 2019, Vol. 40
2019, Vol. 40



2. 三峡大学水利与环境学院, 三峡库区生态环境教育部工程中心, 宜昌 443002
2. Engineering Research Center of Eco-environment in Three Gorges Reservoir Region, Ministry of Education, College of Hydraulic and Environmental Engineering, China Three Gorges University, Yichang 443002, China
三峡水库自2003年蓄水成库以来因改变了库区的水文和水动力情势导致支流水华频发, 对库区人民日常生活、工农业用水及水库的正常运行带来不利影响, 成为国内外广泛关注的生态环境问题之一[1~4].
从藻类生态学角度来看, 水动力条件是影响藻类生长的关键因素[5~8].一方面水动力条件通过影响藻类细胞的分裂、形态、光合特性、光合色素的组成以及藻细胞的生存能力来影响藻类的生长, 另一方面通过直接影响藻细胞周围的环境(例如:光照强度、水温、营养盐和浮游动物分布)间接影响藻类的生长.相关学者和科研团队在三峡水库香溪河野外原位观测发现:降雨的发生总是伴随着水华现象的快速消退[9, 10].综合目前三峡水库支流水华生消过程和机制的研究成果认为“三峡水库的水华问题从本质上讲是因为大坝建设导致的生境条件的演变, 这种演变的核心是水动力背景的改变”.李锦秀等[11]基于监测数据构建了大宁河支流流速与藻类水华关系曲线并提出了临界流速的概念, 认为当支流流速小于临界流速时, 暴发水华现象, 反之, 水华现象消失.曹巧丽等[12]基于室内控制实验指出当室内控制流速大于1 m·s-1时仍然暴发水华, 即在三峡水库支流决定水华消失的流速不存在.徐耀阳[13]基于三峡水库香溪河现场跟踪监测数据指出水体滞留时间越长, 藻类越容易生长聚集从而暴发水华.但是郑丙辉等[14]在2007年12月~2008年1月之间, 在大宁河河段监测到了严重的微囊藻水华现象, 按照前期研究结果, 此时水体滞留时间最长, 水温最低, 从理论上讲不会暴发水华, 因此水体滞留时间并不是影响水华暴发的主控因素.本团队基于近10年的香溪河野外原位观测数据, 运用微弱流场监测技术[15], 发现在三峡水库支流普遍存在分层异重流现象, 同时运用临界深度理论和中度扰动理论, 指出分层异重流所导致的真光层/混合层的变化是影响水华生消的关键因素.本团队通过野外跟踪监测、室内外控制实验和数值模拟等方法开展了降雨对水华消退的影响机制研究.结合其他科研团队在三峡水华生消机制方面的研究结果, 推测降雨所带来的三峡水库支流水动力条件的改变可能是导致降雨期间水华快速消失的原因.纪道斌等[16]基于香溪河暴雨过程中监测数据指出:降雨对水华存在着抑制和促进的双重作用, 降雨通过洪水的冲刷作用对藻类生物量产生稀释作用, 降低藻类生物量浓度; 暴雨冲刷过程中所携带的泥沙颗粒增加了水体中悬浮物浓度, 透明度减少, 导致真光层深度降低, 抑制藻类生长; 降雨增大了水体的垂向掺混强度, 水体表层藻类在水体剪切力的作用下进入中下层, 抑制了水华的发生; 但是, 降雨导致水体温度梯度增大, 混合层减小, 则为日后的水华的暴发提供了适宜条件.崔玉洁[10]基于降雨期间香溪河库湾跟踪监测和数值模拟结果指出:在香溪河库湾分层异重流这种特殊的水动力条件下, 降雨通过增大水体的垂向掺混强度导致藻类的原位沉降作用加强, 表层水华现象消失.高圻烽等[17]基于香溪河干支流水动力和藻类生长耦合模拟结果指出, 水体的垂向掺混强度对于叶绿素浓度和藻类生长具有重要影响, 水体垂向掺混加强抑制了藻类富集作用, 同时通过水体的垂向掺混过程将藻类带入水体真光层深度以下, 抑制藻类的生长, 导致叶绿素浓度降低.基于以上科研成果笔者推测:降雨一方面通过影响香溪河水体垂向掺混强度来直接影响藻类的生长, 另一方面通过影响水体的营养盐浓度、水下光热结构(例如:光混比)来间接影响藻类的生长.但是降雨是通过影响哪些水动力参数来影响藻类的生长过程尚不明晰, 同时水华的暴发是一个藻类演替、快速增殖和上浮聚集到水体表层为肉眼所见的生态学过程, 如果仅仅用叶绿素浓度或者藻类细胞密度这些瞬时值无法准确地表征水华的生消过程.藻类比生长速率通常是指特性藻种在天然水域进行原位培养所得到的生长速率, 因此藻类原位生长实验能有效揭示藻类在特定水域中实际生长特征与生态学行为.李哲[18]通过在三峡水库彭溪河支流开展主要藻种比生长速率的初步研究, 设计适用于三峡水库支流的藻类原位生长培养装置.同时冯婧等[19]在三峡水库彭溪河支流比生长速率及其影响因素的研究结果表明:三峡这种特殊的水动力条件一方面通过改变流速的大小来影响主要藻种的比生长速率; 另一方面通过影响水温、营养盐浓度和藻类的漂移来影响藻类的比生长速率.表明藻类原位生长实验能较好地揭示降雨所带来的水动力条件的改变对水华生消过程的影响.
