 2019, Vol. 40
2019, Vol. 40



硒是维持人体和动物健康的必需微量营养元素.全国营养调查发现, 我国366个县的1.05亿人口因缺硒而出现了人体免疫力下降等健康问题[1], 严重缺硒时还会引发癌症、大骨节病、克山病等[2~4].通过饮食摄入硒是人体补硒的一个重要途径[5, 6].有研究发现, 植物性硒源的吸收利用率及安全性均高于动物性硒源[7].然而我国土壤缺硒严重, 可被植物吸收的硒含量很少, 因此人们通过向土壤中施加硒肥来提高农作物可食部位的硒含量, 进而达到人体补硒的目的[8].
硒在土壤环境中的存在形态主要有硒酸盐、亚硒酸盐、有机硒、元素态硒和硒化物[9].硒的形态不同, 植物对其的吸收利用率也不同, 因此植物根系对不同形态硒的吸收转化和向可食部位转运的能力直接决定了植物的富硒效果[10, 11].目前, 无机硒肥的应用十分广泛, 但不论是向土壤中基施[12]还是采用叶面喷施[13], 植物对硒的利用率都比较低.这就亟需寻找一种植物可高效利用的新型硒肥, 以推进富硒农产品的开发.
纳米硒(SeNPs)是一种新型纳米材料, 在医学上作为抗氧化剂、抗癌药物等已经得到了广泛应用[14~16].已有研究表明, SeNPs比无机硒或有机硒更能被生物有效利用[17~19], 且表现出低毒性[20]. Gao等[21]认为SeNPs也可用于制造硒肥来提高作物中的硒含量.目前, 硒肥的研究主要集中在亚硒酸盐和硒酸盐, 植物对SeNPs的吸收机制尚不清楚[22].基于此, 本文利用水培试验, 研究小麦和水稻两种植物对不同粒径SeNPs和在不同pH条件下对硒的吸收与转运情况, 通过探究植物吸收SeNPs的主要机制, 以期为富硒农产品的开发和对人体进行科学有效地补硒提供理论依据.
1 材料与方法 1.1 SeNPs的制备根据Lin等[23]的方法, 利用亚硒酸钠(Na2SeO3)和硫代硫酸钠(Na2S2O3)化学反应合成SeNPs.通过在不同离心力下分离和浓缩, 得到不同粒径(50、100和150 nm)的SeNPs, 悬浮于去离子水中于4℃保存备用.
1.2 植物培养试验地点是中国农业大学西校区资环温室, 植物生长在25℃/14 h光照和20℃/10 h黑暗条件下、相对湿度为60%~70%、光照强度为240~350 μmol ·(m2 ·s)-1.种子发芽前先浮选去掉瘪粒, 用30% H2O2消毒15 min, 洗净后在饱和CaSO4中浸泡过夜.待种子吸涨后, 移至装有蛭石的育苗盘中发芽.当叶子完全展开时, 挑选大小一致的幼苗洗净并移至营养液培养.
供试小麦(Triticum aestivum L.)品种为农大211, 采用1/5 Hoagland培养液培养, 组分包括:1.0 mmol ·L-1 KNO3、1.0 mmol ·L-1Ca(NO3)2 ·4H2O、0.5 mmol ·L-1 MgSO4 ·7H2O、0.1 mmol ·L-1 KH2PO4、1.0 μmol ·L-1 MnSO4 ·H2O、3 μmol ·L-1 H3BO3、1 μmol ·L-1 (NH4)6Mo7O24 ·5H2O、1 μmol ·L-1 ZnSO4 ·7H2O、0.2 μmol ·L-1 CuSO4 ·5H2O、60 μmol ·L-1 Fe(Ⅲ)-EDTA, 用MES(2 mmol ·L-1)缓冲液保持营养液的pH值在6.0左右(1 mmol ·L-1 KOH和HCl调节pH), 营养液每3~4 d换一次.
