 2019, Vol. 40
2019, Vol. 40



氧化亚氮(N2O)是重要的温室气体之一, 其全球增温潜势是CO2的265倍[1], 且在大气中滞留时间较长. N2O浓度的增加不仅加剧了全球温室效应, 而且参与大气中许多光化学反应, 导致臭氧层的破坏与地面紫外线辐射增强[2].土壤硝化和反硝化微生物作用是N2O产生的主要途径[3]. N2O在农业中的排放量占人类活动产生总量的65%[4].稻田排放的N2O约占中国农田N2O排放总量的7%~11%[5]. N2O是土壤氮循环的中间产物之一, 现有研究已从分子生物学上表明N2O产生的主要微生物学机制, 硝化反硝化作用的一般过程如图 1所示.

|
图 1 土壤N2O排放的微生物学过程[6] Fig. 1 Microbial processes of N2O emissions |
近年来随着集约化畜牧业的发展, 使得大量兽用抗生素随畜禽粪尿进入环境[7], 对土壤中微生物、动物、植物产生直接或间接毒害作用, 并且诱导抗性细菌及抗性基因出现, 从而导致特殊的生态毒理效应, 最终抗生素可以通过饮用水源以及植物吸收进入食物链威胁人类健康[8].抗生素能直接杀死土壤等环境介质中某些微生物或抑制相关微生物的生长, 从而可能影响土壤中硝化、反硝化菌的代谢和繁殖[9, 10].各国的畜禽和畜牧养殖业广泛使用抗生素[11], 有研究表明, 约30%~90%的抗生素不能被机体代谢而释放到环境中[12].由于抗生素的长期不断输入和土壤的吸附作用, 导致抗生素在土壤中持久存在[13].抗生素母体化合物及其代谢产物进入并残留在土壤环境中, 对土壤环境造成不同程度的污染[14].
磺胺类抗生素是一类在农业生产中应用最为广泛的化学合成抗菌药.在目前常用的几类兽用抗生素中, 磺胺类抗生素具有较大的生态毒性和环境持久性[15, 16].我国学者研究发现, 畜禽粪便中磺胺类药物的残留达到mg·kg-1水平[17, 18], 甚至有报道表明, 上海施用过猪粪的农田土壤磺胺类抗生素含量范围高达5.85~33.37 mg·kg-1[19]. Huang等[20]的研究发现磺胺类药物在活性污泥系统中对硝化过程的抑制作用与其取代基的结构特征有关. Ahmad等[21]的研究发现金霉素和磺胺二甲嘧啶对地下水中的反硝化过程均有不同程度的抑制效果.进入土壤中的抗生素可影响土壤微生物的存活力、群落结构、代谢功能、种群数量等, 从而干扰食物链, 使生物多样性水平降低, 生物量和生物区系发生改变, 甚至影响土壤生物学和养分循环过程及自净能力, 最终影响土壤肥力和土壤养分平衡[22].农田土壤N2O的产生是一个微生物参与的过程, 任何影响到微生物活动的条件均能够影响土壤N2O的产生和排放.因此, 兽用抗生素磺胺二甲嘧啶在稻田土壤中的环境行为可能会对土壤硝化、反硝化微生物产生作用, 进而影响稻田土壤N2O的产生和排放.
近年来, 国内外在稻田土壤N2O排放方面开展了大量研究, 但是大多数研究都集中在土壤理化性质对硝化、反硝化的影响[23], 有关抗生素的研究多集中在抗生素在土壤中降解、迁移等环境行为及对土壤微生物的影响[24], 关于随有机粪源大量进入农田的抗生素对土壤硝化反硝化过程产生的N2O排放的影响研究鲜见报道.本文以磺胺类抗生素中的典型代表磺胺二甲嘧啶为研究对象, 通过稻田原位观测试验, 以未经抗生素污染的农家猪粪为基肥, 分别添加不同浓度的SMZ, 探究在不同浓度磺胺二甲嘧啶胁迫条件下稻田N2O排放和硝化、反硝化功能基因的变化, 以期为了解磺胺二甲嘧啶对N2O排放的影响及机制提供科学依据.
