥龄的选择还和温度密切相关, 温度会影响系统的处理性能, 合适的泥龄既可保证功能菌群的数量, 最大限度地维持系统生物活性, 也降低了污泥处置费用.绝大部分微生物适宜生长的温度区间为20~30℃, 但在寒冷地区或冬季温度明显低于此水平, 故确保系统在低温下稳定运行尤为重要.根据秋冬季室温变化特点, 本文探究了不同泥龄(20、30和40 d)对中低温度下(14~21℃)除磷亚硝化颗粒污泥的影响.运行过程中对系统处理性能、颗粒物理性能变化、典型周期等进行测量和分析, 以期为工程实际应用提供参考.
1 材料与方法 1.1 实验装置与运行方法本实验采用有机玻璃制成的SBR反应器, 有效容积为6 L, 换水比为2/3.每天运行4个周期, 每周期进水5 min, 厌氧2 h, 好氧3 h, 沉淀3 min, 排水5 min, 其余时间闲置, 每周期末排出一定体积的泥水混合物.阶段Ⅲ第98~109 d不进水, 颗粒经历12 d的厌氧饥饿期, 期间每8 h曝气10 min, 去除厌氧环境产生的H2S气体.根据泥龄的不同划分实验阶段, 具体参数见表 1.
|
|
表 1 反应器运行工况 Table 1 Operational conditions of the reactor |
1.2 接种污泥与实验用水
接种污泥是以去除P和COD为主的除磷颗粒污泥, 污泥浓度为3 300 mg ·L-1, 实验用水取自北京某大学家属区化粪池.具体水质指标如表 2.
|
|
表 2 生活污水水质情况 Table 2 Water quality of domestic sewage |
1.3 分析项目与检测方法
NH4+-N采用纳氏试剂光度法; NO2--N采用N-(1-萘基)-乙二胺光度法; NO3--N采用麝香草酚紫外分光光度法; COD和TP的测定采用5B-3B型COD多参数快速测定仪, 每两天对上述参数进行测量; pH、DO和温度的测定均采用WTW-pH/Oxi 340i便携式多参数测定仪监测. MLSS、MLVSS、SV30和SVI等指标均采用国家规定的标准方法[3].颗粒的粒径采用Mastersize 2000激光粒度仪测定.
1.4 衰减实验速率测定AOB和NOB的活性用比氨氧化速率(SAOR)和比亚硝酸盐氧化速率(SNOR)表示, PAOs活性则依靠衰减过程中最大比释磷速率(SPRR)来确定.测试方法如下:从洗去杂质的反应器中取1.5 L泥水混合物, 平均分成3份, 用来测量SAOR、SNOR和SPRR.测SAOR、SNOR时, 分别加入NH4Cl、NaNO2, 使NH4+和NO2-初始浓度为50mg ·L-1, 调节pH为7.5±0.1, DO控制在4.0mg ·L-1.测SPRR时加入丙酸钠, 使COD浓度为200mg ·L-1, 保持厌氧环境, 每30 min取样, 直至浓度不再变化.采用Origin 9.0对NH4+-N、NO2--N和P浓度随时间的变化进行拟合, 拟合直线的斜率分别为SAOR、SNOR和SPRR.衰减速率的计算公式如下:
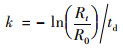
|
式中, R0和Rt分别表示饥饿衰减前后的SAOR、SNOR和SPRR; td表示饥饿时间(d).
2 结果与讨论 2.1 阶段Ⅰ和Ⅱ除磷亚硝化颗粒污染物处理性能 2.1.1 COD及TP去除性能图 1为阶段Ⅰ和阶段Ⅱ运行过程中系统对COD和P的去除情况.阶段Ⅰ接种的除磷颗粒污泥, 具有良好的去除COD和P的性能, 为了加快AOB的富集, 控制泥龄为30 d.延长的污泥龄并未对P的去除产生不利影响, 出水P浓度一直小于0.50mg ·L-1, 去除率保持在90%以上, 除磷效果达到稳定.厌氧段聚磷菌吸收VFA储存在体内, 绝大部分COD在厌氧段被吸收, 以保证后续好氧段PHAs氧化分解释放的能量充分摄取水中的P.阶段Ⅰ接近90%的COD在厌氧段去除, 出水浓度小于50mg ·L-1.