供试水稻(Oryza sativa L.)品种为准两优608, 采用1/2 Kimura培养液培养, 组分包括:0.091 mmol ·L-1 KNO3、0.183 mmol ·L-1 Ca(NO3)2 ·4H2O、0.1 mmol ·L-1 KH2PO4、0.274 mmol ·L-1 MgSO4 ·7H2O、0.183 mmol ·L-1 (NH4)2SO4、1 μmol ·L-1 MnSO4 ·H2O、3 μmol ·L-1 H3BO3、1 μmol ·L-1 (NH4)6Mo7O24 ·5H2O、1 μmol ·L-1 ZnSO4 ·7H2O、0.2 μmol ·L-1 CuSO4 ·5H2O、60 μmol ·L-1 Fe(Ⅲ)-EDTA, 用MES(2 mmol ·L-1)缓冲液保持营养液的pH值在5.5左右(1 mmol ·L-1 KOH和HCl调节pH), 营养液每3~4 d换一次.
1.3 试验设置 1.3.1 植物对不同粒径SeNPs的吸收和转运植物幼苗在营养液中培养4周后, 挑选大小一致的幼苗, 分别转移至含有50、100和150 nm SeNPs(5 μmol ·L-1)营养液的2.5 L塑料盆中进行处理.小麦和水稻处理液的pH用浓度为2 mmol ·L-1的MES缓冲液分别保持为6.0和5.5.每盆放置4株植物, 每个处理设3个重复, 分别在处理24 h和72 h后收获.
1.3.2 pH值对植物吸收转运硒的影响植物幼苗收获后, 转移到含SeNPs(5 μmol ·L-1, 50 nm)营养液的2.5 L塑料盆中进行处理, 并设置同浓度的亚硒酸盐为对照.用浓度为2 mmol ·L-1的MES缓冲液分别将小麦处理液的pH保持为4、6和8, 水稻处理液的pH保持为3.5、5.5和7.5.每盆放置4株植物, 每个处理设3个重复, 分别在处理24 h和72 h后收获.
1.4 样品硒含量的测定将收获后的植物用去离子水冲洗根部, 并置于200 mL预冷的CaSO4(1 mmol ·L-1)溶液中浸泡15 min, 然后植株用去离子水清洗3次, 吸水纸拭干后分离其根部和地上部, 记录鲜重.在烘箱中于105℃杀青30 min后在75℃烘干至恒重.样品称重后用磨样机粉碎, 置于信封袋中保存待测.
称取0.250 0~0.251 0 g干燥植物样品于消煮管中, 加入8 mL硝酸(优级纯)冷消化过夜.次日微波消解(MARS-5, CEM, 美国), 消解结束后冷却定容至50 mL容量瓶中, 并过滤到15 mL离心管中保存待测.
吸取4 mL消解液于15 mL离心管中, 加入1 mL 6 mmol ·L-1盐酸(优级纯), 经100℃水浴2 h, 将六价硒还原为四价硒后, 使用原子荧光光度计(AFS-8220, 北京吉天, 中国)测定溶液中的硒含量.样品消解、测定过程中以生物成分分析标准物质GBW10049(GSB-27)含硒为(0.069±0.009)mg ·kg-1的大葱和空白样品进行质量控制.使用的硒标准贮备液(100 mg ·L-1)购于国家环保总局标准样品研究所.
1.5 数据处理植物根系中硒的总量、地上部中硒的总量、植株中硒的总量、硒的转运系数(TF)以及硒在地上部所占的质量分数(Se地上部, %)的计算(根系、地上部的生物量以干重计)采用公式(1)~(5):
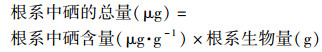
|
(1) |

|
(2) |
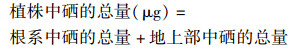
|
(3) |

|
(4) |

|
(5) |
采用Excel进行数据处理, 试验数据表示为3次重复的平均值±标准误差.使用SAS 9.3进行方差分析(多重比较采用Duncan法), 图采用Sigmaplot 12.5绘制.