1 材料与方法 1.1 试验区概况试验田位于江苏省南京市江宁区淳化街道(118°59′E, 31°57′N), 试验时间为2017年6~11月.年平均气温22.68℃, 水稻生长季总降雨量为890.6 mm.土壤充水孔隙度(WFPS)和气温变化如图 2所示.试验田土壤有机碳含量为29.39 g·kg-1, 全氮为1.93 g·kg-1, pH值为7.67, 土壤容重为1.30 g·cm-3.

|
图 2 2017年水稻生长季日均气温和土壤充水孔隙度 Fig. 2 Variation of the average daily temperature and soil water-filled-pore-space during the rice season in 2017 |
本试验供试水稻品种为南粳55号.供试抗生素为上海麦克林公司生产的磺胺二甲嘧啶(SMZ)试剂.供试猪粪为经堆置的从未接触过任何抗生素的家猪排泄物, 全氮含量为8.82 g·kg-1, 有机质为752.03 g·kg-1.试验共设5个处理, 分别为:不施肥不添加抗生素(CK); 以猪粪为基肥, 尿素为追肥, 分别添加0、5、15和30 mg·kg-1的磺胺二甲嘧啶(SMZ0、SMZ5、SMZ15和SMZ30). SMZ随基肥一次性施入, 追肥时不再添加.
试验田采用微区设计, 微区面积为3 m×2 m, 每个处理设置随机排列的3个重复.各小区之间设有用塑料薄膜覆盖的80 cm宽、30 cm高的田埂, 各小区有各自独立的灌排水系统, 以防止水肥串流.
水稻于2017年6月4日播种, 2017年11月11日收获.除CK外, 其它处理施氮总量(以N计)均为220 kg·hm-2, 按基肥:追肥=1:1比例分别于2017年6月4日和2017年7月29日撒施.整个水稻生育期N:P2O5:K2O施肥比例为2:1:1.除肥料外, 试验田按照当地常规措施进行水药管理.水稻生长季前期为持续淹水状态, 7月25~27日为烤田期, 烤田之后为干湿交替状态.
1.3 样品采集及测定方法 1.3.1 农田系统N2O的采集与测定方法采用静态箱-气相色谱法进行采集与测定稻田N2O的排放通量[25].静态暗箱由不锈钢制成, 箱体高度随水稻高度而增加, 箱体顶部及四周密闭并包有海绵和锡箔纸以起到反射阳光和保温的作用, 各微区埋有不锈钢底座, 底座上部有3 cm深凹槽.采样时将采样箱罩在凹槽上, 并向凹槽内注水以保持采样箱密闭.箱内顶部装有微型风扇, 在采样时启动, 以便于箱内气体一直保持混匀状态.施肥后次日开始采样, 采样频率为一周两次, 采样时间为上午08:00~12:00, 施加基肥与追肥后第一周加密采样一次, 2017年9月18日之后每周采样一次.每个采样点于箱子密封后的0、10和20 min时用注射器收集150 mL气体, 储存于铝箔气袋中, 并记录箱内温度.使用气相色谱仪(Agilent 7890B)检测与分析气体样品. N2O测定所用载气为99.999%高纯氮; 检测器为电子捕获检测器(ECD), 检测温度为300℃, 柱温为50℃.通过对每组3个样品N2O浓度与相对应的采样间隔时间(0、10和20 min)进行线性回归, 可求得该采样点的N2O排放通量.根据气温、大气压力、普适气体常数、气体分子量、采样箱有效高度等, 求得单位面积的N2O排放量, 具体计算公式如下:
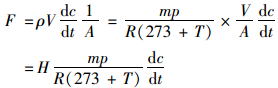
|
式中, F为气体排放通量, μg·(m2·h)-1; ρ为气体密度, g·L-1; R为普适气体常数, 8.314 J·(mol·K)-1; A为采样箱横截面积, m2; H为采样箱高度, m; V为箱内空气体积, V=AH, m3; p为采样点大气压力, 通常视为标准大气压1.013×105 Pa; m为气体摩尔质量, g·mol-1; T为采样箱平均气温, ℃; dc/dt为单位时间内温室气体浓度变化率, nL·(L·h)-1.