|
图 1 阶段Ⅰ和Ⅱ运行过程中COD和TP变化 Fig. 1 Variation of COD and TP concentration during the process of phaseⅠand phaseⅡ |
阶段Ⅱ随着温度的降低, 调整泥龄为40 d, 第65 d之后出水P浓度呈升高趋势, 阶段末去除率跌至65.68%. Brdjanovic等[4]发现过高的SRT导致了内源呼吸, 氧转移的需求量降低, 过量曝气降低了生物除磷的效率. 40d的长泥龄下, 污泥处于老化状态, 部分聚磷菌吸附值达到饱和, 对磷的吸附能力有限, 聚磷菌活性下降影响了吸磷与释磷能力.聚磷作为重要的能量转换器, 释磷量降低影响了对VFA的摄取, 厌氧段COD的去除率下降至81.72%.好氧段过多的COD引起了其他异养菌的增殖, 故出水COD浓度基本不变.由此40 d的泥龄并未影响COD的去除, 但严重破坏了除磷性能.
2.1.2 氮元素转化变化图 2为阶段Ⅰ和阶段Ⅱ氮元素变化情况.运行初期出水氨氮浓度很高, 随着时间延长, 出水氨氮降低至10mg ·L-1左右.本阶段末亚硝酸盐积累量升高至8mg ·L-1, 与此同时NAR从40%逐渐增加至90%, NOB的活性被充分抑制.分析原因如下:A/O除磷运行模式下, 厌氧段相对较长, AOB的“饱食饥饿”特性使其更从容地面对环境变化; 颗粒污泥特殊的结构存在氧气的传质限制, 并且AOB对溶解氧的亲和力要大于NOB, 故AOB占有优先利用溶解氧的外层, NOB紧靠AOB生长在次外层或外层[5]. NOB的生长需要利用AOB代谢产生的亚硝酸盐, 亚硝酸盐的氧化过程滞后于氨氧化过程.上述条件下, NOB增殖速度小于AOB, 在合适的泥龄下, 通过不断排泥将NOB淘洗出去, 保持亚硝化的稳定.

|
图 2 阶段Ⅰ、Ⅱ运行过程中NH4+-N、NO2--N及NO3--N变化 Fig. 2 Variation in NH4+-N, NO2--N, and NO3--N concentration during the process of phase Ⅰ and phase Ⅱ |
阶段Ⅱ增加泥龄为40 d, 运行至第69 d时亚硝化被破坏, NAR大幅下降, 到第87 d时只有22.4%.稳定的亚硝化作用和良好的除磷性能密不可分, 40 d的泥龄限制了聚磷菌的活性, 从而使聚磷菌竞争溶解氧的能力下降, 为硝化作用提供了相对充足的氧气.此外, 本阶段平均温度为15℃, 硝化作用容易受温度影响, AOB活性下降, 对氧的利用率降低导致氧气的穿透深度增加.有研究显示[6], 低温下缩小了AOB与NOB生长速率间的差异.以上诸多因素破坏了AOB在硝化菌群中的优势地位, 导致NOB增殖.第88 d将曝气量降低, NAR略微上升, 但是依然以全程硝化为主.
2.1.3 典型周期分析分别在阶段Ⅰ第43 d和阶段Ⅱ第87 d测量一个周期内COD、TP、三氮、pH及DO变化情况, 进行对比分析, 明晰处理效果恶化的原因, 为后续恢复系统正常运行采取有效的措施..
