 2019, Vol. 40
2019, Vol. 40



工业化在促进我国经济高速发展的同时也引起一系列环境问题, 其中工业废水处理中现有硫化物脱除工艺尚存在缺陷[1].自养-异养联合反硝化(integrated autotrophic and heterotrophic denitrification, IAHD)工艺作为硫化物脱除的生物处理技术, 由于其可以利用硝酸盐作为电子受体同时进行硫化物, 硝酸盐和有机碳的降解近年来广泛应用于工业废水处理并受到越来越多的关注[2].相比于传统的自养反硝化脱硫和异养反硝化工艺, IAHD工艺具有诸多优点.从脱碳的角度看, 其减小了异养反硝化工艺对碳源的依赖性, 对于低碳氮比污水也可以取得较理想的处理效果;从脱硫脱氮的角度看, 其克服了自养反硝化工艺水力停留时间长, 构筑物容积较大及反硝化速率较低等缺点.根据笔者之前的研究, IAHD过程涉及两大类功能微生物:自养硫化物氧化-硝酸盐还原菌和异养硝酸盐还原菌(包括异养硫化物氧化-硝酸盐还原细菌).他们之间可能存在的互作途径如下[3, 4]:

|
(1) |
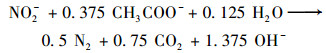
|
(2) |
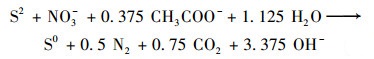
|
(3) |
围绕上述途径, 研究人员开发了诸多调控策略以维持3种功能微生物间的代谢平衡从而强化IAHD工艺的硫氮碳去除效率.例如调节C/N和S/N摩尔比[5]、优化反应器构型、控制水力停留时间(hydraulic retention time, HRT)和运行温度[6]等.然而, 微氧曝气是目前提高IAHD工艺运行效能的最有效策略. Chen等[7]首次将少量氧气引入IAHD工艺系统, 发现当反应器中的DO浓度为0.8 mg·L-1时, 硫化物和硝酸盐去除率均得到提升.微量氧气可以缓解体系内自养反硝化菌与异养反硝化菌对于唯一电子受体(NO3-)的竞争, 从而调节不同水质条件下IAHD的运行效能[8, 9].
研究者们试图从反应动力学、微生物群落结构、功能基因和功能酶的角度解析微氧条件对IAHD工艺运行效能的强化机制[10~12]. Chen等[13]对IAHD过程的物料平衡进行了详细衡算.然而, 该体系中的电子平衡尚未引起足够的关注.氧气作为电子受体被引入IAHD体系, 从电子转移平衡的角度进行探究可能是解析微氧对IAHD运行效能强化机制的关键突破.
因此, 本研究聚焦于IAHD过程中的电子平衡计算, 主要关注IAHD体系中的电子不平衡现象, 并发现了在微氧条件下电子缺失的现象相关性, 通过IAHD和硫化物氧化序批摇瓶实验进一步探究电子缺失率与硫氮碳去除效能特别是硫氧化过程的内在联系.
1 材料与方法 1.1 生物反应器的启动本实验采用UASB(up-flow anaerobic sludge blanket)生物反应器进行.反应器由有机玻璃制成, 工作容积为580 mL.接种污泥取自哈尔滨文昌污水处理厂二沉池污泥, 过筛沉淀后污泥的悬浮固体浓度(suspended solids, SS)和挥发性悬浮固体浓度(volatile suspended solids, VSS)分别为79.6 g·L-1和39.3 g·L-1, 接种量为280 mL.反应器在室温下运行, HRT为12 h.反应器进水中含有:硫化物(S2-)200 mg·L-1、硝酸盐(NO3-)465 mg·L-1、乙酸盐(Ac-)184.4 mg·L-1、氯化铵50 mg·L-1、三水合磷酸氢二钾50 mg·L-1、碳酸氢钠1 500 mg·L-1和微量元素[14]1 mL·L-1.