2 结果与分析 2.1 植物对不同粒径SeNPs的吸收和转运 2.1.1 小麦和水稻对不同粒径SeNPs的吸收由图 1可知, 小麦根部和地上部中的硒含量随粒径的变化呈现不同的趋势.小麦根部硒含量在150 nm SeNPs处理下最高.当处理时间为24 h和72 h时, 150 nm SeNPs处理与50 nm和100 nm处理相比, 小麦根系中硒含量分别增加了10.00%、16.03%和14.21%、29.42%, 但差异不显著.然而, 小麦地上部硒含量在50 nm SeNPs处理下达到最大值, 处理24 h时, 与100 nm和150 nm处理相比, 硒含量分别显著增加了2.50倍和1.95倍(P < 0.05);处理72 h时, 50 nm SeNPs处理下的地上部硒含量分别比100 nm和150 nm处理增加了61.37%和47.16%, 但差异不显著.

|
不同字母表示处理间差异显著, P < 0.05, 下同 图 1 不同时间不同粒径SeNPs处理的小麦根系和地上部的硒含量 Fig. 1 Se content in wheat roots and shoots treated with differently sized SeNPs for different amounts of time |
图 2是水稻幼苗经过不同粒径SeNPs处理24 h和72 h后对硒的吸收情况.结果表明, 当处理时间为24 h时, 根系中的硒含量随SeNPs粒径的增大而逐渐减少, 50 nm SeNPs处理下根系中的硒含量比100 nm和150 nm SeNPs处理分别增加了11.18%和41.81%;同样, 50 nm SeNPs处理下水稻地上部的硒含量分别显著高于100 nm和150 nm SeNPs处理2.42倍和11.83倍(P < 0.05).当处理时间为72 h时, 随着SeNPs的粒径从50 nm增加到150 nm, 根系中的硒含量呈现先增加后减少的趋势, 但差异并不显著; 而地上部的硒含量随粒径的增大呈现逐渐减少的趋势, 50 nm SeNPs处理下地上部的硒含量显著高于100 nm和150 nm处理75.30%和91.86%(P < 0.05).

|
图 2 不同时间不同粒径SeNPs处理的水稻根系和地上部的硒含量 Fig. 2 Se content in rice roots and shoots treated with differently sized SeNPs for different amounts of time |
如表 1所示, 不同粒径SeNPs处理下, 硒在小麦和水稻幼苗体内的转运系数以及地上部分配比均随处理时间的增加而呈现增大的趋势.当处理时间为24 h时, 小麦幼苗在50 nm SeNPs处理下硒的转运系数均显著高于100 nm和150 nm处理2.38倍(P < 0.05), 且该处理下小麦根系吸收的硒被转运到地上部的比例最大, 约占6.00%, 显著高于其他两种处理(P < 0.05).水稻幼苗体内的硒转运规律与小麦幼苗基本一致, 随着SeNPs粒径的增加, 硒的转运系数和地上部硒的分配比均呈现逐渐减小的趋势.
|
|
表 1 不同粒径SeNPs在不同时间对小麦和水稻幼苗转运硒的影响1) Table 1 Effects of differently sized SeNPs on Se transport in wheat and rice seedlings after different periods of time |
当处理时间为72 h时, 随着SeNPs粒径的增加, 小麦体内硒的转运系数和地上部硒的分配比均逐渐减小, 50 nm SeNPs处理高于100 nm和150 nm处理, 但差异均不显著; 然而在水稻体内, 则呈现先减小后增加的趋势, 并且50 nm SeNPs处理下硒的转移系数和地上部分配比均显著高于100 nm和150 nm处理(P < 0.05).