周期实验结果如图 3所示, 两个阶段绝大部分COD在厌氧段去除, COD浓度在前60 min迅速下降, 出水浓度约为30mg ·L-1, 可见泥龄对COD去除无影响.第43 d和87 d的释磷量分别为40.25 mg ·L-1和36.12 mg ·L-1, 但是好氧吸磷有显著不同.第43 d前60 min内平均吸磷速率为33.53 mg ·(h ·L)-1, 明显高于后者的21.05 mg ·(h ·L)-1, 并且后者吸磷不彻底, 周期末仍有2.1mg ·L-1的P残留.徐伟峰等[7]的研究发现随着SRT的延长, 聚磷菌消耗单位PHAs的吸磷量与合成单位PHAs所需释磷量的比值呈下降趋势.即在40 d的长泥龄下, PHA的生成量与消耗量不平衡, 不足以让释放的P被全部吸收, 可认为发生了无效释磷.

|
图 3 典型周期内各参数变化 Fig. 3 Change diagram of each parameter in a typical period |
第43 d, 曝气90 min内NO2--N及NO3--N含量未出现明显增加, 随着反应进行, 亚硝酸积累量增加至8.52mg ·L-1. Pijuan等[8]的研究表明, FNA的存在会对聚磷菌的生长及代谢活动产生抑制作用, 如影响氧利用率, PHA氧化和糖原生成.当FNA浓度达到5×10-4 mg ·L-1, 50%的吸磷性能被抑制. FNA浓度随亚硝盐积累量和pH变化而变化, 从第190 min起FNA含量从9.4×10-5 mg ·L-1逐渐增加至6.2×10-4 mg ·L-1.由图 3可得吸磷作用优先于硝化过程, P浓度在210 min时仅剩3.2mg ·L-1, 且吸磷能力不会完全丧失, 故亚硝酸盐积累对吸磷影响不大.第87 d, 由于吸磷活性的下降, 氨氧化作用增强, 氨氧化率高于第43 d, 这和增长的污泥浓度有关.但是从180 min起开始有硝酸盐生成, 周期末达到9.2mg ·L-1.
pH变化趋势和生化反应有良好的相关性, 厌氧段释磷pH下降, 前60 min内释磷基本结束.好氧段初期在氧化有机物和吸磷的共同作用下pH上升, 随后增强的硝化过程消耗碱度致使pH下降, 所以pH曲线上的拐点可以定性表征硝化反应的强度.第43 d时, 在120~170 min内pH从7.27上升至7.90, 随后16 min内pH值基本不变, 此时溶解氧浓度低, 颗粒存在缺氧区, 这可能是吸磷与短程硝化反硝化共同作用的结果, 直到第187 min时pH开始下降, 最后值为7.58. pH会引起细胞膜电荷变化, 导致微生物细胞吸收营养物质能力改变.不同微生物有各自的适宜生长范围, AOB为7~8.2, NOB为6.5~7.5.由此, 吸磷过程中的pH变化范围也有利于亚硝化的稳定[9].第87 d, 由于较弱的吸磷过程, pH曲线上拐点出现的时间提前, 硝化反应进程加快.长泥龄下聚磷菌的耗氧速率下降, DO浓度在150 min时就达到4.5mg ·L-1, 周期结束时上升至6.22mg ·L-1.相比第43 d, 过高的溶解氧浓度使生成的NO2--N迅速转化成NO3--N, 整个过程中无明显NO2--N积累.
2.2 除磷亚硝化颗粒污泥活性的恢复及低温下稳定运行低铵浓度废水中NOB的抑制主要依赖于AOB和NOB之间的动力学竞争, 通常通过高游离氨、限氧策略(DO<1mg ·L-1)、曝气时间实时控制实现亚硝化过程.本实验中, 根据生活污水的水质和水量变化特征, pH在7~8之间, 若长期投加药剂势必会增加运行成本.其次, 限氧策略下低DO会抑制AOB活性, 使反应时间延长.曝气时间实时控制是根据pH曲线“氨谷”以及DO突跃的特征点的出现, 判断氨氧化结束, 从而及时停止曝气.但是从周期实验中可以看出氨氧化并不完全, pH曲线仍是下降趋势.并且亚硝酸盐氧化过程迅速完成, 不会明显滞后于氨氧化, 无法根据特征点将两个过程分开.由此当NOB活性较强时, 可以采取更为严苛的措施.有研究发现污泥经历饥饿后在恢复期会出现NO2--N积累[10, 11], 这为实现亚硝化提供一种新的思路.大多数实验均是在絮状污泥系统中开展的, 关于饥饿对颗粒污泥中亚硝化性能恢复的研究甚少.本实验探究了经过12 d的厌氧饥饿后, 系统处理性能变化情况.