通过3个重复的观测值按时间加权平均后获得N2O的季节排放总量, 计算公式如下:
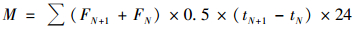
|
式中, M为气体累积排放量, μg·m-2; N为采样次数; t为采样时间即距初次采样的天数, d; F为气体排放通量, μg·(m2·h)-1.
1.3.2 土壤样品的采集与测定方法施肥后每周采集土样一次.各小区采用梅花形布点法用土钻采集0~10 cm土壤样品, 制成混合样, 取好的土壤样品分别装入自封袋, 标记密封带回实验室后迅速过2 mm筛并置于-20℃冰箱保存, 用于测定土壤NH4+-N、NO3--N和基因丰度指标.结合N2O测定结果及施肥日期, 选取6月9日、6月22日、7月6日、7月21日、8月4日、8月24日及收获日11月11日测定相关土壤微生物功能基因丰度.每次气体采样同时, 用MP-406Ⅲ型土壤水分温度测定仪(南通中天精密仪器有限公司)测定5 cm深度的土壤温度和土壤水分(体积比), 根据土壤容重将体积水分换算成土壤空隙含水量(WFPS); 土壤NH4+-N、NO3--N测定方法为:将土壤用2 mol·L-1的氯化钾溶液浸提, 振荡过滤后用连续流动分析仪(型号:AA3, SEAL Analytical, 产地:英国)进行测定.土壤样品全基因组的提取采用PowerSoil® DNA Isolation kit强力土壤DNA提取试剂盒法.所提取的土壤总DNA的浓度用NanoDrop2000超微量紫外分光光度计测定.所有样品的DNA浓度调节为10 ng·μL-1后, 采用实时荧光定量PCR技术测定各基因丰度.采用统一程序扩增所有的基因, PCR扩增采用20 μL反应体系:6.8 μL ddH2O; 0.4 μL Forward Primer(10 μmol·L-1), 0.4 μL Reverse Primer(10 μmol·L-1), 即正反向引物各0.4 μL; 10 μL扩增酶混合物SYBR Premix Ex TaqTM(2×); 0.4 μL ROX Reference Dye(50×); 2 μL DNA模板.扩增程序为: 95℃ 30 s; 95℃ 5 s, 55℃ 30 s, 72℃ 1 min, 40个循环; 95℃ 15 s, 60℃ 1 min, 95℃ 15 s.引物及相应扩增程序见表 1.
|
|
表 1 目的基因的引物序列 Table 1 Primer set for the target genes |
1.3.3 土壤中磺胺二甲嘧啶残留量的测定方法[32]
(1) 前处理 采集的新鲜土样经冷冻干燥机干燥后, 准确称取1 g土样, 置于50 mL离心管中, 加入V(甲醇):V(EDTA-Mcllvaine缓冲液)=1:1混合液5 mL, 振荡, 超声提取各10 min, 离心并收集上清液, 按上述方法反复提取3次, 合并提取液, 浓缩至5 mL. SPC固相萃取柱(SAX柱-HLB柱串联)预先经过6mL甲醇、6mL超纯水进行淋洗.开启真空泵控制流速在3~5 mL·min-1, 将浓缩液依次通过SPC固相萃取柱, 萃取富集后, 用6 mL超纯水对固相萃取柱进行淋洗, 继续抽真空, 除去柱中多余水分, 拆下SAX柱, 继续干燥HLB柱, 然后用3 mL甲醇洗脱HLB柱, 收集洗脱液并浓缩至近干, 最后用2 mL甲醇定容, 待测.