因此本研究在香溪河库湾选取不同生长策略的3种主要藻种进行原位生长实验, 分析降雨期与非降雨期对藻类比生长速率的变化特征, 综合运用数理统计分析方法和生态学理论分析影响藻类比生长速率的主控环境因素, 以期为三峡水库水华生消机制理论体系的构建提供理论依据.
1 材料与方法 1.1 研究区域概述香溪河为三峡水库第一条一级支流, 位于荆山山脉与巫山山脉之间(110°25′~111°06′E, 30°57′~31°34′N), 干流全长94 km.该流域属亚热带大陆性季风气候, 年降雨量900~1 200 mm, 夏季炎热多伏旱, 以暴雨为主; 秋季阴冷潮湿, 以阴雨为主.因此本研究时间选为水华多暴发的10月, 地点选为香溪河野外环境观测站监测平台(图 1), 三峡库区水位及入库流量变化过程见图 2.

|
图 1 香溪河回水区示意 Fig. 1 Backwater area of the Xiangxi River |

|
图 2 三峡库区水位及入库流量变化过程 Fig. 2 Process of water level and inflow discharge in Three Gorges Reservoir area |
整个实验装置设计在野外监测平台支架上.通过钢圈和目绢丝制成直径为0.3 m, 高度为5 m的原位培养笼, 将制成的培养笼悬挂于支架上并高出水面0.5 m以免围格外浮游动物、原有藻种等进入, 影响整个实验结果.因培养笼装置内衬钢圈, 故除去浮游动物和原有藻种的原水可透过目绢丝进入培养笼内部.实验共设计3组原位培养笼并列悬挂于支架上, 每组培养笼内的水体营养条件、环境条件及水动力条件均与外界原水相同.本实验装置示意如图 3所示, 围格设计标准如表 1.

|
图 3 培养装置示意 Fig. 3 Map of the two incubation devices |
|
|
表 1 野外原位实验设计 Table 1 Setup of field in situ experiment |
1.3 实验方法
本研究时间选在营养盐充足、水温适宜且气象多变的汛末蓄水期, 即10月4~18日, 周期为15 d.培养对象依据藻类形态功能组[20] (morphologically-based functional groups, MBFG), 选出3种香溪河水库主要藻种:铜绿微囊藻(Microcystis aeruginosa)、小球藻(Chlorella)和栅藻(Scenedesmus), 藻种均来源于中国科学院水生生物研究所专业技术人员扩培(编号分别为:FACHB-646、FACHB-8和FACHB-489), 各种藻细胞密度在1×107 cell·L-1.将扩培后的藻种放入原位培养笼后, 每隔一天早上, 通过采水器分别在围格内的表层和底层取250 mL水样, 每取一次藻种都用蒸馏水将采水器进行冲洗, 以免影响其它围格内藻种的生长, 将采集后的水样带回实验室做水质分析.
本研究选择以叶绿素a(Chl-a)浓度来衡量藻类生物的指标, 水柱里Chl-a浓度计算参照Parsons计算公式[21]:由于每一监测点Chl-a的垂直分布是深度的函数:Chl-a=f(z), 对其进行积分, 将得到新的Chlt:
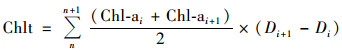
|
(1) |
式中, Chlt为水柱Chl-a浓度(mg·m-2); Chl-ai为第i层Chl-a浓度(mg·m-3); n为采样层数; Di为第i层采样深度(m); 1≤i≤n-1.
由Parsons的计算公式, 可以进一步定义叶绿素总量计算公式:
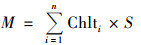
|
(2) |
式中, M为水柱叶绿素总量(mg); Chlti为水柱中第i层叶绿素a浓度(mg·m-2); n为采样层数; S为围格截面积(m2); 1≤i≤n-1.
本研究中所获得的藻类原位生长速率为在培养装置中的表观生长速率[22]:
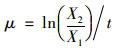
|
(3) |
式中, μ为藻类比生长速率(d-1); Xi为i时刻的Chl-a浓度(mg·L-1); t为培养天数(d).
现场主要监测指标有:水温、相对湿度、气温、pH值、溶解氧、电导率、气压、光照强度等.其中水温、pH值、溶解氧、电导率等参数由Hydrolab DS5多参仪(美国)现场测定; 光照强度(PAR)由LI-1400水下光量子仪(美国)测定; 湿度、气温由室内温湿度计测定.
室内主要监测指标有:叶绿素a(Chl-a)、总氮(TN)、氨氮(NH4+-N)、硝氮(NO3--N)、总磷(TP)、正磷酸盐(PO43-)等营养盐浓度.所有室内化学指标分析方法参照文献[23]来进行测定.
