 2019, Vol. 40
2019, Vol. 40



2. 宁波大学海洋学院, 宁波 315211
2. School of Marine Sciences, Ningbo University, Ningbo 315211, China
我国是猪业生产的第一大国, 近年来, 随着国家经济的快速发展, 养猪业在技术水平、规模化程度等方面也得到了一定的进步和提高, 但经济效益增加的同时也衍生了一系列环境问题, 如养猪业沼气工程在提供清洁能源的同时, 还产生了大量的沼渣和沼液废水, 富营养化的沼液废水对水环境和人类健康产生了严重的危害[1, 2].猪场废水具有有机物、悬浮物浓度高, 富含高氨氮, 处理难度大等特点[3].目前我国规模化养猪场的废水一般采用厌氧-好氧组合工艺进行处理, 然而该方法处理后的沼液中难降解的有机物、氮磷浓度等仍然很高.且当碳氮比较低时, 采用一些接触氧化法、SBR法和氧化沟法等方法处理后, 效果不太理想, 运行不稳定, 建设成本较高, 难以在工程中实际运用[2].国内外对养猪沼液废水的处理已有大量研究, 研究人员也相继开发了不同的养猪沼液废水处理工艺, 但传统的处理方式虽能在一定程度上达到除污的目的, 同时也带来了二次污染.因此, 微藻对养猪沼液废水高效、经济地处理工艺对环境改善与资源化利用都有一定意义[4].
微藻是一种结构简单的光合水生植物, 它能够高效地进行光合作用, 将太阳能转化为可利用的化学能.微藻具有分布广泛, 生长速度快, 产率高, 蛋白质、油脂和多糖等附加值含量高等优点[5].大部分微藻属于光合自养型, 微藻细胞能够利用水体中N、P及微量元素进行光能自养生长[6].微藻的生长对废水的水质情况要求较低, 可以利用光能富集水体中的氮、磷等营养物质, 同时利用二氧化碳为碳源合成有机物, 将无机盐以有机物的形式储存在藻细胞中[7].
沼液是畜禽粪污经厌氧发酵后的产物, 含有氨氮浓度范围约在17~2 050 mg ·L-1.国家废水排放氨氮标准浓度为80 mg ·L-1, 珠三角沼液废水排放氨氮标准浓度为70 mg ·L-1, 应用新技术处理养猪污水时, 自污水净化系统开始运行后, 对系统中各个单元的净化功能进行较长时间的现场观测, 当到达最后一个净化系统氧化塘时氨氮平均值为53.7 mg ·L-1[8].
微藻种类多样、适应性强, 能够在多种生态环境中生长.养猪沼液中富含碳源和氮源等, 可为微藻的生长提供养料.微藻净化沼液能够同时降低沼液中的营养成分和有机物.将猪粪沼液废水净化与微藻的培养相结合, 有望达到排放标准, 还可降低猪粪沼液废水处理成本.微藻在完成自身细胞的增殖与营养物质积累的同时, 实现了沼液废水的无害化处理和微藻生物质高附加值的资源化利用[8~10].但实际利用沼液废水培养微藻时, 过高浓度的NH4+-N会超出微藻的耐受性, 抑制藻细胞的生长, 高氨氮问题是利用微藻处理沼液废水的难点[11].沼液废水中高浓度的氨氮对藻细胞的毒害作用主要是抑制藻细胞的光合系统, 而这一过程可通过藻细胞内的抗氧化体系来反映.但是不同藻株对不同高氨氮浓度的适应性不同, 而藻株的特性与应用价值也是微藻处理沼液资源化利用考虑的方面[12, 13].
基于上述背景, 本文以前期筛选获得的衣藻、葡萄藻、紫球藻和栅藻为研究对象, 通过液体培养, 模拟沼液废水浓度于培养基中设置不同氨氮浓度, 探讨各藻株在不同氨氮浓度下的生长状况, 寻找近似沼液废水氨氮浓度下的优势藻种, 并讨论在高氨氮沼液废水中各藻种SOD、POD、CAT、GR酶活性及MDA含量的变化, 以期为微藻处理沼液废水提供理论基础.
1 材料与方法 1.1 藻株来源与培养本实验所用藻种购于湖北武汉中国科学院水生生物研究所, 藻种培养过程所用培养基为BG11、SE+TAP和KOCK, 其中衣藻的培养基为SE+TAP; 紫球藻的培养基为KOCK; 其他藻种为BG11[14~16].