2.2.1 饥饿期除磷亚硝化颗粒污泥活性变化分别在第0、3、6、9和12 d测量SAOR、SNOR和SPRR, 得到AOB、NOB和PAOs的活性变化情况如图 4所示.饥饿前, SNOR(以N/VSS计, 下同)为2.95mg ·(g ·h)-1, NOB的活性高于AOB.随后的饥饿期使两者的活性均有下降, 但变化快慢有所不同, NOB的活性衰减速率较快.饥饿期结束时, SAOR约为SNOR的1.06倍, 可见饥饿对NOB活性的抑制随着时间延长而加大.但饥饿时间过久, 会对除磷性能造成不利影响并且恢复期过长, 故在饥饿12 d后恢复进水.利用Origin拟合曲线, 得到AOB和NOB的衰减速率分别为0.030 d-1和0.042 d-1. AOB具有许多酶学和分子机制, 使得它能够在饥饿状态下降低维持自身的能量需求, 进入休眠期, 导致较低的衰减速率.并且期间AOB不会终止底物转化酶的活动, 一旦底物重新供给时就能迅速恢复活性[12].所以饥饿后出现NO2--N积累的原因不仅是衰减速率的差异, 还归功于AOB快速适应环境变化的能力.衰减系数与原水水质条件、菌群丰度、饥饿环境等密切相关, 已有报道指出AOB的衰减范围在厌氧条件下为0.006~0.114, NOB为0.007~0.159[13~15].本实验所得数值偏低, 这和本系统的泥龄与温度有关.温度越低, 活性下降越缓慢; 泥龄越长, 菌群会降低生长率, 适应饥饿能力越强[16].

|
图 4 厌氧饥饿期间PAOs、AOB和NOB活性变化 Fig. 4 Variations in the activity of PAOs, AOB, and NOB during the anaerobic starvation periods |
饥饿前PAOs的SPRR(以P/VSS计)为13.70mg ·(g ·h)-1, 饥饿期结束后活性下降至8.18mg ·(g ·h)-1, 拟合曲线得到PAOs衰减速率为0.044 d-1.此数值偏低, 可能由于较长泥龄与菌群系统不同所致, BNR系统中聚磷菌衰减率小于纯富集聚磷菌的系统[17, 18]. Lopez等[19]的研究表明, PAOs可在厌氧饥饿下消耗聚磷和糖原用于维持自身能量生成, 与好氧饥饿相比, 厌氧主要是利用一小部分代谢底物产生维持生命所需的能量, 发生的生物衰减量极低.
2.2.2 除磷亚硝化颗粒污染物处理性能变化图 5为阶段Ⅲ和阶段Ⅳ运行过程中COD、TP和三氮浓度变化情况.阶段Ⅲ刚开始恢复进水时, COD出水浓度几乎等于厌氧末浓度, 可见饥饿期使存储基质能力差的异养菌的活性大幅下降, 4 d的恢复后出水COD浓度小于50mg ·L-1.第110 d, 经历厌氧饥饿期后PAOs释磷浓度达到了48mg ·L-1, 但是吸磷能力未完全恢复, 3 d后出水P浓度趋于稳定.聚磷菌[20]是一种嗜冷菌, 在较低温下对VFA仍有较强的摄取能力.此阶段平均温度为15℃, 不会对除磷产生不利影响, 出水P浓度一直小于0.5mg ·L-1.