1.4 水体垂向掺混强度各具体参数紊动黏性系数[24]是Boussinesq于1887年首次提出的, 其是针对流体处于紊动状态时脉动场的对流输运对整个时均场的影响, 因此紊动黏性系数是联系紊流脉动场和时均场的一个重要的物理参数, 分别包括垂向紊流黏性系数和垂向紊流扩散系数, 反映了水体在垂向上的动能交换能力和扩散的运输能力.需要指出的是, 在不同的相关模型中, 受到边界条件、研究对象等诸多因素的影响时, 紊动黏性系数的取值不尽相同, 只能给出一个大致范围, 甚至同一研究区域的不同时段也可能有所差异.本文采用笛卡尔坐标系下的三维数字模型基本方程, 由以下公式表述[25]:
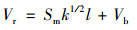
|
(4) |
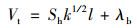
|
(5) |
式中, Vr和Vt表示垂向紊流黏性系数和垂向紊流扩散系数(m2·s-1); Vb和λb表示背景混合系数, 一般取10-6 m2·s-1; k为紊动能; l为混合长; Sm和Sh为稳态函数, 由下列求解:
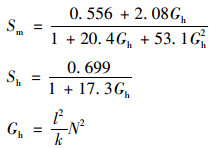
|
雷诺数Re是一种可以表征流体流动情况的无量纲数. Re < 2 000为层流状态, Re>4 000为紊流状态.紊流状态的特征是速度、压力等运动要素在时空上具有随机性, 且速度存在纵向和垂向的脉动速度, 是一种非定常流.雷诺数[26]主要由下式推导得出:
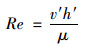
|
(6) |
式中, v′表示相同流向水层的平均流速(m·s-1); h′表示相同流向水层的厚度(m); μ为运动黏性系数, 取1.007 4×10-6 m2·s-1.
在紊流过程中, 因质点从某层流进入相邻的另一层流时会产生能量交换, 其动量发生改变, 引起切应力τ.紊流流体的剪切力τ由黏性剪切力τu和紊流剪切力τt构成, 即:
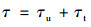
|
(7) |
紊流时, 一般τu可忽略不计, 即[27]:
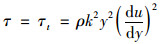
|
(8) |
式中, τ为剪切力(N·m-2); k为卡门常数, k为0.4~0.42; y取水深(m).
2 结果与分析 2.1 研究期间香溪河环境参数特征 2.1.1 研究期间香溪河水文气象特征如图 4所示, 受降雨影响, 降雨量、水温、光照强度、真光层、混合层和光混比(真光层深度/混合层深度)六大气象参数在降雨期与非降雨期存在显著性差异(ANOVA, P < 0.05).如图 4(a)所示, 降雨量在降雨期变化范围为0.60~9.40 mm, 降雨量最大值出现在10月10日, 平均值为(3.70±3.00)mm, 非降雨期降雨量为0 mm, 表明降雨期降雨量对营养盐和藻类生物量有一定的稀释作用.如图 4(b)所示, 水温在降雨期变化范围为21.00~23.30℃, 平均值为(22.41±0.49)℃, 表底温差变化范围0.14~2.89℃, 平均值为(1.14±1.09)℃, 其中表底温差最大值出现在降雨量最大值出现的10月10日; 非降雨期水温变化范围为23.11~23.95℃, 平均值为(23.38±0.30)℃, 非降雨期的表底温差较小, 为0.01~0.29℃; 按照水体稳定分层判别标准(温差>1℃·m-1, 水体形成稳定分层; 0.2℃·m-1<温差 < 1℃·m-1, 水体为弱分层; 温差 < 0.2℃·m-1, 水体为完全混合水体), 降雨期出现了稳定分层现象, 非降雨期为完全混合水体.如图 4(c)所示, 光照强度在降雨期变化范围为5.54~18.29 μmol·(m2·s)-1, 平均值为(11.92±4.41) μmol·(m2·s)-1, 非降雨期光照强度变化范围为23.50~59.37μmol·(m2·s)-1, 平均值为(41.44±25.36) μmol·(m2·s)-1.如图 4(d)所示, 真光层在降雨期变化范围为3.41~4.81 m, 平均值为(3.68±0.55)m, 非降雨期的变化范围为5.93~7.34 m, 平均值为(6.64±0.99)m; 混合层在降雨期的变化范围为5.04~25.74 m, 平均值为(9.69±8.05)m, 非降雨期的变化范围为1.80~2.32 m, 平均值为(2.06±0.37)m; 光混比在降雨期变化范围为0.14~0.93, 平均值为0.54±0.28, 非降雨期变化范围为3.17~3.30, 平均值为3.23±0.1.

|
10-06~10-08为晴天, 其他监测时间段均有持续小雨(降雨量均 < 10 mm), 下同 图 4 研究期间香溪河气象参数特征 Fig. 4 Characteristics of meteorological parameters in the Xiangxi River during the research period |
如图 5所示, 表征水体垂向掺混强度的4个水动力学参数表现出表层<中层<底层的垂向分布趋势, 表明受降雨影响, 培养装置内下层水体对上层水体有很强的黏滞、拉拽作用, 使得上层水体反向运动形成环流, 增强水体上下的交换和掺混; 因此培养装置内下层水体剪切力τ、垂向紊流黏性系数Vr和垂向紊流扩散系数Vt在降雨期显著大于非降雨期(ANOVA, P < 0.05).如图 5(a)所示, 下层水体垂向紊流黏性系数Vr在降雨期变化范围为1.11×10-3~2.18×10-3 m2·s-1, 平均值为(1.71×10-3±4.55×10-4)m2·s-1; 非降雨期垂向紊流黏性系数Vr变化范围为8.13×10-4~9.20×10-4 m2·s-1, 平均值为(8.67×10-4±7.60×10-5)m2·s-1.如图 5(b)所示, 垂向紊流扩散系数Vt在降雨期变化范围为1.86×10-3~3.20×10-3 m2·s-1, 平均值为(2.46×10-3±6.43×10-4)m2·s-1; 非降雨期垂向紊流扩散系数Vt范围为1.16×10-3~1.28×10-3 m2·s-1, 平均值为(1.22×10-3±8.48×10-5) m2·s-1.如图 5(c)所示, 下层水体剪切力τ在降雨期变化范围为8.31~114.51 N·m-2, 最大值出现在10月10日, 平均值为(35.12±40.61)N·m-2, 表明降雨对下层水体剪切力影响较为明显.