实验液体培养所用反应器为玻璃柱式反应器, 内直径0.05 m, 柱高0.55 m, 反应体积0.9 L.反应器内部布置直径5 mm的玻璃通气管, 通入混合有1.5% CO2(体积分数)的压缩空气(0.1 MPa), 以1 mL ·s-1的速率通过通气管从反应器底部曝气, 使藻液搅动并补充碳源.经过前期研究证实, 在光照强度80~100 μmol ·(m2 ·s)-1下, 微藻的藻细胞生长较好.为了考察藻细胞生长的最高效率, 微藻培养过程中连续光照, 培养柱表面光强100 μmol ·(m2 ·s)-1, 培养温度(25±1)℃.
1.2 沼液废水本文藻种培养所用沼液废水取自江西省九江市境内某养殖场经厌氧发酵后的典型沼液废水, 取水时间为2018年4月12日10:00.废水经自然沉降2 d后, 取上清液测定其初始氨氮(NH4+-N)、Cu2+、总磷(TP)和COD等各指标(表 1).
|
|
表 1 沼液废水的参数指标 Table 1 Features of the raw piggery wastewater |
1.3 氨氮浓度的设置
本实验使用NH4Cl作为N源, 沼液是畜禽粪污经厌氧发酵后的产物, 含有氨氮浓度范围为17~2 050 mg ·L-1.国家废水排放氨氮标准浓度为80 mg ·L-1, 珠三角沼液废水排放氨氮标准浓度为70 mg ·L-1, 应用新技术处理养猪污水时, 自污水净化系统开始运行后, 对系统中各个单元的净化功能进行较长时间的现场观测, 当到达最后一个净化系统氧化塘时氨氮平均值为53.7 mg·L-1[17].
以此为基准, 分别添加不同的NH4Cl浓度, 使得本实验所用培养基中氨氮终浓度分别为:低浓度50 mg ·L-1、近似沼液废水浓度500 mg ·L-1和高浓度2 000 mg ·L-1.
1.4 藻细胞的生物量测定与生物产率计算将0.45 μm, 直径50 mm, 面积为0.001 m2的醋酸纤维滤膜煮沸3次后, 在105℃烘箱中烘至恒重(W1), 将待测藻细胞(DW, mg)用移液器取15 mL藻样到抽滤装置内, 抽滤至已称重的滤膜上, 将附着藻的滤膜放入105℃烘箱中烘至恒重(W2), 用分析天平称量, 并计算藻细胞的生物量(g ·L-1).
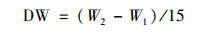
|
基于本研究培养时间为10 d, 待测藻生物产率[BP, g·(L ·d)-1]计算公式如下:
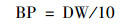
|
TP测定采用钼酸铵分光光度法(GB 11893-1989); NH4+-N的测定采用纳氏试剂分光光度法(GB 7479-1987); COD的测定采用重铬酸钾氧化处理法(GB 11914-1989).
1.6 酶活性的测定方法本次实验所用测定酶活性的方法, 试剂盒均购置于南京建成生物工程研究所.
采用黄嘌呤氧化酶法(羥胺法)测定SOD:超氧化物歧化酶活力通过黄嘌呤及黄嘌呤氧化酶反应系统产生超氧阴离子(O2-), O2-可还原氮蓝四唑生成蓝色甲臜, 后者在560 nm处有吸收; SOD可清除O2-, 从而抑制了甲臜的形成;
采用愈创木酚法测定过氧化物酶(POD)活性:该方法可催化过氧化氢氧化酚类和胺类化合物, 具有消除化氢和酚类、胺类毒性的双重作用. POD催化H2O2氧化特定底物, 在470 nm有特征光吸收.
过氧化氢酶(CAT)的原理在于H2O2在240 nm下有特征吸收峰, CAT能够分解H2O2, 使反应溶液240 nm下的吸光度随反应时间而下降, 根据吸光度的变化率可计算出CAT活性.
谷胱甘肽还原酶(GR)的原理在于GR能催化NADPH还原GSSG再生GSH, 同时不断消耗NADPH生成NADP+; NADPH在340 nm有特征吸收峰; 通过测定340 nm吸光度下降速率; 来测定NADPH脱氢速率, 从而计算GR活性.
丙二醛(MDA)的测定采用可见分光光度法, MDA与硫代巴比妥酸缩合, 生成红色产物, 在532 nm有最大吸收峰; 同时测定600 nm下的吸光度, 利用532 nm与600 nm下的吸光度的差值计算MDA的含量.