1.2 序批静态摇瓶实验待反应器运行稳定后, 从反应器底部取样口取泥样, 用去离子水离心洗涤3次后进行IAHD序批静态摇瓶实验, 经预处理后, 待接种污泥的SS及VSS含量分别为38.8 g·L-1和22.1 g·L-1.厌氧血清瓶体积为250 mL, 污泥接种体积为5 mL, 培养基(成分与反应器进水相同)体积为95 mL.接种前用氩气对培养基及瓶体上方空气吹脱15 min, 以除掉氧气形成完全厌氧的条件.除厌氧实验组外, 为探究氧气对IAHD过程的影响, 分别向两个实验组的血清瓶中注入5 mL和10mL的氧气, 在注入氧气前, 抽出等量的瓶体上方气体以维持注入氧气后的瓶中的气压.每个实验组设置3个平行, 3个实验组均置于摇床中培养, 摇床转速为135 r·min-1, 反应温度为30℃. 3组反应结束后, 收集接种污泥, 每组分别经去离子水离心洗涤3次, 用于进行后续的硫化物氧化序批静态摇瓶实验.每瓶的污泥接种量及培养基体积均与IAHD序批静态摇瓶实验保持一致.硫化物氧化培养基除不含有乙酸盐和硝酸盐外, 其余组分及浓度均与IAHD培养基相同.接种污泥前对培养基及上方空气曝氩气除氧, 同样设置微氧5 mL和10 mL实验组及化学对照组(厌氧瓶中注射10 mL无污泥接种), 瓶内压强通过抽除与氧气等体积的气体保持恒定, 每组设置3个平行.所有厌氧血清瓶均置于摇床中30℃培养, 转速135 r·min-1.
1.3 数据测定本实验中污泥样品的SS和VSS采用103~105℃烘干恒重法测定[15].所有水样在取样后先在12 000 r·min-1下离心3 min, 后通过孔径0.22 μm的聚醚砜水系微孔滤膜过滤掉水样中的微生物杂质待测.硫离子的测定采用亚甲基蓝分光光度法[16].水样中乙酸根、亚硝酸根、硝酸根、硫酸根和硫代硫酸根离子浓度采用离子色谱(Dionex ICS-3000)测定.采用的离子色谱柱型号为AS11-HC 4×250 mm, 淋洗液为浓度1.5 mmol·L-1的NaOH, 流速为1 cm3·min-1.水中溶解氧浓度由型号pH/Oxi 340i的溶解氧仪测定. pH值由型号为pHS-25的pH仪测定.气体中的O2含量由安捷伦7890A型气相色谱测定.
1.4 DNA提取及高通量测序待硫化物氧化序批实验结束后, 分别收集各实验组污泥, 用土壤基因组DNA提取试剂盒(DP336)进行DNA的提取.选取515F (5′-GTGC CAGCMGCCGCGG-3′)和907R (5′-CCGTCAATTC MTTTRAGTTT-3′)作为引物对细菌16S rRNA在V4-V5区进行扩增. 20 μL的扩增体系包括:DNA母版10 ng, 5×FastPfu缓冲液4 μL, dNTP s (2.5 mmol·L-1) 2 μL, 正向引物(5 μmol·L-1) 0.8 μL, 反向引物(5 μmol·L-1) 0.8 μL, FastPfu DNA聚合酶0.4 μL, 及无菌双蒸水12 μL. PCR扩增程序为:95℃初始变性3 min, 30个循环, 其中95℃变性30 s, 55℃退火30 s, 72℃延伸45 s, 循环结束后最终于72℃下延伸10 min. PCR产物经1%琼脂糖凝胶电泳-紫外投射核实扩增成功, 采用AxyPrep DNA纯化试剂盒对扩增产物进行纯化并进行后续高通量测序.采用Illumina MiSeq PE250平台对群落DNA片段进行双端测序, 调用Usearch (vsesion 7.0, http://drive5.com/uparse/)软件对所有相似度大于97%的序列进行OTU分类划分.每个OTU对应的物种分类信息, 采用RDP classifier贝叶斯算法依照Silva(Release128, http://www.arb-silva.de)数据库进行分类学分析. 16S rRNA原始测序序列上传至NCBI数据库中, 登录号为:SRP125349.