|
图 5 阶段Ⅲ、阶段Ⅳ中COD、TP及氮素变化 Fig. 5 Variation in COD, TP, and nitrogen elements concentration during the process Ⅲ and Ⅳ |
恢复进水后缩短泥龄为20 d, 9 d后NAR便恢复至80%.短期饥饿对AOB活性影响不大[12, 21], 甚至比异养菌能更快地恢复活性, 因为异养生物饥饿后再恢复时需改变其酶模式以使得能够使用初始底物, 基因调控与表达需要时间.饥饿期削弱了NOB相对AOB的活性, 且AOB有更优的环境适应能力及更快的生长速率, 加上排泥使NOB彻底地从系统中淘洗出去, 继而确立了AOB的种群优势.第139 d, 出水NH4+-N达到10.8mg ·L-1, 即此温度下维持20 d的泥龄不利于氨氮的去除.遂将泥龄增加至30 d, 出水NH4+-N逐渐降低, NO2--N含量增加. Guo等[22]发现随着温度从25℃降至15℃, 比氨氧化速率降低了1.5倍, 氧利用率降低.为防止过度曝气破坏亚硝化, 根据周期实验结果(图 3)此阶段控制曝气量为4L ·(h ·L)-1, 保证周期末溶解氧浓度为不超过4mg ·L-1.有研究表明由于PAOs可以在有氧和缺氧条件下生长[23], 因此PAOs将存在于整个颗粒中, 且它们的性能将受到氧渗透深度变化的不显著影响, 故即使曝气量降低后, 除磷性能良好.
2.3 不同阶段PAOs和AOB的比耗氧速率(SOUR)变化SOUR(以O2/VSS计)代表微生物的分解代谢活性, 分别在阶段Ⅰ、Ⅱ和Ⅳ运行后期中取出适量污泥测量PAOs及AOB的比耗氧速率, 消除污泥内源呼吸对测量的影响[24, 25], 结果如图 6所示.阶段Ⅰ中, 聚磷菌耗氧速率为35.87mg ·(g ·h)-1, 过长的泥龄使吸磷过程的SOUR下降至26.89mg ·(g ·h)-1.当温度从20℃下降到15℃时, 根据计算出PAOs和AOB温度依赖系数θ分别为1.023和1.071, 即AOB受温度影响更大.聚磷菌在15℃时仍有着较高的氧利用率, 以此维持着吸磷与亚硝化间的平衡, 这也是阶段Ⅳ与阶段Ⅰ相比, 温度降低亚硝化却仍然稳定的重要原因.阶段Ⅱ与阶段Ⅳ比, 虽然曝气量大, 但AOB的SOUR无明显增加, 故低温下适当降低曝气的方法是正确的, 此时溶解氧不是限制硝化反应速率的重要因素, 而对硝化类型产生了影响.

|
图 6 不同阶段PAOs和AOB的比耗氧速率变化 Fig. 6 Variations in the specific oxygen uptake rate of PAOs and AOB during different phases |
图 7为不同阶段颗粒形态的变化.阶段Ⅰ颗粒呈棕黄色, 具有规则的形状和清晰的边界.阶段Ⅱ颗粒颜色变浅, 出现少部分破碎颗粒, 在显微镜下可观察到颗粒边缘部分被残蚀, 并伴有丝状菌产生.这是由于长泥龄降低了聚磷菌的耗氧速率, 过度曝气使自身储存的PHA产生对吸磷没有积极作用的消耗浪费, 吸磷不充分继而造成后续释磷动力不足.长时间的恶性循环加之搅拌、曝气等水力剪切力作用下, 颗粒致密结构受到影响.饥饿期使颗粒上缺乏营养的丝状菌脱落, 阶段Ⅳ观察到颗粒结构有所恢复, 可能受到温度影响, 颗粒形状没有阶段Ⅰ规则.