(2) 色谱条件 固定相为:ODS-反相C18柱; 流动相为:乙腈(A):0.1%甲酸溶液(B)=17:83;检测波长270 nm; 检测器为紫外检测器; 柱温25℃; 流速0.8 mL·min-1; 进样量20 μL.
1.4 数据处理本文采用SPSS 19.0和Microsoft 2010软件进行数据计算以及统计分析, 采用OriginPro 9.1软件对数据进行作图.
2 结果与分析 2.1 SMZ对稻田N2O排放通量、累积排放量及土壤NH4+-N、NO3--N的影响整个水稻观测期各处理N2O排放通量如图 3所示, 各处理动态变化趋势基本一致, 不同SMZ处理均未改变稻田N2O的季节性排放规律, 水肥交互状况决定了N2O排放通量峰值的出现(图 2和图 3).将整个水稻生长季分为基肥阶段(6月4日~7月28日)和追肥阶段(7月29日~11月11日).各处理均在基肥期的第5 d(6月9日)达到一个峰值, 其中以SMZ15处理的N2O排放通量峰值最大, 为202.03 μg·(m2·h)-1.追肥后, 各处理在第9 d(8月7日)达到峰值时的N2O排放通量大小为:SMZ30>SMZ15>SMZ0>SMZ5, 其中以SMZ30排放通量最大, 为726.58 μg·(m2·h)-1.各处理的N2O排放通量均在达到峰值后迅速下降, 最终达到相对较低的稳定水平.

|
CK代表未添加猪粪及磺胺二甲嘧啶处理; SMZ0、SMZ5、SMZ15和SMZ30分别代表添加猪粪和0、5、15和30 mg·kg-1的磺胺二甲嘧啶, 下同 图 3 2017年水稻季不同浓度SMZ处理N2O季节排放动态变化 Fig. 3 Seasonal variation in N2O emissions from different SMZ treatments during the rice season in 2017 |
以各处理和每次独立观测时间对N2O排放通量作二因子方差分析表明, 基肥和追肥阶段各处理之间的N2O排放通量无显著差异(P>0.05), 而每次独立观测时间之间的N2O排放通量有极显著差异(P < 0.01).整个观测期, SMZ15与SMZ0之间具有显著差异(P < 0.05), SMZ30、SMZ5与SMZ0均无显著差异(P>0.05).
不同观测阶段N2O累积排放量如表 2所示.结果表明, 与CK相比, 施加高有机质猪粪的SMZ0明显降低了N2O的排放(P < 0.1), 而SMZ的存在改变了此规律, 中、高浓度SMZ处理与CK相比增加了N2O的累积排放, 尽管在统计学上未达显著性差异(P>0.1).均值水平上整个水稻季SMZ15和SMZ30处理的N2O累积排放量分别是SMZ0处理的3.47和4.67倍.
|
|
表 2 水稻季不同施肥处理N2O-N累积排放量1) Table 2 Cumulative emissions of N2O-N from different fertilization treatments during the rice season |
图 4为水稻季各处理的土壤NH4+-N和NO3--N含量动态变化, 整个观测期内各处理的土壤NH4+-N和NO3--N均在施加追肥后出现最大峰值.土壤NH4+-N在整体水平上明显高于NO3--N含量.以各处理和观测时间对土壤NH4+-N和NO3--N含量作二因子方差分析, 结果如下:在基肥期, 与SMZ0处理相比, SMZ30处理显著增加了土壤NH4+-N和NO3--N含量(P < 0.05), 其与SMZ0处理NH4+-N和NO3--N含量比值的平均值分别为1.56和1.18; SMZ15、SMZ5与SMZ0处理之间的NH4+-N和NO3--N含量均无显著差异(P>0.05).追肥期, 与SMZ0处理相比, 除SMZ30显著增加了土壤NO3--N含量(P < 0.05), SMZ15极显著减少了土壤NH4+-N含量(P < 0.01)外, 其它处理与SMZ0之间的NH4+-N和NO3--N含量均无显著差异(P>0.05).