2.2 pH值对植物吸收转运硒的影响 2.2.1 pH值对小麦、水稻吸收硒的影响图 3是不同时间处理下, pH对小麦根系吸收硒的影响情况.结果表明, 当处理时间为24 h时, 随着pH值的增大, 小麦根系对SeNPs的吸收量呈现先增大后减小的趋势, 而亚硒酸盐处理下, 硒的吸收量呈现逐渐减小的趋势.此外, 在各pH处理下, SeNPs在小麦根系中的吸收量均高于亚硒酸盐, 其中, 当pH为6时, 二者差异显著.当处理时间为72 h时, 小麦对SeNPs和亚硒酸盐的吸收量均随pH的增加而减小, 并且亚硒酸盐的吸收量始终高于SeNPs, 但差异均不显著.

|
图 3 不同pH条件和处理时间小麦根部对不同形态硒的吸收速率 Fig. 3 Uptake rate of Se by different species in wheat roots under different pH conditions after different periods of time |
图 4是不同时间处理下, pH对水稻根系吸收硒的影响.结果表明, 当处理时间为24 h时, 随着pH值的增大, 水稻根系对SeNPs的吸收量呈现先增大后减小的趋势, 而亚硒酸盐的吸收量呈现逐渐减少的趋势.同时, 在各pH处理下, 水稻根系对亚硒酸盐的吸收量均高于SeNPs.其中, 当pH为3.5和5.5时, 亚硒酸盐的吸收量显著高于SeNPs 1.42倍和0.53倍(P < 0.05), 而pH为7.5时, 水稻对两者的吸收量无显著差异.当处理时间为72 h时, 水稻对两种形态硒的吸收量均随pH的增加呈现先增大后减小的趋势.其中, pH为3.5和5.5时, 亚硒酸盐在水稻体内的吸收量显著高于SeNPs 1.19倍和0.75倍(P < 0.05), 而pH为7.5时, 二者无显著差异.

|
图 4 不同pH条件和处理时间水稻根部对不同形态硒的吸收速率 Fig. 4 Uptake rate of Se with different species in rice roots under different pH conditions after different periods of time |
小麦和水稻幼苗在不同时间和pH处理下, 对硒的转运情况如表 2所示.结果表明, 当添加SeNPs和亚硒酸盐处理时, 硒在小麦体内的转运系数及地上部分配比在相同pH条件下均随处理时间的增加而呈现增大的趋势.当处理时间为24 h时, SeNPs处理下, 小麦幼苗对硒的转运系数在pH为4时取得最大值, 比pH为8时显著增加了87.50%(P < 0.05);而亚硒酸盐处理下, 硒在小麦体内的转运系数随pH值的增大而增大.但是无论硒以何种形态添加, 不同pH条件对硒在小麦地上部的分配比均无显著影响.当处理时间为72 h时, 两种形态硒处理下, 硒在小麦体内的转运系数及其地上部的分配比均随pH值的增大而增大, 但无显著差异.
|
|
表 2 不同pH条件和处理时间对小麦和水稻幼苗转运硒的影响1) Table 2 Effects of pH on Se transport in wheat and rice seedlings after different periods of time |
对于水稻而言, 相同pH条件下添加SeNPs和亚硒酸盐处理, 硒在其体内的转运系数随处理时间的增加而逐渐减小, 但硒在水稻地上部中的分配比却随时间而增大. SeNPs在24 h和72 h处理下, 水稻对硒的转运系数和其地上部分配比均在pH为3.5时取得最大值.而亚硒酸盐在24 h处理下, 与其他pH条件相比, pH为5.5时水稻对硒的转运效果最佳.但用亚硒酸盐处理水稻72 h时, pH对硒在植株体内的转运系数无显著影响, 然而硒的地上部分配比在pH为3.5时取得最大值, 且显著高于pH为7.5处理(P < 0.05).