1.5 电子平衡计算在连续流IAHD系统中, 硫化物和乙酸盐被氧化, 失去电子充当电子供体, 而伴随进水裹挟入反应器的微量氧气和硝酸盐被还原, 得到电子充当电子受体.由于硫化物本身呈现强还原性, 在进水中硫化物浓度为200 mg·L-1时无法检测到DO值.在不考虑氧气的情况下, 依据反应器内的物质转化及物料守恒, 参见以下公式对IAHD反应器进行初步的电子平衡计算:
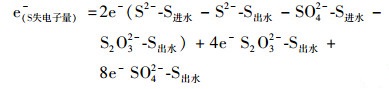
|
(4) |
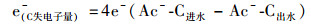
|
(5) |
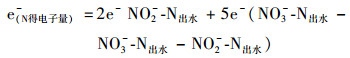
|
(6) |
式中, 各物质分别代表IAHD反应器进出水的硫、氮和碳系物的浓度(mmol·L-1).
在序批静态摇瓶实验中, 依据反应初始硫化物, 硝酸盐和乙酸盐浓度及反应结束时硫氮碳氧化及还原产物的浓度对整个过程进行电子平衡分析, 参见上式进行计算.注入氧气的摩尔质量在已知氧气体积的情况下依据理想气体状态方程(PV=nRT)进行计算.
2 结果与分析 2.1 连续运行的UASB-IAHD反应器的效能在连续运行的41 d内, 进水硫化物浓度始终保持在200 mg·L-1左右, S:N:C的摩尔比为5:6:5. 图 1和表 1展示了UASB生物反应器的详细运行参数及运行效能.如图 1所示, 在整个运行周期内, 硫化物, 硝酸盐和乙酸盐的去除率均为100%.反应器出水中没有检测到硫代硫酸盐, 70.0%的进水硫化物在微生物的作用下转化成硫酸盐从反应器流出.出水亚硝酸盐积累率在开始启动的第1~15 d波动很大, 达到平稳期后稳定在6.3%.

|
S :N :C=5 :6 :5, S2-=200 mg·L-1 图 1 生物反应器连续运行41 d的进出水指标 Fig. 1 Bioreactor performance over the 41 operational days |
|
|
表 1 UASB生物反应器稳态运行性能和电子转化 Table 1 Operational performance and electron transformation in IAHD bioreactor in the steady state |
如表 1所示, 硝酸盐在去除和转化的过程中接受电子的量为39.37 mmol·L-1, 不能满足硫化物(-38.31 mmol·L-1)和乙酸盐(-27.02 mmol·L-1)生成氧化产物的电子需求.笔者将电子得失不平衡的差值与接受电子的比率定义为电子缺失率.在连续流反应器的稳定运行阶段, 电子得失差值为20.94 mmol·L-1, 此时的电子缺失率为65.9%, 这可能与体系内无法检测的微量氧气有关. Chen等[7]也发现了微氧条件下IAHD过程中的电子不平衡现象, 其利用EGSB生物反应器在微氧条件下对硫化物负荷为800 mg·L-1的IAHD过程进行强化, 硫化物、硝酸盐和乙酸盐的去除率均为100%时, 电子缺失率为31.5%.为了进一步明确微量氧气在IAHD工艺中对电子得失不平衡的贡献度, 将氧气含量作为IAHD过程中的独立变量来研究碳氮硫共脱除过程中的电子转移.