|
图 4 2017年水稻季土壤NH4+-N和NO3--N含量动态变化 Fig. 4 Dynamic variation of soil NH4+-N and NO3--N contents during the rice season in 2017 |
图 5为各处理土壤总细菌16S rRNA基因丰度动态变化.整个观测期内, 各处理土壤总细菌16S rRNA基因丰度变化趋势整体一致, 在不同测定期各处理间差异性不同.除SMZ30处理在7月21日达到最大值外, 其余处理均在6月22日达到峰值.施加追肥后的第6 d(8月4日), SMZ30极显著增加了土壤总细菌16S rRNA基因丰度(P < 0.01), SMZ15处理也起促进作用但未达到显著性差异(P>0.05).

|
相同字母表示该两者间差异性不显著(P>0.05), 不同字母表示差异性达到显著水平(P < 0.05), 下同 图 5 2017年水稻季土壤总细菌16S rRNA基因丰度动态变化 Fig. 5 Dynamic variation of the total soil bacterial 16S rRNA gene abundance during the rice season in 2017 |
经二因子方差分析可得, 整个观测期内, SMZ30、SMZ15、SMZ5与SMZ0处理之间16S rRNA基因丰度有极显著差异(P<0.01), 且SMZ30、SMZ15、SMZ5与SMZ0处理的16S rRNA基因丰度拷贝数比值的平均值分别为:1.58、1.24和0.80, 可知SMZ30和SMZ15处理对土壤总细菌16S rRNA有激活作用; SMZ5对土壤总细菌16S rRNA有轻微抑制作用.
图 6为各处理的土壤AOA amoA和AOB amoA基因丰度动态变化.不同处理的土壤AOA amoA和AOB amoA在整个观测期内变化幅度较大.各处理AOA amoA和AOB amoA基因丰度在6月22日达到峰值, 之后整体呈现下降趋势并最终趋于稳定.在SMZ施入后的第5 d(6月9日), SMZ30和SMZ15处理极显著增加了土壤AOA amoA基因丰度(P < 0.01).施加追肥后的第6 d(8月4日), SMZ30极显著增加了土壤AOA amoA和AOB amoA基因丰度(P < 0.01).

|
图 6 2017年水稻季土壤AOA amoA和AOB amoA基因丰度动态变化 Fig. 6 Dynamic variation of the AOA amoA and AOB amoA genes abundance during the rice season in 2017 |
经二因子方差分析可得, 整个观测期内, SMZ30与SMZ0处理之间AOA amoA和AOB amoA基因丰度有极显著差异(P<0.01), SMZ15与SMZ0处理之间AOA amoA基因丰度有显著差异(P<0.05), 且SMZ30、SMZ15、SMZ5与SMZ0处理的AOA amoA基因丰度拷贝数比值的平均值分别为:1.77、1.37和0.99; AOB amoA基因丰度拷贝数比值的平均值分别为:2.15、1.08和0.92, 可知SMZ30和SMZ15处理对AOA amoA和AOB amoA基因均有激活作用.
2.2.2 SMZ对土壤nirK、nirS和nosZ基因丰度的影响图 7为土壤nirK、nirS和nosZ基因丰度的动态变化.各处理nirK基因丰度在施加SMZ后的第5 d(6月9日)达到峰值, 且在此天和施加追肥后的第6 d(8月4日)SMZ30和SMZ15处理均极显著促进了nirK基因丰度(P < 0.01). 各处理的nirS基因丰度前期逐渐降低, 中后期趋于稳定, 8月24日后各处理均升高; 在施加追肥后的第6 d(8月4日), SMZ30处理显著促进了nirS基因丰度(P < 0.05).各处理的nosZ基因丰度在6月22日达到峰值, 在施加SMZ后的第5 d(6月9日), SMZ30处理显著促进了nosZ基因丰度(P < 0.05);施加追肥后的第6 d(8月4日), SMZ30与SMZ15处理显著增加了nosZ丰度(P < 0.05).