|
图 5 实验期间香溪河水动力参数特征 Fig. 5 Characteristics of hydrodynamic parameters in the Xiangxi River during the research period |
如表 2所示:研究期间pH、TN、DTN、在降雨期与非降雨期存在显著差异(ANOVA, P<0.05);电导率、溶解氧、NO3--N、NH4+-N、TP、DTP、PO43-和SI等营养盐浓度不存在显著差异(ANOVA, P<0.05); pH值在非降雨期的平均值为7.95±0.04, 为弱碱性水体, 受到降雨稀释作用的影响, 降雨期pH略有升高, 其平均值为8.07±0.03. TN与DTN在非降雨期的平均值分别为(2.19±0.08)mg·L-1和(2.09±0.08)mg·L-1, 受到降雨稀释的影响, 降雨期TN和DTN略有下降, 其平均值分别为(2.05±2.58)mg·L-1和(1.94±0.09)mg·L-1.
|
|
表 2 香溪河研究期间理化环境参数特征 Table 2 Characteristics of physical and chemistry environmental parameters in the Xiangxi River during the research period |
2.2 藻种生物量及比生长速率变化趋势
如图 6所示, 微囊藻Chl-a总量在非降雨期和降雨期存在极显著差异(ANOVA, P < 0.01);非降雨期Chl-a总量上升, 降雨期Chl-a总量下降, 表明降雨对培养装置水柱中微囊藻表层的生物量有明显的抑制作用, 而对小球藻和栅藻的影响较弱.

|
图 6 研究期间原位培养装置内Chl-a总量变化特征 Fig. 6 Characteristics of the total amount Chl-a in in situ culture apparatus during the research period |
如图 7所示:微囊藻、小球藻比生长速率在非降雨期和降雨期存在极显著差异(ANOVA, P < 0.01);栅藻比生长速率在非降雨期和降雨期存在显著差异(ANOVA, P < 0.05).降雨期和非降雨期微囊藻、小球藻和栅藻比生长速率变化范围分别为:(-0.22±0.22)~(0.51±0.20)、(-0.19±0.14)~(0.48±0.20)和(-0.15±0.25)~(0.39±0.23)d-1; 降雨期3种藻的比生长速率多为负值, 而非降雨期3种藻的比生长速率均为正值, 表明降雨对3种藻的原位生长均有明显的抑制作用.

|
图 7 研究期间培养装置内比生长速率变化特征 Fig. 7 Characteristics of the specific growth rate in culture apparatus during the research period |
相关分析表明, 表征垂向掺混的4个水动力参数与3种主要藻种比生长速率呈显著/极显著负相关关系(rmic-Vr=-0.881, P<0.01; rmic-Vt=-0.886, P<0.01; rmic-τ=-0.751, P<0.05; rmic-Re=-0.850, P<0.05; rChl-Vr=-0.824, P<0.05; rChl-Vt=-0.799, P<0.05; rChl-τ=-0.738, P<0.05; rChl-Re=-0.717, P<0.05; rsce-Vt=-0.852, P<0.01; rsce-Re=-0.780, P<0.05).光混比与3种藻种比生长速率呈显著/极显著正相关(rmic=0.873, P<0.01; rChl=0.868, P<0.01; rsce=0.687, P<0.01).降雨量与3种藻种比生长速率呈显著负相关关系(rmic=-0.818, rChl=-0.897, rsce=-0.806, P<0.05). WT与3种藻种比生长速率呈显著/极显著正相关关系(rmic=0.844, P<0.01; rChl=0.828, P<0.05; rsce=0.738, P<0.05). TN与3种藻种比生长速率呈显著/极显著正相关关系(rmic=0.707, P<0.05; rChl=0.840, P<0.01; rsce=0.739, P<0.05).微囊藻比生长速率和混合层、气压、pH呈显著负相关关系(r混合层=-0.710, r气压=-0.698, rpH=-0.766, P<0.05);小球藻比生长速率和光强、气温、DTN呈显著正相关关系(r光强=0.910, r气温=0.743, rDTN=0.722, P<0.05);栅藻比生长速率和光强呈显著正相关关系(r光强=0.813, P<0.05).
相关分析表明, 降雨量与3种藻种的Chl-a浓度呈极显著正相关关系(rmic=0.969, rChl=0.967, rsce=0.825, P<0.01).微囊藻、小球藻和栅藻Chl-a浓度均与NO3--N呈显著负相关关系(rmic=-0.760, rChl=-0.706, rsce=-0.622, P<0.05).微囊藻Chl-a浓度与PO43-呈显著负相关关系(r=-0.661, P<0.05).小球藻Chl-a浓度和SPC呈显著正相关关系(r=0.629, P<0.05).
综合前期的气象条件, 常规理化参数, 3种藻的Chl-a总量和比生长速率的变化特征及相关分析表明, 降雨通过降雨量的增加对藻的生长起到一个稀释作用, 抑制了藻的生长; 降雨通过增加水体垂向掺混强度, 一方面使得原位垂向掺混作用加强, 致使藻类生物量降低, 另一方面, 通过上下水体交换抑制了藻类生长; 降雨影响了混合层的深度, 光混比降低, 抑制了藻类的生长.