2.2 不同氧气含量下的IAHD序批静态摇瓶实验从稳定运行的IAHD生物反应器中取出共50 mL的驯化污泥进行序批实验. 图 2显示了IAHD过程中不同氧气含量下硫化物, 硝酸盐和乙酸盐的去除与转化规律.在氧气含量为10 mL的条件下, 硫化氢和乙酸盐在1.5 h内降解至0 mmol·L-1[图 2(a)].当氧气含量为5 mL和0 mL时, 此时间缩短至0.5 h.摇瓶实验中, 硫酸盐是硫化物的主要氧化产物.当氧含量为10、5和0 mL时, 硫酸盐转化率分别为84%、78%和48%.可以看到, 随着氧气含量的增高, 硫酸盐转化率逐渐增高.无论在何种氧气含量条件下, 当硝酸盐浓度几乎降低至零时, 亚硝酸盐转化率高达50%, 出现亚硝酸盐的累积峰.产生的亚硝酸盐继续发生反硝化反应, 当亚硝酸盐浓度降低至0 mmol·L-1, 在5 mL和0 mL的氧气注入量下再次检测到硫化物, 且其浓度出现累积上升的趋势.在5 mL的氧气含量下, 硫化物浓度在反应开始3 h后发生反弹, 在5 h时浓度增加至0.81 mmol·L-1.在厌氧条件下(0 mL氧气), 硫化物在反应开始1.5 h后发生反弹, 且在反应结束时反弹至初始硫化物浓度的一半(2.92 mmol·L-1).氧含量的增加明显抑制了硫化物的这种反弹现象. Zhou等[17]使用古菌引物对IAHD体系中的污泥进行高通量测序, 发现Methanobacterium丰度较高. Methanobacterium可以利用乙酸盐作为电子供体将单质硫还原为硫化物, 因此推测IAHD体系中的硫化物反弹可能和这类功能微生物关系密切.

|
图 2 不同氧气含量下的硫氮碳降解规律 Fig. 2 Time course of sulfide, nitrate, and acetate removal and transformation in IAHD batch tests |
计算在IAHD序批静态摇瓶实验过程中的电子得失情况(表 2), 可以发现在不同氧气含量的条件下, 均存在电子得失不平衡的现象, 即电子受体缺失. 10、5和0 mL氧气条件下, 电子缺失率分别为24.3%、55.1%和39.2%.如图 2所示, 在氧气含量为5 mL的情况下, 最初0.5 h内硫化物, 硝酸盐和乙酸盐的去除率[mmol·(L·h)-1]高于氧气含量为10 mL时的去除率, 而硫化物反弹后的最终浓度(0.81 mmol·L-1)则低于厌氧条件下硫化物反弹后的最终浓度(2.92 mmol·L-1).因此, 综合以上指标考量IAHD过程的运行效能, 推测电子缺失率越大, IAHD可能更能获得更稳定更高效的硫氮碳去除效能.
|
|
表 2 硫化物浓度为200 mg·L-1时不同氧气含量下IAHD过程的电子平衡计算 Table 2 Electron balance calculation of batch tests with sulfide concentration of 200 mg·L-1 under different oxygen contents |
2.3 不同氧气含量下的硫化物氧化序批静态摇瓶实验
在IAHD序批实验结束后, 分别将不同氧气含量下的每组污泥取出并用蒸馏水洗涤3次, 注入氧气作为唯一的电子受体进行硫化物氧化序批实验(图 3).如图 3(a)和3(b)所示, 当氧气含量为10 mL和5 mL时, 硫化物到硫代硫酸盐的转化率分别为66.9%和32.4%.除了在10 mL氧含量下生成了2.07 mmol·L-1的SO42--S外, 厌氧和氧气含量为5 mL的条件下几乎没有硫酸盐生成[图 3(a)].在化学对照组中, 硫代硫酸盐也是硫化物主要的氧化产物, 转化率为39%[图 3(d)].当没有氧气注入时(厌氧条件)[图 3(c)], 整个体系没有电子受体, 硫化物不发生氧化, 浓度始终保持在6 mmol·L-1左右.

|
图 3 不同氧气含量下的硫化物及其氧化产物的变化规律 Fig. 3 Time evolution of sulfide, sulfate and thiosulfate concentrations for sulfide oxidizing batch tests |
表 3中计算了硫化物氧化序批实验中, 硫化物利用氧气发生氧化过程中的电子得失情况.在厌氧条件下, 硫化物不发生氧化, 体系中没有电子转移的发生.当氧气含量为5 mL和10 mL时, 电子受体缺失的量为0.14 mmol和0.73 mmol, 电子缺失率为18.7%和38.2%.在化学对照组中, 硫化物氧化成硫酸盐和硫代硫酸盐失去的1.2 mmol电子完全被氧气接受, 不存在电子受体缺失的现象.根据以上4组实验的结果可知, 在硫氧化过程当中出现的电子得失差值仅存在于生物硫化物氧化过程中, 且电子缺失率与氧含量呈现正相关.