3 讨论 3.1 植物对不同粒径SeNPs的吸收和转运本试验结果表明, 3种粒径(50、100和150 nm)的SeNPs均能被小麦和水稻的根系吸收, 并且可以被转运至地上部.当处理时间为24 h时, 小麦根系在不同粒径SeNPs处理下的硒含量之间无显著差异.然而, 水稻经24 h处理后, 50 nm SeNPs处理下根系吸收的硒含量最多, 且显著高于150 nm SeNPs处理(P < 0.05), 这可能是因为不同种植物对SeNPs的吸收特性有所差异.当处理时间为72 h时, 小麦和水稻根系对3种粒径SeNPs的吸收均无显著性差异, 可能是由于此时两种植物根系对硒的吸收趋于稳定状态.因此, 在小麦和水稻体内, SeNPs粒径的不同对硒的吸收无显著影响.
此外, 50 nm SeNPs处理下, 硒在小麦和水稻体内更易从植物根系转运至地上部. 50 nm SeNPs处理硒的转运系数在24 h和72 h处理下, 均高于100 nm和150 nm处理, 并且各粒径硒的转运系数均随处理时间的增加而增大.与本试验结果不同的是, Hu等[24]用40、140和240 nm的SeNPs分别处理小麦(鲁原502)60 min, 发现40 nm SeNPs更易被小麦根系吸收, 且与另外两种处理差异显著(P < 0.05), 这可能是因为处理时间或供试小麦品种的不同, 导致小麦对不同粒径硒的吸收有所差异.同时, 其试验结果表明, 24 h处理下SeNPs的转运系数高于72 h处理, 但差异不显著.但是, 本研究与文献[24]都发现, 在所有处理下, 硒在小麦体内的转运系数均小于0.1.这说明大部分的硒被小麦吸收后会累积在根部, 向地上部转运的极少.另一方面, 植物对硒的转运特性也受硒形态的影响[25].有研究表明, 硒酸盐被植物根系吸收后, 可以保持原有的形态转移到地上部, 且转移的比例和速率较高[26, 27].而植物对亚硒酸盐的转运能力较低, 主要是由于其极易在植物根部被转化成有机硒等其他形态, 从而直接累积在根部, 不易向地上转移[27, 28].
3.2 pH值对植物吸收转运硒的影响本研究结果发现, 不同pH条件会影响硒在植物体内的吸收和转运.有研究表明, 土壤pH及氧化还原电位(Eh)是影响硒形态和植物有效性的主要因素[29].在酸性土壤中或还原条件下(7.5 < pe+pH < 15), 亚硒酸盐是硒的主要存在形态; 在碱性和氧化条件下(pe+pH>15), 硒酸盐是主要存在形态; 而元素态硒仅存在于低氧化还原条件下(pe+pH < 7.5), 在土壤中含量极少[30, 31].并且, SeNPs和亚硒酸盐在不同pH条件下的存在形态也可能有所不同.当处理时间为24 h时, SeNPs处理下小麦和水稻根系对硒的吸收量随pH的增加呈现先增大后减小的趋势[图 3(a)和图 4(a)], 而硒的转运系数却随pH的增加而减小(表 2), 这可能是因为SeNPs在植物的最适生长pH下更易被吸收, 但在低pH条件更易被转运.亚硒酸盐处理植物24 h时, 小麦和水稻吸收硒的量随pH的增加而逐渐减少[图 3(a)和图 4(a)], 该规律与王琪[32]和张联合等[33]的试验结果一致.不同pH条件下, 亚硒酸盐的存在形态不同[33], 其在低pH处理下可能更易被植物吸收.然而亚硒酸盐在两种植物中的转运情况却不相同, 小麦在pH为8时更易转运硒, 水稻在pH为5.5时对硒的转运能力更强.这可能是因为亚硒酸盐处理下, 硒在高pH环境更易被植物转运, 但水稻转运硒的能力受营养液pH的影响更大.当处理时间为72 h时, 添加SeNPs和亚硒酸盐处理, pH对硒在小麦体内的转运情况无显著影响, 但是水稻在较低pH环境下更易转运硒, 这可能是由于不同种类作物对硒的转运速率也不同[11].