2 结果与讨论 2.1 氨氮浓度对不同藻株生长的影响一般而言, 氨氮易于被藻细胞吸收利用, 对氨氮的利用优于硝态氮和亚硝氮[18, 19].利用正常培养基和不同氨氮浓度50、500和2 000 mg ·L-1培养衣藻10 d后[图 1(a)与图 2], 生物量由初始的0.17 g ·L-1分别增加至1.31、0.8、0.73和0.71 g ·L-1.不同浓度下, 各组生物量的变化随着时间的延长而增加.但是, 相比正常培养基, 不同浓度的氨氮处理都不同程度地抑制了衣藻的生长, 且随着氨氮浓度的增大和培养时间的延长, 它对衣藻的抑制作用也越来越大.在培养的前4 d, 衣藻在不同浓度氨氮和SE+TAP培养基中生长状况相近, 但在第6~10 d, 在SE+TAP培养基中衣藻的生长状况相比其他处理组生长较好.在添加不同浓度NH4Cl的培养基中, 微藻以较为缓慢趋势生长, 尤其在500 mg ·L-1和2 000 mg ·L-1的氨氮浓度下藻细胞生长最为缓慢.相对而言, 50 mg ·L-1下衣藻的生长相对较好.一定的氨氮浓度条件下微藻能较好地生长, 过低的氨氮浓度导致营养不足, 无法满足微藻的生长代谢[20], 而过高的氨氮浓度则会产生较多铵离子, 从而无法迅速转移到氨基酸的合成中, 影响微藻的生长甚至死亡[21].或许50 mg ·L-1的浓度含量较低, 而其他的氨氮浓度超出了衣藻的耐受范围, 影响了藻细胞的生长.

|
图 1 不同浓度氨氮下不同藻株的生物量 Fig. 1 Biomass of different algae under different concentrations of ammonia nitrogen |

|
“Normal”指正常培养基, 下同 图 2 不同浓度氨氮下不同藻株的生物产率 Fig. 2 Biomass productivities of different algae under different concentrations of ammonia nitrogen |
从图 1(b)中可以看出, 正常培养基和不同氨氮浓度条件中(50、500和2 000 mg ·L-1), 葡萄藻由初始生物量0.16 g ·L-1分别增加至1.88、1.63、1.95和1.18 g ·L-1, 其生物产率分别为0.17、0.15、0.18和0.10 g ·(L ·d)-1(图 2).在培养前期(0~2 d), 各处理组保持良好生长, 但2 000 mg ·L-1氨氮处理后的生物量明显低于其他组.在2 000 mg ·L-1氨氮下, 葡萄藻一直保持较低增长速率, 可能是过高的氨氮浓度对葡萄藻的生长产生了一定的抑制作用[21].而50 mg ·L-1和500 mg ·L-1氨氮中葡萄藻的生物量较高, 并且500 mg ·L-1氨氮中葡萄藻的生物量超出了BG11正常培养基的值. Maldonado等[22]的研究发现, 氨氮浓度超过100 mg ·L-1的培养液会影响葡萄藻的生长代谢, 但本实验研究中当培养基中氨氮浓度为500 mg ·L-1时, 藻细胞的生长仍较好, 这或许与不同藻株的葡萄藻有关.
由图 1(c)可知, 正常培养基和不同氨氮浓度条件下(50、500和2 000 mg ·L-1), 紫球藻由初始生物量0.18 g ·L-1分别增长至1.62、1.78、1.02和0.86 g ·L-1, 相应的生物产率分别为0.14、0.16、0.08和0.07 g ·(L ·d)-1(图 2).培养前期, 各组均保持良好增长趋势, 在第4 d, 500 mg ·L-1和2 000 mg ·L-1氨氮中紫球藻的生物量明显低于正常培养基和50 mg ·L-1的氨氮处理.第4 d后, KOCK培养基和50 mg ·L-1氨氮中紫球藻的生物量较高, 且生长速率较快; 而500 mg ·L-1和2 000 mg ·L-1的氨氮浓度下紫球藻的生物量虽然保持增长状态, 但生长速率较为缓慢.因此, 紫球藻对氨氮浓度更加敏感, 过高的氨氮浓度影响了紫球藻的生长, 导致紫球藻对铵态氮的吸收能力弱[21].而50 mg ·L-1的氨氮浓度较低, 为紫球藻的生长提供了一定的营养盐, 促进了紫球藻的生物量积累.