|
图 7 不同阶段颗粒污泥的显微镜照片和数码照片 Fig. 7 Optical microscopy images and digital images of granulesduring different phases |
在40、97(饥饿前)、110(饥饿结束后)和175 d测量颗粒粒径, 得到粒径分布结果如图 8所示(d0.1、d0.5和d0.9分别表示粒径累计分布数达到10%、50%和90%时所对应的粒径).第40 d, 系统平均粒径为1 092 μm, 90%以上的颗粒粒径大于613 μm, 几乎没有小于420 μm的颗粒.第97 d颗粒平均粒径降低至935 μm, 长泥龄下, 颗粒过度消耗自身物质维持生命活动, 导致颗粒的结构和功能恶化.而大颗粒更容易受到影响, 粒径大于1000 μm颗粒所占的体积分数明显降低.经历饥饿期后, 部分破碎的颗粒使小粒径所占百分比增加, d10降至210 μm.粒径500~1 000 μm颗粒的体积分数下降幅度不大, 因为颗粒污泥含有大量EPS, EPS在厌氧饥饿期间维持颗粒的结构完整性方面起着至关重要的作用[26].泥龄恢复为30 d后, 加大排泥, 破碎颗粒被淘洗, 颗粒粒径有所增长, 175 d时90%的颗粒粒径大于533 μm, 平均达到1 057 μm.

|
图 8 第40、97、110和175 d粒径分布情况 Fig. 8 Particle size distribution of 40, 97, 110, and 175 d |
图 9是不同阶段MLSS、MLVSS、f及SVI的变化.阶段Ⅰ污泥浓度从3 300 mg ·L-1逐渐增长至4 150 mg ·L-1, 沉淀性能良好, SVI在35~37 mL ·g-1.随着泥龄增加到40 d, MLSS继续保持增长, 但是MLVSS增势明显缓慢.泥龄过长, 污泥处于老化状态, 无机质含量高, 生物活性降低, 第97 d, f值从阶段Ⅱ初的0.8降至0.73.丝状菌的增殖影响了颗粒的沉降性能, SVI值增加至55 mL ·g-1.饥饿期有污泥减量的效果, 加上沉降性能不好的颗粒被排出, 第112 d, MLSS降低至4 236 mg ·L-1, 但SVI值在逐渐恢复. 20 d的污泥龄下污泥浓度趋于稳定, 延长泥龄至30 d时, 污泥浓度有所增长, 受到基质和温度限制, 增长速度明显缓于阶段Ⅰ.低温使AOB活性下降, 和阶段Ⅰ相比, 维持同样的氨氮处理效果, 则需要更高的污泥量.

|
图 9 运行过程中SVI、MLSS、MLVSS及f的变化 Fig. 9 Variation in SVI, MLSS, MLVSS, and f during the procession of operation |
(1) 以实际生活污水为研究对象, 控制泥龄为30 d, 常温下完成了除磷颗粒污泥系统中AOB的富集.吸磷过程优先于亚硝化, 亚硝酸盐积累量 < 10mg ·L-1时, 吸磷性能未受影响. 15℃时延长泥龄至40 d, 氨转化和COD去除稳定, 但除磷受到影响, 颗粒致密结构被破坏, 过度曝气导致NOB大量增殖.故通过提高泥龄的方式弥补低温带来系统处理性能损失的同时, 亦应兼顾长泥龄对特定微生物活性的影响, 从而确定合适的温度-泥龄组合.
(2) 12d饥饿胁迫削弱了NOB相对于AOB的活性, 联合排泥9 d后可建立AOB在硝化菌群的优势地位. AOB应对饥饿特殊的生理机制和聚磷菌较低的衰减量, 使它们在基质重新供给时, 能较快地恢复活性, 适时的饥饿对颗粒结构影响不大.
(3) 低温下AOB活性变化比聚磷菌更敏感, 20 d的泥龄不利于氨氮去除.溶解氧此时不是限制硝化反应速率的显著因素, 而是决定了硝化类型, 故低温下应适当降低曝气量.控制泥龄为30 d, 协同调控曝气量4L ·(h ·L)-1, 以较高的生物质浓度保证了处理效果, 实现了低温下除磷亚硝化颗粒污泥系统的稳定运行.