3 讨论 3.1 水动力条件对藻类影响的机制三峡水库和香溪河支流降雨前后现状跟踪监测和数值模拟表明:降雨明显增加了干流水体[10, 16]从表层倒灌入香溪河库湾与上游受冲刷水体从中层潜入水底, 这两股水流对藻类生物量有明显的稀释作用; 降雨对水动力条件的改变增加了水体的剪切力, 产生垂向掺混, 一方面水体垂向掺混强度会对藻细胞造成一定的机械损伤[24], 促使部分藻细胞死亡; 另一方面水体垂向掺混增加上下水层的交换强度, 将水体表层的藻细胞带入光照强度较弱不适宜藻类生长的水体中下层, 抑制了藻类的表层繁殖, 降低水柱中藻总的生物量.本研究结果表明:降雨带来的水动力条件的改变对藻类生长有一定的抑制作用, 剪切力τ、垂向紊流黏性系数Vr和垂向紊流扩散系数Vt在降雨期显著大于非降雨期(ANOVA, P < 0.05), 降雨中剪切力τ、垂向紊流黏性系数Vr和垂向紊流扩散系数Vt的改变是导致藻类迅速衰亡的关键因素, 这3个水动力参数的改变增加了水体的垂向掺混, 对表层水体有很强的黏滞和拉拽作用, 从而使得表层藻类被带入光照强度低的水体下层, 受到光强抑制的效果, 从而藻类表层繁殖受到抑制.
3.2 混合层变化对藻类影响的机制Sverdrup理论一般采用真光层深度(Zeu)和混合层深度(Zmix)之比(即光混比)来判别光照在垂向的分布和水体在垂向的掺混对浮游植物生长的影响[28].当真光层深度大于混合层深度(即Zeu/Zmix>1)时, 浮游植物生长不受光照的限制; 当Zeu/Zmix<1时, 浮游植物会因垂向掺混被动地带入真光层[29, 30]以下, 从而造成其生长受到光限制, 为此Sverdrup建立了临界层理论(Critical Depth Theory).本团队在前期香溪河现场监测结果表明[16, 31]:大、中型降雨导致水体真光层降低, 而会使混合层深度增加.水体混合层的加深, 水温分层减弱, 是水华消失的根本原因[32].本研究结果表明:降雨前(10月6~8日), 研究期间培养装置内混合层深度都较低(1~2m); 降雨导致真光层深度降低, 混合层深度显著增加, 此时Zeu/Zmix显著降低, 按照临界层理论, 抑制了藻类的生长.
3.3 环境参数对不同生存策略藻类的影响影响藻类生长的主要生境因子包括:物质基础(二氧化碳、营养盐和微量元素等)、能量要素(光照、温度等)、生物要素(浮游动物捕食等)以及水动力过程(水体掺混扰动条件)的影响[33~36].在水生态系统中, 藻类光合作用是物质循环和能量流动的基础.光照是浮游植物光合作用的必要条件, 光照强度和时间均会影响藻类初级生产力, 且不同的藻类对光的需求和适应能力也不同[37].水温作为重要的生态因子, 通过影响藻类的酶促反应和水体中营养物质的溶解度从而直接、间接地影响藻类的代谢与增殖.有研究表明, 在保持其它环境条件均适宜的情况下, 水温每上升10℃, 浮游植物代谢活动的强度将增加2倍[38], 但当温度超过最适温度时浮游植物初级生产迅速下降.而且不同藻种对温度的适应范围各有差异, 蓝藻的最适生长温度为25~35℃, 硅藻在15~35℃之间均可较好地生长, 金藻、黄藻最适水温为14~18℃, 绿藻为20~30℃[39].营养盐是构成水域生态系统的重要化学物质基础, 水中的营养盐结构、浓度会对浮游植物的生长起到重要的调控作用.主要体现在两个方面:一方面, 营养盐之间的比例会影响浮游植物整体的数量和群落结构; 另一方面, 营养盐的浓度高低, 直接会影响到浮游植物的生长和繁殖[40].降雨除改变水动力条件, 导致水体垂向掺混, 使得水体的混合层不断扩大, 生长受到抑制外.降雨还会带来水温、光照强度、气温、营养盐、气压、pH值等环境参数的改变从而影响藻类的生长[41, 42].本文依据藻类形态功能组[20]将3种藻类分为Ⅶ、Ⅳ型2组, 形态功能组Ⅶ具有无鞭毛无硅质外壳带黏质胶被的大型藻, 如微囊藻; 无鞭毛无硅质外壳无伪空泡大小藻种的小球藻、栅藻划入形态功能组Ⅳ.代表形态功能组Ⅶ的微囊藻最适光照强度为50μmol·(m2·s)-1左右, 代表形态功能组Ⅳ的小球藻和栅藻最适光照强度为50~90μmol·(m2·s)-1左右, 光照强度在降雨期平均值为(11.92±4.41) μmol·(m2·s)-1, 非降雨期平均值为(41.44±25.36) μmol·(m2·s)-1, 降雨期光照强度显著低于Ⅶ、Ⅳ型形态功能藻类所需最适光强, 因此抑制了3种主要藻类的生长, 导致3种主要藻类在研究期间呈现一致的时间变化趋势; 本研究3种主要藻类均为无鞭毛无硅质外壳藻类, 因此水体扰动较强的环境条件对其生长产生抑制作用.本研究结果表明:表征垂向掺混的4个水动力参数与3种主要藻种比生长速率呈显著/极显著负相关关系, 同时降雨期水体垂向掺混强度显著大于非降雨期, 抑制了3种主要藻类的生长, 因此3种主要藻类在研究期间呈现一致的时间变化趋势.