从图 1(d)中可得知, 正常培养基和不同氨氮浓度条件中(50、500和2 000 mg ·L-1), 栅藻由初始生物量0.16 g ·L-1分别增长至2.38、2.26、2.18和1.46 g ·L-1, 其生物产率分别是0.22、0.21、0.20和0.13 g ·(L ·d)-1(图 2).在培养前6 d内, BG11、50 mg ·L-1和500 mg ·L-1氨氮中栅藻的生物量无明显差异, 而2 000 mg ·L-1的氨氮对栅藻产生了一定的抑制作用.第6 d后, BG11生物量持续增长, 50 mg ·L-1和500 mg ·L-1氨氮中栅藻的生物量略有下降, 之后增长速率与BG11基本一致. 2 000 mg ·L-1氨氮中栅藻的生物量增长较为缓慢, 可能因为2 000 mg ·L-1的氨氮浓度超出了栅藻的承受范围, 对藻细胞生长产生了抑制作用[21], 而低浓度的氨氮为栅藻的生长补充了一定的营养盐, 更有利于栅藻的生长[20].
在本实验中, 各处理组藻种的生长情况存在较大差异, 对不同氨氮浓度表现出了不同的耐受性.其中不同浓度氨氮下衣藻和栅藻的生物量均低于正常培养基的值, 受到了不同程度的抑制, 且在50 mg ·L-1和500 mg ·L-1氨氮中栅藻的生物量比BG11培养基略低, 仅在2 000 mg ·L-1氨氮中抑制作用显著; 但相对SE+TAP培养基, 所有浓度的氨氮均对衣藻的生长产生了明显的抑制作用, 表明衣藻对氨氮耐受能力较差.葡萄藻在500 mg ·L-1氨氮中所得生物量高于BG11培养基, 紫球藻在50 mg ·L-1氨氮中所得生物量高于KOCK培养基, 500 mg ·L-1氨氮近似沼液废水浓度, 似乎葡萄藻更适合处理沼液废水, 但反观4种藻类在500 mg ·L-1氨氮中的生物产率, 栅藻的生物产率为0.20 g ·(L ·d)-1, 高于葡萄藻的0.18 g ·(L ·d)-1.
2.2 氨氮浓度对不同藻株酶活性的影响植物在生命过程中不断产生活性氧, 同时又形成一个清除活性氧的防御系统, 即酶促系统和非酶促系统, 使植物体内活性氧的产生与清除之间维持在一个动态平衡状态[23].活性氧酶促系统主要有超氧化物歧化酶(SOD)、过氧化物酶(POD)和过氧化氢酶(CAT)等.这些酶活性在不同程度上反映了植物抗性的强弱.丙二醛(MAD)是膜脂质过氧化的次级产物, 其含量变化可反映细胞脂质过氧化程度和损伤程度[24].
不同氨氮浓度对各藻种藻细胞POD活性的影响如图 3(a)所示.正常培养基及50、500和2 000 mg ·L-1浓度的氨氮中葡萄藻POD活性分别为1.80、0.12、3.10和2.00 U ·mg-1; 紫球藻的POD活性分别为0.28、1.02、0.22和0.18 U ·mg-1, 随着氨氮浓度的升高, 活性呈下降的趋势, 且均低于正常培养基的值; 衣藻的POD活性分别为0.61、4.03、0.12和0.10 U ·mg-1, 活性亦呈下降的趋势; 栅藻的POD活性分别为1.60、0.19、3.28和3.14 U ·mg-1, 同葡萄藻的POD活性变化相近.综上, 本实验的藻种中紫球藻、衣藻的POD活性随着氨氮浓度的升高而降低, 而葡萄藻、栅藻的POD活性呈现先升高后降低的趋势, 可能是因为抗氧化酶的活性因藻种的不同而存在一定的差异[25].

|
图 3 不同浓度氨氮下不同藻株的酶活性及MDA含量 Fig. 3 Enzyme activity and MDA content of different algae under different concentrations of ammonia nitrogen |
超氧化物歧化酶(SOD)能消除生物体在新陈代谢过程中产生的自由基.就图 3(b)而言, 正常培养基及50、500和2 000 mg ·L-1浓度的氨氮中葡萄藻的SOD活性分别为1.55、2.66、2.00和0.88 U ·mg-1; 紫球藻的SOD活性分别为8.21、9.37、5.97和4.89 U ·mg-1; 衣藻的SOD活性分别为5.34、2.46、0.89和0.62 U ·mg-1; 栅藻的SOD活性分别为7.21、7.34、8.43和12.12 U ·mg-1.比较而言, 栅藻的SOD活性较高, 且活性随氨氮浓度的升高而升高, 但其它几种藻株的SOD活性均随着氨氮浓度的升高而降低, 这可能是因为氨氮降低了细胞代谢中去除活性氧的需求[26].