|
图 7 2017年水稻季土壤nirK、nirS和nosZ基因丰度动态变化 Fig. 7 Dynamic variation of the soil nirK, nirS, and nosZ genes abundance during the rice season in 2017 |
经二因子方差分析可得, 整个观测期内, SMZ30与SMZ0处理之间nirK、nirS和nosZ丰度均有显著差异(P<0.05), SMZ15与SMZ0处理之间nosZ丰度有极显著差异(P<0.01), SMZ5与SMZ0处理之间nirK、nirS和nosZ丰度均有极显著差异(P<0.01), 且SMZ30、SMZ15、SMZ5与SMZ0处理的nirK基因丰度拷贝数比值的平均值分别为:1.38、1.65和0.76; nirS基因丰度拷贝数比值的平均值分别为:1.33、1.11和0.76, nosZ基因丰度拷贝数比值的平均值分别为:1.42、1.64和0.77.可知, 整个观测期内SMZ30和SMZ15处理对nirK、nirS和nosZ基因丰度均有激活作用; SMZ5对nirK、nirS和nosZ基因丰度均有一定的抑制作用.
因激活nirK和nirS有利于N2O的产生, 而激活nosZ则反之, 故进一步计算整个水稻生长季不同浓度SMZ处理条件下SMZ30/SMZ0, SMZ15/SMZ0和SMZ5/SMZ0的nirK/nosZ与nirS/nosZ比值的商, 用于判定SMZ影响N2O在反硝化作用中产生(nirK, nirS)和消耗(nosZ)的功能基因丰度在一定程度上的相对强弱. SMZ30/SMZ0, SMZ15/SMZ0和SMZ5/SMZ0的nirK/nosZ比值的商分别为1.02, 1.21和1.01; nirS/nosZ比值的商分别为1.00, 0.84和1.03, 说明除SMZ15的nirS/nosZ小于1外, 其余处理SMZ对nirK和nirS的作用较nosZ更强.
2.3 稻田N2O排放通量与土壤总细菌16S rRNA及硝化反硝化相关功能基因丰度和土壤NH4+-N、NO3--N相关性将各处理的N2O排放通量与各功能基因丰度进行相关性分析, 如表 3所示, N2O排放通量与nirK基因丰度呈极显著正相关(P < 0.01), 与其他基因则无显著相关性(P>0.05), 表明本研究中含nirK的反硝化菌直接影响稻田N2O排放.
|
|
表 3 水稻季N2O排放与相关功能基因及土壤NH4+-N和NO3--N的相关性1) Table 3 Correlation between N2O emission and related functional genes and soil NH4+-N and NO3--N during the rice season |
3 讨论
磺胺二甲嘧啶水溶性较强, 土壤表面对SMZ的吸附量较小, 吸附率较低, 进入土壤环境后容易迁移和渗漏. Pan等[33]研究了5种抗生素在土壤中的吸附性能, 结果表明磺胺二甲嘧啶的移动性最强, 在施肥过程中容易向下迁移.许静等[34]的研究表明磺胺类抗生素在土壤中滞留期长, 完全消解需要较长时间, 且土壤中粪便的加入对磺胺类药物的降解性产生了较大影响, 土壤加入粪便后, 其有机质含量增大, 促进了降解微生物的生长, 进而加快了磺胺类药物在混合基质中的降解速率, 同时其结果证明了微生物降解是磺胺类药物土壤降解的主导途径.本研究中, 7月6日即施入SMZ的第32 d, SMZ5、SMZ15和SMZ30处理土壤中的浓度分别为0.07、0.28和1.36 mg·kg-1, 母体去除率分别高达98.5%、98.2%和95.5%; 8月4日即SMZ施入的第61 d其土壤中的浓度分别为0.05、0.23和0.54 mg·kg-1, 此阶段去除率分别达99.0%、98.4%和98.2%;水稻收获时即SMZ施入的第160 d土壤中的浓度分别为0.03、0.08和0.18 mg·kg-1, 母体土壤残留率分别为0.5%、0.5%和0.6%, 不同浓度SMZ处理母体残留率接近一致.