| [1] |
马斌, 张树军, 王俊敏, 等. A/O工艺实现城市污水半亚硝化与生物除磷[J]. 哈尔滨工业大学学报, 2012, 44(2): 70-74. Ma B, Zhang S J, Wang J M, et al. Partial nitritation and biological phosphorus removal in A/O process treating municipal wastewater[J]. Journal of Harbin Institute of Technology, 2012, 44(2): 70-74. |
| [2] |
周元正, 李冬, 苏东霞, 等. 改良SBR工艺实现生活污水除磷与半亚硝化[J]. 中国环境科学, 2015, 35(5): 1351-1358. Zhou Y Z, Li D, Su D X, et al. Partial nitritation and phosphorus removal in improved SBR process treating domestic wastewater[J]. China Environmental Science, 2015, 35(5): 1351-1358. DOI:10.3969/j.issn.1000-6923.2015.05.009 |
| [3] | 国家环境保护局. 水和废水监测分析方法[M]. 第四版. 北京: 中国环境科学出版社, 2002: 100-124. |
| [4] | Brdjanovic D, Van Loosdrecht M C M, Hooijmans C M, et al. Temperature effects on physiology of biological phosphorus removal[J]. Journal of Environmental Engineering, 1997, 123(2): 144-153. |
| [5] |
杨洋, 左剑恶, 孙寓姣, 等. 好氧亚硝化颗粒污泥特性的研究[J]. 环境科学, 2007, 28(12): 2710-2715. Yang Y, Zuo J E, Sun Y J, et al. Characteristics of aerobic short-cut nitrification granular sludge[J]. Environmental Science, 2007, 28(12): 2710-2715. DOI:10.3321/j.issn:0250-3301.2007.12.008 |
| [6] |
傅金祥, 王颖, 孙铁珩, 等. 低温条件下SBR系统中短程硝化研究[J]. 水处理技术, 2008, 34(7): 29-32. Fu J X, Wang Y, Sun T H, et al. Study on shortcut nitrification in SBR system at low temperature[J]. Technology of Water Treatment, 2008, 34(7): 29-32. |
| [7] |
徐伟锋, 顾国维, 陈银广. SRT对A2/O工艺脱氮除磷的影响[J]. 水处理技术, 2007, 33(9): 68-71. Xu W F, Gu G W, Chen Y G. Research on effect of SRT on A/A/O biological nutrient removal and simulation[J]. Technology of Water Treatment, 2007, 33(9): 68-71. DOI:10.3969/j.issn.1000-3770.2007.09.017 |
| [8] | Pijuan M, Ye L, Yuan Z G. Free nitrous acid inhibition on the aerobic metabolism of poly-phosphate accumulating organisms[J]. Water Research, 2010, 44(20): 6063-6072. DOI:10.1016/j.watres.2010.07.075 |
| [9] | Zhang J H, Zhang Q, Li X Y, et al. 2017. Rapid start-up of partial nitritation and simultaneously phosphorus removal (PNSPR) granular sludge reactor treating low-strength domestic sewage[J]. Bioresource Technology, 2017, 243: 660-666. DOI:10.1016/j.biortech.2017.06.129 |
| [10] | Morgenroth E, Obermayer A, Arnold E, et al. Effect of long-term idle periods on the performance of sequencing batch reactors[J]. Water Science & Technology, 2000, 41(1): 105-113. |
| [11] | Elawwad A, Sandner H, Kappelmeyer U, et al. Long-term starvation and subsequent recovery of nitrifiers in aerated submerged fixed-bed biofilm reactors[J]. Environmental Technology, 2013, 34(8): 945-959. DOI:10.1080/09593330.2012.722758 |
| [12] | Bollmann A, Schmidt I, Saunders A M, et al. Influence of starvation on potential ammonia-oxidizing activity and amoA mRNA levels of Nitrosospira briensis[J]. Applied and Environmental Microbiology, 2005, 71(3): 1276-1282. DOI:10.1128/AEM.71.3.1276-1282.2005 |
| [13] | Pijuan M, Werner U, Yuan Z G. Effect of long term anaerobic and intermittent anaerobic/aerobic starvation on aerobic granules[J]. Water Research, 2009, 43(14): 3622-3632. DOI:10.1016/j.watres.2009.05.007 |
| [14] | Geets J, Boon N, Verstraete W. Strategies of aerobic ammonia-oxidizing bacteria for coping with nutrient and oxygen fluctuations[J]. FEMS Microbiology Ecology, 2006, 58(1): 1-13. |