|
|
表 3 以氧气作为唯一电子受体时硫系物的浓度变化及电子平衡计算 Table 3 Variation of sulfur containing compounds and electronic balance calculation in batch tests with oxygen only |
2.4 微生物群落结构分析
在硫化物氧化序批实验结束后, 采用Illumina二代测序对各个实验组的活性污泥进行微生物群落结构分析.在97%序列相似性水平上鉴定了424个OTU. 表 4中给出了群落的Shannon、Simpson、Ace和Chao1指数. Ace和Chao1指数反映微生物群落的丰富度, 该值越大, 则群落的丰富度越高. Shannon和Simpson指数则在考虑群落丰度的基础上也可以反映群落的均匀度, Shannon指数越大, Simpson指数越小则表示群落的多样性越高[18].厌氧条件下, 微生物群落的多样性最高, 随着氧气含量的增加, 微生物群落多样性逐渐降低.
|
|
表 4 微生物群落的多样性及丰富度指数 Table 4 Species richness and diversity analysis of microbial communities |
图 4展示了微生物群落结构在属水平上的分类, 测序丰度小于0.5%的属统一归类为others.在硫氧化序批摇瓶实验的群落结构中, Thiobacillus、Thauera、Mangroviflexus和Erysipelothrix是主要占优属, 在S0、S5和S10中总相对丰度达75%以上. Thiobacillus是污水处理过程中常见的化能自养微生物, 可以利用硝酸盐, 亚硝酸盐和氧气作为电子受体进行硫化物, 单质硫和硫代硫酸盐等硫系物的氧化[19].随着氧气含量的增加, 微生物群落中Thiobacillus的相对丰度逐渐增加, 在S0、S5和S10中分别占54%、60%和61%.作为反硝化脱氮工艺及IAHD过程中常见的异养微生物, Thauera在不同氧气含量下其丰度占到总群落11%~15%. Erysipelothrix也可以利用有机碳源进行生长繁殖, 同时可以将硝酸盐还原成氮气[20].在IAHD批次实验中, 最初0.5 h的硝酸盐还原速率为5.88 mmol·(L·h)-1, 这仅为氧气含量为5 mL和0 mL(厌氧)时的一半.相应地, 在S5和S0中, Erysipelothrix的相对丰度分别为2%和5%, 而在S10中仅仅占到1%.据报道, Mangroviflexus是一类兼性厌氧菌, 同时具有将硫酸盐或亚硫酸盐还原为硫化物的能力[21].因此, 推测Mangroviflexus可能与IAHD序批测试中的硫化物反弹现象相关.在厌氧条件及氧气含量为5 mL时, Mangroviflexus的丰度分别占3%和4%, 而在氧气含量为10 mL硫化物反弹现象消失的情况下, 其比例下降至0%.

|
图 4 硫化物氧化序批实验中不同氧气含量下的微生物群落结构 Fig. 4 Bacterial community structures at genus level in sulfide oxidizing batch tests corresponding to different oxygen contents |
从丰度排名前15的属可以看出, 经IAHD过程长期驯化的接种污泥, 在仅有硫化物与氧气共存的硫氧化环境下, 自养反硝化菌虽绝对占优, 但群落结构仍具有自养反硝化与异养反硝化菌共存的特征, 这与先前的研究工作相符[22].本课题组前期利用经IAHD过程成功驯化的活性污泥, 在硫化物浓度为200 mg·L-1, 硫:氮:碳摩尔比为5:6:5的条件下设置不同氧气梯度探究最适宜的微氧浓度, 随着氧气含量的升高, 也发现了IAHD过程中的电子缺失率逐渐升高的现象[9].微生物群落分析结果表明在氧气含量为8 mL和12 mL的情况下, Thauera、Azoarcus和Thiobacillus为占优属, 其中Thauera的丰度高达60%左右.但是, 由于微氧的IAHD过程涉及多种电子供体及受体, 硫化物好氧氧化及厌氧氧化, 自养及异养反硝化以及有机物的好氧氧化过程同时发生, 微氧IAHD群落结构在属水平的分布特征与电子缺失率并没有展现出明显相关性.因此, 针对某一过程建立占优属与电子缺失率间的联系对于解读微氧IAHD中的电子不平衡现象具有重要意义.