可见SMZ在施入水稻土壤后母体去除较快, 尤其前期最快, 可能与其水溶性强, 迁移快、土壤淹水厌氧降解速率快有关.磺胺二甲嘧啶在土壤中虽然母体残留率并不高, 但其降解产物有可能持久存在, 有研究鉴定出磺胺二甲基嘧啶降解初级产物为2-氨基-4, 6-二甲基嘧啶和4-氨基-羟基苯磺酰胺, 前者永久存在于培养基中, 后者可以进一步降解, 最终分解为二氧化碳和水[35]. Shan等[36]的培养试验也直接证实, 较低浓度磺胺二甲嘧啶通过抑制硝酸盐还原菌降低硝酸盐的分解过程, 随着抗生素浓度增加, nirS/nosZ比值显著增大, 对N2O的产生有刺激作用.结合本研究中高浓度SMZ处理在7月6日和8月4日均增加了土壤16S rRNA基因丰度结果(图 5), 说明磺胺二甲嘧啶母体及其初级降解产物都可能通过改变细菌活性和群落功能多样性进而对土壤硝化反硝化过程产生作用, 并且这种影响可能较为持久.关于SMZ母体及其中间代谢产物的环境生态风险还需进一步研究.
本研究中, 中、高浓度SMZ在整个水稻生长季均表现为对N2O排放有促进作用, 且对其产生的相关微生物均有激活效应, 而低浓度的SMZ作用效果不明显. Hou等[37]的结果表明μg·L-1级低剂量抗生素水平下的磺胺甲嘧啶抑制了河口和海岸沉积物中的反硝化速率, 却促进了N2O的排放, 与此结果存有不一致性, 可能由于有机肥的施入使低浓度SMZ被土壤吸附, 使其作用效果不显著, 鲍陈燕[38]的研究表明, 猪粪的添加可增加土壤对抗生素的吸附强度, 降低土壤中抗生素活性. Hao等[39]的研究表明, 在饲料中添加44 mg·kg-1的磺胺嘧啶和四环素可提高牛粪堆肥中N2O排放量, 与本研究中高浓度SMZ对N2O促排效果基本一致.究其原因, 可能在于SMZ对于N2O产生过程的相关微生物活性有激活效应[40], 土壤硝化作用在高浓度抗生素残留中一直呈现“激活”的规律, 这种激活效应可能因为磺胺类药物残留会刺激硝化细菌的生长, 使其菌落群体活性增强导致.较高浓度(50 mg·kg-1)磺胺类药物对土壤反硝化细菌数量的影响也一直呈现“激活”的规律, 这种激活作用可能是因为反硝化细菌对该药物表现出了免疫作用或是该药物进入到土壤后, 发生了一定的物理化学变化, 从而为反硝化细菌提供了碳源, 促进其生长[41]. Awad等[42]的研究发现, 家禽粪便与20 mg·kg-1的SMZ混施通过改变微生物群落组成, 土壤硝化速率在短期内下降, 而牛建平等[43]的研究认为较高的土壤水分含量可加快SMZ残留的降解速率.
本研究中稻田长期淹水, N2O主要源于反硝化过程.且发现N2O排放通量与nirK基因丰度呈极显著正相关(P < 0.01).作物类型、肥料类型及施用量、土地利用方式、温度、土壤含水量、pH值等均会影响Nir酶的变化, 且nirK基因比nirS基因对环境因子的响应更敏感[44, 45], 含nirK基因的反硝化细菌常被用来表征和预测N2O的释放[46, 47].一般认为, 有机质作为微生物的重要碳源, 是控制反硝化速率的重要因子之一, 本试验中肥源中有机质为752.03 g·kg-1, 含量相对较高, 曾希柏等[48]的研究表明, 有机肥对nirK型反硝化细菌的影响显著. Kleineidam等[49]的研究表明, 磺胺嘧啶与猪粪混施可增加土壤中nirK型反硝化菌含量. Kotzerke等[50]的研究表明, 磺胺嘧啶可提高蚯蚓肠道中nirK型反硝化细菌的活性. Li等[51]的研究结果也表明, N2O排放通量和nirK丰度呈显著正相关, 而与nosZ丰度呈负相关[52], 本研究与其基本一致.