此外, 小麦的试验结果表明, 根系在24 h处理下对SeNPs的吸收量高于亚硒酸盐, 然而72 h时, 亚硒酸盐的吸收量反而高于SeNPs.这可能是由于SeNPs被小麦根系吸收后会在植物体内转化为亚硒酸盐、硒酸盐等其他形态的硒[24].同时, 水稻的试验结果表明, 当pH为3.5和5.5时, 亚硒酸盐处理下水稻对硒的吸收量均显著高于SeNPs处理(P < 0.05), 这说明水稻作为淹水作物, 其吸收亚硒酸盐的能力要强于SeNPs.而当pH为7.5时, 水稻对两种形态硒的吸收无显著差异, 可能是因为pH对水稻吸收硒能力的影响远大于其对亚硒酸盐和SeNPs生物有效性的影响.
4 结论(1) 小麦和水稻均可以吸收SeNPs, 但其主要累积在植物根部.此外, SeNPs粒径越小越易从植物根部转运至地上部.
(2) SeNPs一般在植物的最适生长pH条件下易被吸收, 在pH较低时更易被转运; 亚硒酸盐在pH较低的环境下易被植物吸收, pH较高时易被转运.
| [1] | Wang J, Li H R, Yang L S, et al. Distribution and translocation of selenium from soil to highland barley in the Tibetan Plateau Kashin-Beck disease area[J]. Environmental Geochemistry and Health, 2017, 39(1): 221-229. DOI:10.1007/s10653-016-9823-3 |
| [2] | Ávila F W, Yang Y, Faquin V, et al. Impact of selenium supply on Se-methylselenocysteine and glucosinolate accumulation in selenium-biofortified Brassica sprouts[J]. Food Chemistry, 2014, 165: 578-586. DOI:10.1016/j.foodchem.2014.05.134 |
| [3] | Fairweather-Tait S J, Bao Y P, Broadley M R, et al. Selenium in human health and disease[J]. Antioxidants & Redox Signaling, 2011, 14(7): 1337-1383. |
| [4] | Rayman M P. The importance of selenium to human health[J]. The Lancet, 2000, 356(9225): 233-241. DOI:10.1016/S0140-6736(00)02490-9 |
| [5] |
姚远溪, 顾艳耿, 吕蒙, 等. 富硒蔬菜和谷物与人体健康分析[J]. 农产品加工, 2018(12): 80-82, 85. Yao Y X, Gu Y G, Lv M, et al. Analysed on the relationship between selenium-rich vegetables, selenium-rich grains and human health[J]. Farm Products Processing, 2018(12): 80-82, 85. |
| [6] | Broadley M R, White P J, Bryson R J, et al. Biofortification of UK food crops with selenium[J]. Proceedings of the Nutrition Society, 2006, 65(2): 169-181. DOI:10.1079/PNS2006490 |
| [7] |
张驰, 吴永尧, 彭振坤. 植物硒的研究进展[J]. 湖北民族学院学报(自然科学版), 2002, 20(3): 58-62. Zhang C, Wu Y Y, Peng Z K. Progress in the research of selenium in plants[J]. Journal of Hubei Institute for Nationalities (Natural Science Edition), 2002, 20(3): 58-62. DOI:10.3969/j.issn.1008-8423.2002.03.019 |
| [8] |
郭璐, 满楠, 梁东丽, 等. 小白菜对外源硒酸盐和亚硒酸盐动态吸收的差异及其机制研究[J]. 环境科学, 2013, 34(8): 3272-3279. Guo L, Man N, Liang D L, et al. Differences of selenium uptake pattern of Pakchoi and the possible mechanism when amended with selenate and selenite[J]. Environmental Science, 2013, 34(8): 3272-3279. |
| [9] | Fernández-Martínez A, Charlet L. Selenium environmental cycling and bioavailability:A structural chemist point of view[J]. Reviews in Environmental Science and Bio/Technology, 2009, 8(1): 81-110. DOI:10.1007/s11157-009-9145-3 |
| [10] | Slekovec M, Goessler W. Accumulation of selenium in natural plants and selenium supplemented vegetable and selenium speciation by HPLC-ICPMS[J]. Chemical Speciation & Bioavailability, 2005, 17(2): 63-73. |
| [11] |