综上所述, 降雨一方面通过增大水体垂向掺混强度降低藻类生物量抑制水华, 另一方面通过影响光照强度抑制藻类的光合作用降低藻类生物量.
4 结论(1) 降雨带来的水动力条件的改变对藻类生长有一定的抑制作用, 其中剪切力τ、垂向紊流黏性系数Vr和垂向紊流扩散系数Vt的改变(降雨期均大于非降雨期)是导致藻类迅速衰亡的关键因素.
(2) 10月10~18日出现降雨现象使得培养装置内Chl-a浓度明显降低, 持续降雨现象会抑制藻类的生长, 但降雨所导致的水动力条件不是藻类衰亡的根本原因; 降雨导致水体垂向扰动增强, 混合层不断扩大, 打破水体原有水温分层从而抑制藻类的生长增殖是水华消失的根本原因.
(3) 由方差分析及相关分析表明:降雨带来降雨量、水温、光照强度、总氮(TN)和溶解性总氮(DTN)改变是影响3种主要藻种比生长速率的关键环境参数.
| [1] |
李欣, 宋林旭, 纪道斌, 等. 春季敏感时期三峡水库典型支流沉积物-水界面氮释放特性[J]. 环境科学, 2018, 39(3): 1113-1121. Li X, Song L X, Ji D B, et al. Characteristics of nitrogen release at the sediment-water interface in the typical tributaries of the Three Gorges Reservoir during the sensitive period in spring[J]. Environmental Science, 2018, 39(3): 1113-1121. |
| [2] | Karp-Boss L, Boss E, Jumars P A. Nutrient fluxes to planktonic osmotrophs in the presence of fluid motion[J]. Oceanography and Marine Biology, 1996, 34: 71-107. |
| [3] | Edwards N, Beeton S, Bull A T, et al. A novel device for the assessment of shear effects on suspended microbial cultures[J]. Applied Microbiology and Biotechnology, 1989, 30(2): 190-195. |
| [4] | Sin Y, Wetzel R L, Anderson I C. Spatial and temporal characteristics of nutrient and phytoplankton dynamics in the York River Estuary, Virginia:analyses of long-term data[J]. Estuaries, 1999, 22(2): 260-275. DOI:10.2307/1352982 |
| [5] |
黄宁秋.水动力条件对三峡库区次级支流典型藻类生长影响[D].重庆: 重庆大学, 2015. Huang N Q. Effects of hydrodynamic condition on the growth of algae at secondary tributary in the Three Gorges Reservoir[D]. Chongqing: Chongqing University, 2015. http://cdmd.cnki.com.cn/Article/CDMD-10611-1015970162.htm |
| [6] | Hondzo M, Lyn D. Quantified small-scale turbulence inhibits the growth of a green alga[J]. Freshwater Biology, 1999, 41(1): 51-61. DOI:10.1046/j.1365-2427.1999.00389.x |
| [7] | Warnaars T A, Hondzo M. Small-scale fluid motion mediates growth and nutrient uptake of Selenastrum capricornutum[J]. Freshwater Biology, 2006, 51(6): 999-1015. DOI:10.1111/fwb.2006.51.issue-6 |
| [8] | Berger S A, Diehl S, Stibor H, et al. Water temperature and mixing depth affect timing and magnitude of events during spring succession of the plankton[J]. Oecologia, 2007, 150(4): 643-654. |
| [9] |
刘心愿, 宋林旭, 纪道斌, 等. 降雨对蓝藻水华消退影响及其机制分析[J]. 环境科学, 2018, 39(2): 774-782. Liu X Y, Song L X, Ji D B, et al. Effect of the rainfall on extinction of cyanobacteria bloom and its mechanism analysis[J]. Environmental Science, 2018, 39(2): 774-782. |
| [10] |
崔玉洁.三峡水库香溪河藻类生长敏感生态动力学过程及其模拟[D].武汉: 武汉大学, 2017. Cui Y J. The sensitive ecological dynamic processes and their simulations of algal growth of Xiangxi Bay in the Three Gorges Reservoir[D]. Wuhan: Wuhan University, 2017. http://cdmd.cnki.com.cn/Article/CDMD-10486-1017189909.htm |
| [11] |
李锦秀, 杜斌, 孙以三. 水动力条件对富营养化影响规律探讨[J]. 水利水电技术, 2005, 36(5): 15-18. Li J X, Du B, Sun Y S. Effect of hydrodynamics on the eutrophication[J]. Water Resources and Hydropower Engineering, 2005, 36(5): 15-18. |
| [12] |
曹巧丽, 黄钰玲, 陈明曦. 水动力条件下蓝藻水华生消的模拟实验研究与探讨[J]. 人民珠江, 2008(4): 8-10, 13. Cao Q L, Huang Y L, Chen M X. Experimental research on hydro-dynamic simulation of cyanobateria bloom outbreak and extinction[J]. Pearl River, 2008(4): 8-10, 13. DOI:10.3969/j.issn.1001-9235.2008.04.003 |
| [13] |
徐耀阳. 浅述湖泊生态系统营养状态二维坐标评价方法[J]. 安徽农学通报, 2011, 17(17): 53-54. Xu Y Y. A brief introduction of a two-dimensional graphical approach for assessing lake ecosystems[J]. Auhui Agricultural Science Bulletin, 2011, 17(17): 53-54. DOI:10.3969/j.issn.1007-7731.2011.17.025 |
| [14] |