不同藻株的CAT活性如图 3(c)所示, 正常培养基及50、500和2 000 mg ·L-1浓度的氨氮中葡萄藻的CAT活性分别为627.35、242.72、102.46和40.36 U ·mg-1; 紫球藻的CAT活性分别为928. 70、612.32、473.40和344.70 U ·mg-1, 具有较高的CAT活性; 衣藻的CAT活性分别为366.06、148.74、6.15和2.32 U ·mg-1; 栅藻的CAT活性分别为86.54、114.87、300.10和128.14 U ·mg-1.栅藻的CAT活性随氨氮浓度升高先升高后降低, 500 mg ·L-1氨氮下活性最高, 而其他藻株则表现为随着氨氮浓度的升高, CAT的活性逐渐降低.
GR的活性如图 3(d)所示, 正常培养基及50、500和2 000 mg ·L-1浓度的氨氮中葡萄藻的GR活性分别为0.29、0.31、0.27和0.25 U ·mg-1; 紫球藻的GR活性分别为0.25、0.10、0.12和0.32 U ·mg-1; 衣藻的GR活性分别为0.30、0.34、0.24和0.18 U ·mg-1; 栅藻的GR活性分别为0.76、0.88、1.21和1.48 U ·mg-1, 其中葡萄藻、衣藻的GR活性随氨氮浓度升高而降低, 而紫球藻、栅藻的GR活性则随氨氮浓度的升高而升高.
当植物受到胁迫时, 随胁迫时间的延长和胁迫程度的加重, 活性氧积累得越来越多, 最终使细胞膜发生膜质过氧化并发生自由基链式反应, 形成丙二醛, 使细胞膜流动性下降, 膜功能受到伤害[27]. 图 3(e)为MDA的含量, 正常培养基及50、500和2 000 mg ·L-1浓度的氨氮中葡萄藻的MDA含量分别为0.12、0.45、0.46和0.28 U ·mg-1; 紫球藻的MDA含量分别为0.09、0.02、0.23和0.21 U ·mg-1; 衣藻的MDA含量分别为1.49、0.26、0.08和0.08 U ·mg-1; 栅藻的MDA含量分别为1.62、1.68、1.96和2.52 U ·mg-1.其中栅藻的MDA含量随氨氮浓度的升高而升高, 葡萄藻和紫球藻的MDA含量均随着氨氮浓度的升高而呈现出先升后降的趋势, 衣藻的MDA含量随氨氮浓度的升高而下降.
总体上, 随着氨氮浓度的不断升高, 栅藻的SOD、CAT和GR酶活性不断升高, POD酶活性先升高后下降, MDA含量不断升高, 可能是栅藻在受到氨氮胁迫时, 会产生大量的ROS伤害机体, 酶活性的升高表明藻细胞产生了应激性反应, 保护藻细胞免受氧化压力带来的损伤, 但POD酶活性后期又呈下降的趋势, 表明它的活性不是无限上升的, 即它对膜系统的保护是有限度的[28, 29]. MDA含量的升高可能是ROS对栅藻产生的氧化压力导致细胞膜脂质过氧化增强, 将对细胞产生不利影响[30].葡萄藻和紫球藻中多数酶活性随氨氮浓度的升高而降低, MDA含量也先升后降, 500 mg ·L-1的氨氮中MDA含量达到最大值.衣藻POD、SOD、CAT和GR这4种酶活性在不断降低, MDA含量不断减少, 表明衣藻藻细胞无法通过自身的防御系统来消除或减轻氨氮的胁迫, 以致酶活性降低, 但其MDA含量却并未升高, 表明酶活性的降低并没有造成ROS的积累.
2.3 氨氮浓度对不同藻株单位生物产率的酶活性的影响图 4为不同氨氮浓度下不同藻株单位生物产率上的酶活性, 表明氨氮胁迫对其影响是一个动态的变化过程[31].不同浓度氨氮处理衣藻时, 50 mg ·L-1氨氮下POD酶活性显著提高, 但高浓度时活性明显下降.氨氮浓度为500 mg ·L-1时, 4种藻株单位生物产率上的POD酶活性为葡萄藻>栅藻>紫球藻>衣藻; SOD酶活性为紫球藻>栅藻>衣藻>葡萄藻; CAT酶活性为紫球藻>栅藻>葡萄藻>衣藻; GR酶活性为栅藻>衣藻>紫球藻=葡萄藻; MDA含量为栅藻>紫球藻>葡萄藻>衣藻.几种酶活性未表现出同步性, 可能是因为他们在功能上具有相对独立性[28].