| [15] |
陈佼, 陆一新, 张建强, 等. 饥饿对人工快速渗滤系统硝化性能的影响[J]. 环境科学学报, 2018, 38(12): 4703-4712. Chen J, Lu Y X, Zhang J Q, et al. Impact of starvation on the nitrification performance of constructed rapid infiltration systems[J]. Acta Scientiae Circumstantiae, 2018, 38(12): 4703-4712. |
| [16] | Salem S, Moussa M S, Van Loosdrecht M C M. Determination of the decay rate of nitrifying bacteria[J]. Biotechnology and Bioengineering, 2006, 94(2): 252-262. DOI:10.1002/bit.20822 |
| [17] |
苗志加, 薛桂松, 翁冬晨, 等. 不同温度及厌氧/好氧运行条件对聚磷菌衰减特性的影响[J]. 土木建筑与环境工程, 2013, 35(2): 113-117, 123. Miao Z J, Xue J S, Weng D C, et al. Effect of different temperature and anaerobic/aerobic conditions on the decay characteristics of PAOs[J]. Journal of Civil, Architectural & Environmental Engineering, 2013, 35(2): 113-117, 123. |
| [18] |
王啟林, 郝晓地, 曹亚莉. 聚磷菌富集实验及其内源特征探究[J]. 环境科学学报, 2010, 30(12): 2405-2413. Wang Q L, Hao X D, Cao Y L. Enriched experiment and endogenous characteristics of polyphosphate-accumulating organisms (PAOs)[J]. Acta Scientiae Circumstantiae, 2010, 30(12): 2405-2413. |
| [19] | Lopez C, Pons M N, Morgenroth E. Endogenous processes during long-term starvation in activated sludge performing enhanced biological phosphorus removal[J]. Water Research, 2006, 40(8): 1519-1530. DOI:10.1016/j.watres.2006.01.040 |
| [20] | Helmer C, Kunst S. Low temperature effects on phosphorus release and uptake by microorganisms in EBPR plants[J]. Water Science & Technology, 1998, 37(4-5): 531-539. |
| [21] | Liu W L, Yang Q, Ma B, et al. Rapid achievement of nitritation using aerobic starvation[J]. Environmental Science & Technology, 2017, 51(7): 4001-4008. |
| [22] | Guo J H, Peng Y Z, Huang H J, et al. Short-and long-term effects of temperature on partial nitrification in a sequencing batch reactor treating domestic wastewater[J]. Journal of Hazardous Materials, 2010, 179(1-3): 471-479. DOI:10.1016/j.jhazmat.2010.03.027 |
| [23] | De Kreuk M K, Pronk M, Van Loosdrecht M C M. Formation of aerobic granules and conversion processes in an aerobic granular sludge reactor at moderate and low temperatures[J]. Water Research, 2005, 39(18): 4476-4484. DOI:10.1016/j.watres.2005.08.031 |
| [24] |
郝晓地, 朱景义, 曹秀芹, 等. 硝化细菌AOB与NOB衰减速率实验测定[J]. 环境科学学报, 2008, 28(12): 2499-2502. Hao X D, Zhu J Y, Cao X Q, et al. Experimental determination of the decay rate of nitrifying bacteria:AOB and NOB[J]. Acta Scientiae Circumstantiae, 2008, 28(12): 2499-2502. DOI:10.3321/j.issn:0253-2468.2008.12.015 |
| [25] |
肖剑波, 卢培利, 曾善文, 等. 基于氧利用速率表征好氧吸磷过程特性[J]. 环境工程学报, 2016, 10(10): 5479-5484. Xiao J B, Lu P L, Zeng S W, et al. Characteristic of aerobic phosphorus uptake process based on OUR[J]. Chinese Journal of Environmental Engineering, 2016, 10(10): 5479-5484. DOI:10.12030/j.cjee.201505081 |
| [26] | He Q L, Zhang W, Zhang S L, et al. Performance and microbial population dynamics during stable operation and reactivation after extended idle conditions in an aerobic granular sequencing batch reactor[J]. Bioresource Technology, 2017, 238: 116-121. DOI:10.1016/j.biortech.2017.03.181 |