郑丙辉, 张佳磊, 王丽婧, 等. 大宁河水华敏感期浮游植物与环境因子关系[J]. 环境科学, 2011, 32(3): 641-648. Zheng B H, Zhang J L, Wang L J, et al. Exploration of relationships between phytoplankton and related environmental factors in the Daning river during sensitive period of algal blooms[J]. Environmental Science, 2011, 32(3): 641-648. |
| [15] |
马骏, 刘德富, 纪道斌, 等. 三峡水库支流库湾低流速条件下测流方法探讨及应用[J]. 长江科学院院报, 2011, 28(6): 30-34, 54. Ma J, Liu D F, Ji D B, et al. Flow measurement methods under low flow velocity at the tributary bays of three gorges reservoir[J]. Journal of Yangtze River Scientific Research Institute, 2011, 28(6): 30-34, 54. DOI:10.3969/j.issn.1001-5485.2011.06.007 |
| [16] |
纪道斌, 李媛, 孔松, 等. 典型暴雨洪水对三峡水库香溪河库湾水华的影响[J]. 中国农村水利水电, 2013(6): 39-44. Ji D B, Li Y, Kong S, et al. The influence of a typical storm flood on the water bloom in the Xiangxi Bay, Three Gorges Reservoir[J]. China Rural Water and Hydropower, 2013(6): 39-44. DOI:10.3969/j.issn.1007-2284.2013.06.011 |
| [17] |
高圻烽, 何国建, 方红卫, 等. 三峡库区支流中水体的垂向掺混对于藻类生长的影响[J]. 水利学报, 2017, 48(1): 96-103. Gao Q F, He G J, Fang H W, et al. Effects of vertical mixing on algal growth in the tributary of Three Gorges Reservoir[J]. Journal of Hydraulic Engineering, 2017, 48(1): 96-103. |
| [18] |
李哲, 谢丹, 郭劲松, 等. 三峡水库澎溪河典型优势藻原位生长速率的初步研究[J]. 湖泊科学, 2012, 24(5): 746-754. Li Z, Xie D, Guo J S, et al. Preliminary study on in situ growth rate of dominant algae species in Pengxi River of the Three Gorges Reservoir[J]. Journal of Lake Sciences, 2012, 24(5): 746-754. DOI:10.3969/j.issn.1003-5427.2012.05.016 |
| [19] |
冯婧, 李哲, 闫彬, 等. 三峡水库不同运行阶段澎溪河典型优势藻原位生长速率[J]. 湖泊科学, 2014, 26(2): 235-242. Feng J, Li Z, Yan B, et al. In situ growth rate of dominant algae species in Pengxi River of the Three Gorges Reservoir[J]. Journal of Lake Sciences, 2014, 26(2): 235-242. |
| [20] |
杨毓, 卢金锁, 张颖. 深水型水库藻形态功能组(MBFG)的季节演替特征[J]. 环境科学, 2017, 38(1): 121-128. Yang Y, Lu J S, Zhang Y. Seasonal succession characteristics of the morphologically-based functional groups (MBFG) in deep-water reservoir[J]. Environmental Science, 2017, 38(1): 121-128. DOI:10.3969/j.issn.1673-2049.2017.01.016 |
| [21] | Butler E I. A manual of chemical and biological methods for sea water analysis:by Timothy Parsons, Yoshiaki Maita and Carol M. Lalli, Pergamon Press, Oxford, 1984, 184 pp., USMYM8.95/19.50, £5.50/£12.25[J]. Deep Sea Research Part A. Oceanographic Research Papers, 1984, 31(12): 1523. DOI:10.1016/0198-0149(84)90086-4 |
| [22] | Littler M M, Littler D S. Handbook of phycological methods[M]. Cambridge: Cambridge University Press, 1985. |
| [23] | 国家环境保护总局. 水和废水监测分析方法[M]. (第四版). 北京: 中国环境科学出版社, 2002. |
| [24] | Schmitt F G. About Boussinesq's turbulent viscosity hypothesis:historical remarks and a direct evaluation of its validity[J]. Comptes Rendus Mécanique, 2007, 335(9-10): 617-627. DOI:10.1016/j.crme.2007.08.004 |
| [25] |
何金平.金山湖水体运行方案及物质输运数值模拟研究[D].南京: 河海大学, 2007. He J P. The operational scheme and numerical simulation of pollutant transportation in Jinshan Lake[D]. Nanjing: Hohai University, 2007. http://cdmd.cnki.com.cn/Article/CDMD-10294-2007167539.htm |
| [26] | Hondzo M M, Kapur A, Lembi C A. The effect of small-scale fluid motion on the green alga Scenedesmus quadricauda[J]. Hydrobiologia, 1997, 364(2-3): 225-235. |
| [27] | 普朗特.流体力学概论[M].郭永怀, 陆士嘉, 译.北京: 科学出版社, 1981. |
| [28] | Wetzel R G. Photosynthesis, productivity and growth: the physiological ecology of phytoplankton: G. P. Harris. Ergebnisse die Limnologie, Archiv für Hydrobiologie, Beiheft 10. E. Schweizerbart'sche Verlagsbuchhandlung, Stuttgart, 1978, 171 pp., ISBN 3-510-47008-7[J]. Aquatic Botany, 1980, 9: 98-99. https://www.researchgate.net/publication/37879648_Photosynthesis_productivity_and_growth_The_physiological_ecology_of_phytoplankton |