|
图 4 不同浓度氨氮下不同藻株单位生物产率上的酶活性及MDA含量 Fig. 4 Enzyme activity and MDA content in the biomass productivity of different algae under different concentrations of ammonia nitrogen |
藻细胞对高氨氮浓度具有一定的抗逆性能和适应能力.但长时间高浓度氨氮环境下, 藻类可能由于体内的碳水化合物已逐渐消耗殆尽, 从而可能使得高浓度氨氮对藻类产生抑制作用, 因而无法维持酶的响应活力, 从而导致酶活性下降[32].
3 结论(1) 不同氨氮浓度50、500和2 000 mg ·L-1均不同程度抑制了衣藻和栅藻的生长, 50 mg ·L-1的低浓度氨氮微藻生长较好, 紫球藻生长状况最佳, 其生物量为1.78 g ·L-1, 生物产率为0.16 g ·(L ·d)-1; 在500 mg ·L-1氨氮中葡萄藻的生长状况最佳, 其生物量为1.95 g ·L-1, 生物产率为0.18 g ·(L ·d)-1; 而栅藻在500 mg ·L-1氨氮中的生物量为2.18 g ·L-1, 生物产率为0.20 g ·(L ·d)-1.因此, 可进一步研究葡萄藻和栅藻处理沼液废水的效果.
(2) 葡萄藻的SOD、CAT和GR等酶活性随氨氮浓度的升高而升高, POD酶活性先升后降, 但MDA含量也在升高; 衣藻的SOD、POD、CAT和GR活性均表现为随氨氮浓度的升高, 活性下降, MDA亦然.
| [1] | Deng X Y, Gao K, Zhang R C, et al. Growing Chlorella vulgaris on thermophilic anaerobic digestion swine manure for nutrient removal and biomass production[J]. Bioresource Technology, 2017, 243: 417-425. DOI:10.1016/j.biortech.2017.06.141 |
| [2] |
顾建辉, 李彩珍, 黄天寅, 等. 分段进水SBR处理高浓度氨氮废水的中试研究[J]. 中国给水排水, 2018, 34(17): 104-107. Gu J H, Li C Z, Huang T Y, et al. Treatment of high-concentration ammonia nitrogen wastewater by a pilot step-feed SBR[J]. China Water & Wastewater, 2018, 34(17): 104-107. |
| [3] |
卫丹, 万梅, 刘锐, 等. 嘉兴市规模化养猪场沼液水质调查研究[J]. 环境科学, 2014, 35(7): 2650-2657. Wei D, Wan M, Liu R, et al. Study on the quality of digested piggery wastewater in large-scale farms in Jiaxing[J]. Environmental Science, 2014, 35(7): 2650-2657. |
| [4] |
姜奕圻, 廖若莹, 黄意淇, 等. 利用沼液养殖微藻研究进展[J]. 中国沼气, 2017, 35(4): 36-42. Jiang Y Q, Liao R Y, Huang Y Q, et al. Progress of microalgae cultivation in liquid digestate[J]. China Biogas, 2017, 35(4): 36-42. DOI:10.3969/j.issn.1000-1166.2017.04.007 |
| [5] |
王愿珠, 程鹏飞, 刘德富, 等. 生物膜贴壁培养小球藻净化猪粪沼液废水的效果[J]. 环境科学, 2017, 38(8): 3354-3361. Wang Y Z, Cheng P F, Liu D F, et al. Purification effect of piggery wastewater with Chlorella pyrenoidosa by immobilized biofilm-attached culture[J]. Environmental Science, 2017, 38(8): 3354-3361. |
| [6] |
程鹏飞, 王艳, 杨期勇, 等. 微藻贴壁培养对沼液废水的处理效果[J]. 浙江农业学报, 2015, 29(9): 1564-1569. Cheng P F, Wang Y, Yang Q Y, et al. Purification effect for swine wastewater with attached culture of microalgae[J]. Acta Agriculturae Zhejiangensis, 2015, 29(9): 1564-1569. |
| [7] | Fazal T, Mushtaq A, Rehman F, et al. Bioremediation of textile wastewater and successive biodiesel production using microalgae[J]. Renewable and Sustainable Energy Reviews, 2018, 82: 3107-3126. DOI:10.1016/j.rser.2017.10.029 |
| [8] | 吴买生. 推广发酵床生态养猪技术实现生猪产业可持续发展[J]. 湖南畜牧兽医, 2009(5): 27-29. DOI:10.3969/j.issn.1006-4907.2009.05.015 |
| [9] | Ndikubwimana T, Zeng X H, Murwanashyaka T, et al. Harvesting of freshwater microalgae with microbial bioflocculant:a pilot-scale study[J]. Biotechnology for Biofuels, 2016, 9: 47. DOI:10.1186/s13068-016-0458-5 |
| [10] | 刘刈, 宋立, 邓良伟. 我国规模化养殖场粪便污水处理利用现状及对策[J]. 猪业科学, 2011, 28(6): 30-33, 42. DOI:10.3969/j.issn.1673-5358.2011.06.023 |