作为硫化物氧化群落结构中丰度最高的属, Thiobacillus的相对丰度随着氧气含量的增加而增加(S0: 54%、S5: 60%和S10: 61%), 并且与硫化物氧化序批实验中的电子得失差值呈现一定的正相关关系.目前, 已有文献报道了Thiobacillus可以在电子受体不足的条件下进行硫化物氧化过程. Janssen等[23]证明, 接种在连续流气升式生物反应器中的混合Thiobacillus菌株, 能够在O2/H2S摩尔比低至0.35时将约20%硫化物完全氧化成硫酸盐, 经计算, 其电子缺失率为128%.此外, Alcántara等[24]在其生物反应器中也以Thiobacillus属的纯菌作为接种物, 当O2/H2S摩尔比为1.0时发现60%的硫化物完全氧化成了硫酸盐, 而基于电子平衡计算, 反应器中提供的氧气仅仅能达到33%的硫酸盐转化率. Klok等[25]依据其运行的曝气生物反应器中的现象提出了一条硫化物在碱性条件下发生氧化时可能存在的特殊电子传递途径(limited oxygen route, LOR).其中硫化物氧化的直接电子受体不是氧气, 而是通过蒽醌类化合物将电子传递给NAD+, 再通过胞内NAD+与NADH间的循环再生实现硫化物的氧化.但在IAHD过程中, 除发生硫化物氧化过程外, 还存在硝酸盐和亚硝酸盐的还原及有机物的降解过程, 因此, 是否仅在硫化物的氧化过程中存在电子得失的差值, 以及通过碳固定再生的NAD+是否是IAHD过程中潜在的内源电子受体, 仍需要进一步实验验证.
与常规好氧工艺相比, 采用微氧工艺可以显著节省曝气运行的成本.针对IAHD过程来说, 微氧条件是一种可以同步强化硫化物、硝酸盐和有机物去除效率的有效手段.在本研究中, 对反应器中硫、氮、碳进行电子平衡计算发现了微氧条件下运行的IAHD反应器中存在电子得失不平衡的现象, 并且试图建立电子缺失率, 功能微生物和高效IAHD性能之间的关系.通过IAHD序批实验发现当电子缺失率最大时, 可以获得满意的IAHD性能.因此, 通过对IAHD过程的电子平衡计算可以对曝气速率及曝气量进行优化, 以在微氧条件下获得最高的硫氮碳去除效率.此外, 找出潜在的内源电子受体不仅可以明确自养反硝化和异养反硝化菌对于体系的贡献度, 进一步解析IAHD过程中硫, 氮和碳的生物降解机制, 而且可以为微氧强化IAHD工艺在实际应用中提供调控的新思路.
3 结论(1) 在微氧条件下, 当硫化物浓度为200 mg·L-1, S:N:C为5:6:5时, IAHD生物反应器在硝酸盐供给不足的情况下, 可以实现100%的硫化物, 硝酸盐和乙酸盐去除率以及70%的硫酸盐转化率. IAHD序批实验中达到最佳的硫、氮、碳去除效能时, 相应的电子缺失率也达到最大值, 为55.1%.
(2) 电子得失不平衡现象仅存在于硫化物的生物氧化过程中, 且氧气含量越高电子缺失率越大, 氧气含量和电子缺失率之间存在正相关关系.
(3) 经过IAHD过程驯化的活性污泥, 在硫氧化条件下进行微生物群落结构分析, Thiobacillus为主要占优属, 在厌氧、氧气含量5 mL及10 mL的条件下丰度分别为54%、60%和61%, 其相对丰度随着电子缺失率的增大而增加.