陈思杨, 江荣风, 李花粉. 苗期小麦和水稻对硒酸盐/亚硒酸盐的吸收及转运机制[J]. 环境科学, 2011, 32(1): 284-289. Chen S Y, Jiang R F, Li H F. Uptake and translocation of selenate or selenite by wheat and rice seedlings[J]. Environmental Science, 2011, 32(1): 284-289. |
| [12] | Haug A, Graham R D, Christophersen O A, et al. How to use the world's scarce selenium resources efficiently to increase the selenium concentration in food[J]. Microbial Ecology in Health and Disease, 2007, 19(4): 209-228. DOI:10.1080/08910600701698986 |
| [13] | Ros G H, Van Rotterdam A M D, Bussink D W, et al. Selenium fertilization strategies for bio-fortification of food:An agro-ecosystem approach[J]. Plant and Soil, 2016, 404(1-2): 99-112. DOI:10.1007/s11104-016-2830-4 |
| [14] | Hariharan H, Al-Dhabi N A, Karuppiah P, et al. Microbial synthesis of selinium nanocomposite using Saccharomyces cerevisiae and its antimicrobial activity against pathogens causing nosocomial infection[J]. Chalcogenide Letters, 2012, 9(12): 509-515. |
| [15] | Forootanfar H, Adeli-Sardou M, Nikkhoo M, et al. Antioxidant and cytotoxic effect of biologically synthesized selenium nanoparticles in comparison to selenium dioxide[J]. Journal of Trace Elements in Medicine and Biology, 2014, 28(1): 75-79. DOI:10.1016/j.jtemb.2013.07.005 |
| [16] | Yang F, Tang Q M, Zhong X Y, et al. Surface decoration by Spirulina polysaccharide enhances the cellular uptake and anticancer efficacy of selenium nanoparticles[J]. International Journal of Nanomedicine, 2012, 7: 835-844. DOI:10.2217/nnm.11.154 |
| [17] | Benko I, Nagy G, Tanczos B, et al. Subacute toxicity of nano-selenium compared to other selenium species in mice[J]. Environmental Toxicology and Chemistry, 2012, 31(12): 2812-2820. DOI:10.1002/etc.1995 |
| [18] | Hu C H, Li Y L, Xiong L, et al. Comparative effects of nano elemental selenium and sodium selenite on selenium retention in broiler chickens[J]. Animal Feed Science and Technology, 2012, 177(3-4): 204-210. DOI:10.1016/j.anifeedsci.2012.08.010 |
| [19] | Shi L G, Xun W J, Yue W B, et al. Effect of sodium selenite, Se-yeast and nano-elemental selenium on growth performance, Se concentration and antioxidant status in growing male goats[J]. Small Ruminant Research, 2011, 96(1): 49-52. DOI:10.1016/j.smallrumres.2010.11.005 |
| [20] | Wang H L, Zhang J S, Yu H Q. Elemental selenium at nano size possesses lower toxicity without compromising the fundamental effect on selenoenzymes:Comparison with selenomethionine in mice[J]. Free Radical Biology and Medicine, 2007, 42(10): 1524-1533. DOI:10.1016/j.freeradbiomed.2007.02.013 |
| [21] | Gao X Y, Zhang J S, Zhang L D. Hollow sphere selenium nanoparticles:Their in-vitro anti hydroxyl radical effect[J]. Advanced Materials, 2002, 14(4): 290-293. DOI:10.1002/1521-4095(20020219)14:4<290::AID-ADMA290>3.0.CO;2-U |