| [11] | Dai G Z, Qiu B S, Forchhammer K. Ammonium tolerance in the cyanobacterium Synechocystis sp. strain PCC 6803 and the role of the psbA multigene family[J]. Plant, Cell & Environment, 2014, 37(4): 840-851. |
| [12] |
孙玉焕, 吴友浩, 杨志海. 初始氨氮浓度对钝顶螺旋藻生长及其去除率的影响[J]. 环境保护科学, 2012, 38(4): 33-36. Sun Y H, Wu Y H, Yang Z H. Effect of different initial ammonia nitrogen concentrationon on spirulina platensis growth and ammonia nitrogen removal[J]. Environmental Protection Science, 2012, 38(4): 33-36. DOI:10.3969/j.issn.1004-6216.2012.04.009 |
| [13] | Converti A, Casazza A A, Ortiz E Y, et al. Effect of temperature and nitrogen concentration on the growth and lipid content of Nannochloropsis oculata and Chlorella vulgaris for biodiesel production[J]. Chemical Engineering and Processing:Process Intensification, 2009, 48(6): 1146-1151. DOI:10.1016/j.cep.2009.03.006 |
| [14] | Rippka R, Deruelles J, Waterbury J B, et al. Generic assignments, strain histories and properties of pure cultures of cyanobacteria[J]. Journal of General Microbiology, 1979, 111(6): 1-61. |
| [15] | Miao X L, Wu Q Y. Biodiesel production from heterotrophic microalgal oil[J]. Bioresource Technology, 2006, 97(6): 841-846. DOI:10.1016/j.biortech.2005.04.008 |
| [16] | Liu Z Y, Wang G C, Zhou B C. Effect of iron on growth and lipid accumulation in Chlorella vulgaris[J]. Bioresource Technology, 2008, 99(11): 4717-4722. DOI:10.1016/j.biortech.2007.09.073 |
| [17] | 常功法, 毕学军.高浓度氨氮废水处理[A].全国高浓度有机废水处理及工程应用新技术、新设备交流研讨会论文集[C].北京: 中国高科技产业化研究会, 2011. |
| [18] |
卞少伟, 武丹, 方普杰, 等. 天津近岸海域冬季浮游植物群落结构特征[J]. 环境影响评价, 2015, 37(3): 77-80. Bian S W, Wu D, Fang P J, et al. Community structure of phytoplankton in tianjin offshore waters in winter[J]. Environmental Impact Assessment, 2015, 37(3): 77-80. |
| [19] | Lampitt R S, Wishner K F, Turley C M, et al. Marine snow studies in the Northeast Atlantic Ocean:distribution, composition and role as a food source for migrating plankton[J]. Marine Biology, 1993, 116(4): 689-702. DOI:10.1007/BF00355486 |
| [20] |
蒋汉明, 高坤山. 氮源及其浓度对三角褐指藻生长和脂肪酸组成的影响[J]. 水生生物学报, 2004, 28(5): 545-551. Jiang H M, Gao K S. Effects of nitrogen sources and concentrations on the growth and fatty acid composition of Phaeoda-ctylum tricornutum[J]. Acta Hydrobiologica Sinica, 2004, 28(5): 545-551. DOI:10.3321/j.issn:1000-3207.2004.05.015 |
| [21] |
王璐瑶, 桑敏, 李爱芬, 等. 不同缺氮营养水平对金色奥杜藻生长及光合生理的影响[J]. 中国生物工程杂志, 2012, 32(6): 48-56. Wang L Y, Sang M, Li A F, et al. Effects of different nitrogen nutrition level on the growth and photosynthetic physiology of Odontella aurita[J]. China Biotechnology, 2012, 32(6): 48-56. |
| [22] | Maldonado J G G, Bastoul D, Baig S, et al. Effect of solid characteristics on hydrodynamic and mass transfer in a fixed bed reactor operating in co-current gas-liquid up flow[J]. Chemical Engineering and Processing:Process Intensification, 2008, 47(8): 1190-1200. DOI:10.1016/j.cep.2007.02.013 |