| [29] | Delgadillo-Hinojosa F, Gaxiola-Castro G, Segovia-Zavala J A, et al. The effect of vertical mixing on primary production in a bay of the gulf of California[J]. Estuarine, Coastal and Shelf Science, 1997, 45(1): 135-148. DOI:10.1006/ecss.1996.0167 |
| [30] | Oliver R L, Whittington J, Lorenz Z, et al. The influence of vertical mixing on the photoinhibition of variable chlorophyll a fluorescence and its inclusion in a model of phytoplankton photosynthesis[J]. Journal of Plankton Research, 2003, 25(9): 1107-1129. DOI:10.1093/plankt/25.9.1107 |
| [31] |
田泽斌, 刘德富, 姚绪姣, 等. 水温分层对香溪河库湾浮游植物功能群季节演替的影响[J]. 长江流域资源与环境, 2014, 23(5): 700-707. Tian Z B, Liu D F, Yao X J, et al. Effect of water temperature stratification on the seasonal succession of phytoplankton function grouping in Xiangxi Bay[J]. Resources and Environment in the Yangtze Basin, 2014, 23(5): 700-707. |
| [32] | Godlewska M, Mazurkiewicz-Boroń G, Pociecha A, et al. Effects of flood on the functioning of the Dobczyce reservoir ecosystem[J]. Hydrobiologia, 2003, 504(1-3): 305-313. DOI:10.1023/B:HYDR.0000008530.31142.81 |
| [33] |
望雪, 程豹, 杨正健, 等. 澜沧江流域沉积物间隙水-上覆水营养盐特征与交换通量分析[J]. 环境科学, 2018, 39(5): 2126-2134. Wang X, Cheng B, Yang Z J, et al. Differences in diffusive fluxes of nutrients from sediment between the natural river areas and reservoirs in the Lancang River Basin[J]. Environmental Science, 2018, 39(5): 2126-2134. |
| [34] | Falkowski P G. The role of phytoplankton photosynthesis in global biogeochemical cycles[J]. Photosynthesis Research, 1994, 39(3): 235-258. DOI:10.1007/BF00014586 |
| [35] | Lavery P S, Lukatelich R J, McComb A J. Changes in the biomass and species composition of macroalgae in a eutrophic estuary[J]. Estuarine, Coastal and Shelf Science, 1991, 33(1): 1-22. DOI:10.1016/0272-7714(91)90067-L |
| [36] | Domingues R B, Anselmo T P, Barbosa A B, et al. Light as a driver of phytoplankton growth and production in the freshwater tidal zone of a turbid estuary[J]. Estuarine, Coastal and Shelf Science, 2011, 91(4): 526-535. DOI:10.1016/j.ecss.2010.12.008 |
| [37] |
刘青, 张晓芳, 李太武, 等. 光照对4种单胞藻生长速率、叶绿素含量及细胞周期的影响[J]. 大连水产学院学报, 2006, 21(1): 24-30. Liu Q, Zhang X F, Li T W, et al. Effects of light on growth rate, chlorophyll level and cell cycle in four alga species[J]. Journal of Dalian Fisheries University, 2006, 21(1): 24-30. DOI:10.3969/j.issn.1000-9957.2006.01.005 |
| [38] |
蓝天.淀山湖浮游植物与环境因子的相关性研究[D].上海: 上海师范大学, 2014. Lan T. Study on the correlation between phytoplankton and environmental factors in Dianshanhu[D]. Shanghai: Shanghai Normal University, 2014. http://cdmd.cnki.com.cn/Article/CDMD-10270-1014337542.htm |
| [39] |
王利利.水动力条件下藻类生长相关影响因素研究[D].重庆: 重庆大学, 2006. Wang L L. Research on the relevant factors of the algal growth in hydrodynamic condition[D]. Chongqing: Chongqing University, 2006. |
| [40] |
梁英, 刘春强, 田传远, 等. 不同营养盐浓度对6种海洋微藻群落演替的影响[J]. 水产科学, 2013, 32(11): 627-635. Liang Y, Liu C Q, Tian C Y, et al. Effects of nutrient concentrations on community succession in six marine microalga species[J]. Fisheries Science, 2013, 32(11): 627-635. DOI:10.3969/j.issn.1003-1111.2013.11.001 |
| [41] | Clausing R J, Fong P. Environmental variability drives rapid and dramatic changes in nutrient limitation of tropical macroalgae with different ecological strategies[J]. Coral Reefs, 2016, 35(2): 669-680. DOI:10.1007/s00338-016-1403-6 |
| [42] |
韦力元, 王真, 洪松. 气象与水质对滇池草海叶绿素a浓度的影响[J]. 环境科学与技术, 2016, 39(10): 26-30, 35. Wei L Y, Wang Z, Hong S. Influences of meteorological conditions and water quality factors on chlorophyll-a in Caohai, Dianchi Lake[J]. Environmental Science & Technology, 2016, 39(10): 26-30, 35. |