(4) 微氧条件下的电子得失不平衡现象打破了先前需要严格依照化学计量比例来运行IAHD工艺的思维模式, 给出了微氧强化可以拓宽IAHD工艺水质适用范围的另一理论依据.
| [1] |
李名升, 周磊, 陈远航, 等. 经济结构调整的污染减排效应:以COD减排为例[J]. 环境科学, 2014, 35(8): 3212-3218. Li M L, Zhou L, Chen Y H, et al. Effect of economic structure adjustment on pollution emission:a case study of COD[J]. Environmental Science, 2014, 35(8): 3212-3218. |
| [2] |
李祥, 马航, 黄勇, 等. 异养与硫自养反硝化协同处理高硝氮废水特性研究[J]. 环境科学, 2016, 37(7): 2646-2651. Li X, Ma H, Huang Y, et al. Characteristics of a combined heterotrophic and sulfur autotrophic denitrification technology for removal of high nitrate in water[J]. Environmental Science, 2016, 37(7): 2646-2651. |
| [3] |
谭文勃, 马晓丹, 黄聪, 等. 1株异养反硝化硫细菌的分离鉴定及代谢特性[J]. 环境科学, 2017, 38(2): 809-814. Tan W B, Ma X D, Huang C, et al. Isolation, identification and metabolic characteristics of a heterotrophic denitrifying sulfur bacterial strain[J]. Environmental Science, 2017, 38(2): 809-814. |
| [4] | Zhang R C, Xu X J, Chen C, et al. Interactions of functional bacteria and their contributions to the performance in integrated autotrophic and heterotrophic denitrification[J]. Water Research, 2018, 143: 355-366. DOI:10.1016/j.watres.2018.06.053 |
| [5] |
郭磊, 王爱杰, 刘春爽, 等. 碳氮比对碳氮硫同步脱除的影响[J]. 给水排水, 2008, 34(S1): 224-227. Guo L, Wang A J, Liu C S, et al. Effect of C/N ratio on the simultaneous removal of carbon, nitrogen and sulfur[J]. Water & Wastewater Engineering, 2008, 34(S1): 224-227. |
| [6] | Ghorbel L, Coudert L, Gilbert Y, et al. Determination of critical operational conditions favoring sulfide production from domestic wastewater treated by a sulfur-utilizing denitrification process[J]. Journal of Environmental Management, 2017, 198: 16-23. |
| [7] | Chen C, Ren N Q, Wang A J, et al. Enhanced performance of denitrifying sulfide removal process under micro-aerobic condition[J]. Journal of Hazardous Materials, 2010, 179(1-3): 1147-1151. DOI:10.1016/j.jhazmat.2010.02.065 |
| [8] | Chen C, Zhang R C, Xu X J, et al. Enhanced performance of denitrifying sulfide removal process at high carbon to nitrogen ratios under micro-aerobic condition[J]. Bioresource Technology, 2017, 232: 417-422. DOI:10.1016/j.biortech.2017.02.031 |
| [9] | Chen C, Shao B, Zhang R C, et al. Mitigating adverse impacts of varying sulfide/nitrate ratios on denitrifying sulfide removal process performance[J]. Bioresource Technology, 2018, 267: 782-788. DOI:10.1016/j.biortech.2018.07.096 |
| [10] | Wang X W, Zhang Y, Zhang T T, et al. Effect of dissolved oxygen on elemental sulfur generation in sulfide and nitrate removal process:characterization, pathway, and microbial community analysis[J]. Applied Microbiology and Biotechnology, 2016, 100(6): 2895-2905. DOI:10.1007/s00253-015-7146-4 |
| [11] |
于皓, 陈川, 张莉, 等. 溶解氧对碳氮硫共脱除工艺中微生物群落影响解析[J]. 环境科学, 2013, 34(6): 2368-2374. Yu H, Chen C, Zhang L, et al. Effect of dissolved oxygen on microbial community in simultaneous removal of carbon, nitrogen and sulfur process[J]. Environmental Science, 2013, 34(6): 2368-2374. |
| [12] | Guo H L, Chen C, Lee D J, et al. Proteomic analysis of sulfur-nitrogen-carbon removal by Pseudomonas sp. C27 under micro-aeration condition[J]. Enzyme and Microbial Technology, 2014, 56: 20-27. DOI:10.1016/j.enzmictec.2013.12.013 |