| [22] | Li K E, Chang Z Y, Shen C X, et al. Toxicity of nanomaterials to plants[A]. In: Siddiqui M H, Al-Whaibi M H, Mohammad F (Eds.). Nanotechnology and Plant Sciences Nanoparticles and Their Impact on Plants[M]. Switzerland: Springer International Publishing, 2015. 101-123. |
| [23] | Lin Z H, Wang C R C. Evidence on the size-dependent absorption spectral evolution of selenium nanoparticles[J]. Materials Chemistry and Physics, 2005, 92(2-3): 591-594. DOI:10.1016/j.matchemphys.2005.02.023 |
| [24] | Hu T, Li H F, Li J X, et al. Absorption and bio-transformation of selenium nanoparticles by wheat seedlings (Triticum aestivum L.)[J]. Frontiers in Plant Science, 2018, 9: 1-10. DOI:10.3389/fpls.2018.00001 |
| [25] |
王晓芳, 陈思杨, 罗章, 等. 植物对硒的吸收转运和形态转化机制[J]. 农业资源与环境学报, 2014, 31(6): 539-544. Wang X F, Chen S Y, Luo Z, et al. Mechanisms of selenium uptake, translocation and chemical speciation transformation in plants[J]. Journal of Agricultural Resources and Environment, 2014, 31(6): 539-544. |
| [26] | Huang Q Q, Wang Q, Wan Y N, et al. Application of X-ray absorption near edge spectroscopy to the study of the effect of sulphur on selenium uptake and assimilation in wheat seedlings[J]. Biologia Plantarum, 2017, 61(4): 726-732. DOI:10.1007/s10535-016-0698-z |
| [27] | Li H F, Mcgrath S P, Zhao F J. Selenium uptake, translocation and speciation in wheat supplied with selenate or selenite[J]. New Phytologist, 2008, 178(1): 92-102. DOI:10.1111/j.1469-8137.2007.02343.x |
| [28] | Jiang Y, Zeng Z H, Bu Y, et al. Effects of selenium fertilizer on grain yield, Se uptake and distribution in common buckwheat (Fagopyrum esculentum Moench)[J]. Plant, Soil and Environment, 2016, 61(8): 371-377. DOI:10.17221/284/2015-PSE |
| [29] |
周越, 吴文良, 孟凡乔, 等. 土壤中硒含量、形态及有效性分析[J]. 农业资源与环境学报, 2014, 31(6): 527-532. Zhou Y, Wu W L, Meng F Q, et al. Review on the content, specification of selenium and its availability in soils[J]. Journal of Agricultural Resources and Environment, 2014, 31(6): 527-532. |
| [30] |
沈燕春, 周俊. 土壤硒的赋存状态与迁移转化[J]. 安徽地质, 2011, 21(3): 186-191. Shen Y C, Zhou J. Occurrence, migration and transformation of selenium in soil[J]. Geology of Anhui, 2011, 21(3): 186-191. DOI:10.3969/j.issn.1005-6157.2011.03.006 |
| [31] | Barker A V, Pilbeam D J. Handbook of plant nutrition (2nd ed.)[M]. Boca Raton, FL: CRC Press, 2015. |
| [32] |
王琪.水稻和小麦对有机硒的吸收、转运及形态转化机制[D].北京: 中国农业大学, 2017. Wang Q. Mechanisms of organic selenium uptake, translocation and speciation transformation in rice and wheat[D]. Beijing: China Agricultural University, 2017. http://cdmd.cnki.com.cn/Article/CDMD-10019-1017164649.htm |
| [33] |
张联合, 李友军, 苗艳芳, 等. pH对水稻离体根系吸收亚硒酸盐生理机制的影响[J]. 土壤学报, 2010, 47(3): 523-528. Zhang L H, Li Y J, Miao Y F, et al. Effects of pH on the physiological mechanism of selenite uptake by excised roots of rice seedlings[J]. Acta Pedologica Sinica, 2010, 47(3): 523-528. |