| [23] |
于娟, 唐学玺, 张培玉, 等. CO2加富对两种海洋微绿藻的生长、光合作用和抗氧化酶活性的影响[J]. 生态学报, 2005, 25(2): 197-202. Yu J, Tang X X, Zhang P Y, et al. Effects of CO2 enrichment on growth, photosynthesis and activities of antioxidant enzymes of two marine micro-green-algae[J]. Acta Ecologica Sinica, 2005, 25(2): 197-202. |
| [24] | 秦洪伟, 陈柳芳, 鲁楠, 等. 氧氟沙星对斜生栅藻的毒性效应[J]. 环境化学, 2011, 30(4): 885-886. |
| [25] |
焉翠蔚, 肖宜华, 朱岩松, 等. 多效唑对2种海洋微藻生长和抗氧化酶活性的影响[J]. 中国海洋大学学报, 2008, 38(2): 291-296. Yan C W, Xiao Y H, Zhu Y S, et al. Effects of PP333 on growth and activities of antioxidative enzymes of two marine microalgae[J]. Periodical of Ocean University of China, 2008, 38(2): 291-296. |
| [26] |
彭金良, 严国安, 沈国兴, 等. α-萘酚胁迫对普通小球藻生长及抗氧化酶活性的影响[J]. 武汉大学学报(理学版), 2001, 47(4): 449-452. Peng J L, Yan G A, Shen G X, et al. The effects of α-naphthol on the growth and the antioxidase activities of Chlorella vulgaris[J]. Journal of Wuhan University (Natural Science Edition), 2001, 47(4): 449-452. DOI:10.3321/j.issn:1671-8836.2001.04.016 |
| [27] |
任红旭, 陈雄, 吴冬秀. CO2浓度升高对干旱胁迫下蚕豆光合作用和抗氧化能力的影响[J]. 作物学报, 2001, 27(6): 729-736. Ren H X, Chen X, Wu D X. Effects of elevated CO2 on photosynthesis and antioxidative ability of broad bean plants grown under drought condition[J]. Acta Agronomica Sinica, 2001, 27(6): 729-736. DOI:10.3321/j.issn:0496-3490.2001.06.008 |
| [28] |
王丽平, 郑丙辉, 孟伟. 荧蒽对两种海洋硅藻生长、SOD活力和MDA含量的影响[J]. 海洋通报, 2008, 27(4): 53-58. Wang L P, Zheng B H, Meng W. Effects of fluoranthene on growth, SOD activity and MDA content of two marine diatoms[J]. Marine Science Bulletin, 2008, 27(4): 53-58. DOI:10.3969/j.issn.1001-6392.2008.04.007 |
| [29] |
陈淑芳, 郭沛涌, 阮滨. 稳定性二氧化氯对水华微囊藻叶绿素a及酶活性的影响[J]. 中南大学学报(自然科学版), 2016, 47(2): 414-419. Chen S F, Guo P Y, Ruan B. Effects of stable chlorine dioxide on chlorophyll a and antioxiant enzymes of Microcystis flos-aquae[J]. Journal of Central South University (Science and Technology), 2016, 47(2): 414-419. |
| [30] |
杨国远, 万凌琳, 雷学青, 等. 重金属铅、铬胁迫对斜生栅藻的生长、光合性能及抗氧化系统的影响[J]. 环境科学学报, 2014, 34(6): 1606-1614. Yang G Y, Wan L L, Lei X Q, et al. Effects of lead and chromium on the growth, photosynthetic performance, and antioxidant activity of Scenedesmus obliquus[J]. Acta Scientiae Circumstantiae, 2014, 34(6): 1606-1614. |
| [31] |
赖廷和, 何斌源, 范航清, 等. 重金属Cd胁迫对红树蚬的抗氧化酶、消化酶活性和MDA含量的影响[J]. 生态学报, 2011, 31(11): 3044-3053. Lai T H, He B Y, Fan H Q, et al. Effects of cadmium stress on the activities of antioxidant enzymes, digestive enzymes and the membrane lipid peroxidation of the mangrove mud clam Geloina coaxans (Gmelin)[J]. Acta Ecologica Sinica, 2011, 31(11): 3044-3053. |
| [32] |
颜昌宙, 曾阿妍, 金相灿, 等. 不同浓度氨氮对轮叶黑藻的生理影响[J]. 生态学报, 2007, 27(3): 1050-1055. Yan C Z, Zeng A Y, Jin X C, et al. Physiological effects of ammonia-nitrogen concentrations on Hydrilla verticillata[J]. Acta Ecologica Sinica, 2007, 27(3): 1050-1055. DOI:10.3321/j.issn:1000-0933.2007.03.027 |