| [13] | Chen C, Wang A J, Ren N Q, et al. High-rate denitrifying sulfide removal process in expanded granular sludge bed reactor[J]. Bioresource Technology, 2009, 100(7): 2316-2319. DOI:10.1016/j.biortech.2008.10.023 |
| [14] | Zhang R C, Xu X J, Chen C, et al. Bioreactor performance and microbial community analysis of autotrophic denitrification under micro-aerobic condition[J]. Science of the Total Environment, 2019, 647: 914-922. DOI:10.1016/j.scitotenv.2018.07.389 |
| [15] |
周虹丽, 朱建华, 韩冰, 等. 重量法测量悬浮物浓度关键技术研究[J]. 海洋技术学报, 2004, 23(3): 15-20. Zhou H L, Zhu J H, Han B, et al. Research on the key technique of the analysis of suspended matter by weighting method[J]. Ocean Technology, 2004, 23(3): 15-20. |
| [16] |
林李秋. 亚甲基蓝分光光度法测定水中硫化物影响因素的探讨[J]. 化学工程师, 2006, 20(10): 32-33, 35. Lin L Q. Study on factors effecting the determination of sulfide in water by methylene blue spectrophotometry[J]. Chemical Engineer, 2006, 20(10): 32-33, 35. DOI:10.3969/j.issn.1002-1124.2006.10.014 |
| [17] | Zhou X, Liu L H, Chen C, et al. Reduction of produced elementary sulfur in denitrifying sulfide removal process[J]. Applied Microbiology and Biotechnology, 2011, 90(3): 1129-1136. DOI:10.1007/s00253-011-3087-8 |
| [18] | Haegeman B, Sen B, Godon J J, et al. Only Simpson diversity can be estimated accurately from microbial community fingerprints[J]. Microbial Ecology, 2014, 68(2): 169-172. |
| [19] |
车轩, 罗国芝, 谭洪新, 等. 脱氮硫杆菌的分离鉴定和反硝化特性研究[J]. 环境科学, 2008, 29(10): 2931-2937. Che X, Luo G Z, Tan H X, et al. Isolation, identification and denitrification characterization of Thiobacillus denitrificans[J]. Environmental Science, 2008, 29(10): 2931-2937. DOI:10.3321/j.issn:0250-3301.2008.10.042 |
| [20] | Mielcarek A, Rodziewicz J, Janczukowicz W, et al. Citric acid application for denitrification process support in biofilm reactor[J]. Chemosphere, 2017, 171: 512-519. DOI:10.1016/j.chemosphere.2016.12.099 |
| [21] | Han R, Yuan Y Z, Cao Q W, et al. PCR-DGGE analysis on microbial community structure of rural household biogas digesters in Qinghai Plateau[J]. Current Microbiology, 2018, 75(5): 541-549. DOI:10.1007/s00284-017-1414-8 |
| [22] | Huang C, Liu Q, Chen C, et al. Elemental sulfur recovery and spatial distribution of functional bacteria and expressed genes under different carbon/nitrate/sulfide loadings in up-flow anaerobic sludge blanket reactors[J]. Journal of Hazardous Materials, 2017, 324: 48-53. DOI:10.1016/j.jhazmat.2016.03.024 |
| [23] | Janssen A J H, Meijer S, Bontsema J, et al. Application of the redox potential for controling a sulfide oxidizing bioreactor[J]. Biotechnology and Bioengineering, 1998, 60(2): 147-155. DOI:10.1002/(SICI)1097-0290(19981020)60:2<147::AID-BIT2>3.0.CO;2-N |
| [24] | Alcántara S, Velasco A, Muñoz A, et al. Hydrogen sulfide oxidation by a microbial consortium in a recirculation reactor system:sulfur formation under oxygen limitation and removal of phenols[J]. Environmental Science & Technology, 2004, 38(3): 918-923. |
| [25] | Klok J B M J, Van Den Bosch P L F, Buisman C J N, et al. Pathways of sulfide oxidation by haloalkaliphilic bacteria in limited-oxygen gas lift bioreactors[J]. Environmental Science & Technology, 2012, 46(14): 7581-7586. |