反硝化作用是由相关功能基因和环境因素共同调节的复杂生态过程, 具有不同功能基因型的反硝化细菌群落对环境因素的响应程度存在较大差异.并且, 目前农业生产养殖所用抗生素类型较多, 化学结构差异较大, 抗生素单一及复合污染对反硝化过程的生物学效应可能不尽相同, 有待于进一步研究.
4 结论不同浓度SMZ均未改变稻田N2O的季节性排放规律, 但中、高浓度增加了N2O排放, 也增加了土壤NO3--N含量, 且对土壤总细菌16S rRNA、AOA amoA、AOB amoA、nirK、nirS和nosZ基因丰度均有显著激活作用, 且SMZ对nirK和nirS基因的作用效果较nosZ更强, 低浓度SMZ作用则不明显.此外, N2O排放通量与nirK基因丰度呈极显著正相关, 说明SMZ主要通过影响稻田土壤反硝化微生物活性进而影响N2O的排放.
| [1] | IPCC. Climate Change 2013: The physical science basis. Contribution of working group I to the fifth assessment report of the intergovernmental panel on climate change[R]. Cambridge: Cambridge University Press, 2013. |
| [2] | IPCC. Climate Change 2007: The physical science basis. Contribution of working group I to the fourth assessment report of the intergovernmental panel on climate change[R]. Cambridge: Cambridge University Press, 2007. |
| [3] |
李平, 郎漫. 硝化和反硝化过程对林地和草地土壤N2 O排放的贡献[J]. 中国农业科学, 2013, 46(22): 4726-4732. Li P, Lang M. Contribution of nitrification and denitrification to the nitrous oxide emission from forest and grassland soils[J]. Scientia Agricultura Sinica, 2013, 46(22): 4726-4732. DOI:10.3864/j.issn.0578-1752.2013.22.010 |
| [4] | Nan W G, Yue S C, Li S Q, et al. Characteristics of N2 O production and transport within soil profiles subjected to different nitrogen application rates in China[J]. Science of the Total Environment, 2016, 542: 864-875. DOI:10.1016/j.scitotenv.2015.10.147 |
| [5] | Zou J W, Huang Y, Zheng X H, et al. Quantifying direct N2 O emissions in paddy fields during rice growing season in mainland China:dependence on water regime[J]. Atmospheric Environment, 2007, 41(37): 8030-8042. DOI:10.1016/j.atmosenv.2007.06.049 |
| [6] |
朱永官, 王晓辉, 杨小茹, 等. 农田土壤N2 O产生的关键微生物过程及减排措施[J]. 环境科学, 2014, 35(2): 792-800. Zhu Y G, Wang X H, Yang X R, et al. Key microbial processes in nitrous oxide emissions of agricultural soil and mitigation strategies[J]. Environmental Science, 2014, 35(2): 792-800. |
| [7] |
金彩霞, 司晓薇, 王万峰, 等. 不同形态磺胺类药物在根-土界面的空间分布及毒性评价[J]. 环境科学, 2017, 38(4): 1683-1688. Jin C X, Si X W, Wang W F, et al. Forms distribution and ecotoxicity of three forms of sulfonamides in root-soil interface of maize[J]. Environmental Science, 2017, 38(4): 1683-1688. |
| [8] |
赵方凯, 杨磊, 乔敏, 等. 土壤中抗生素的环境行为及分布特征研究进展[J]. 土壤, 2017, 49(3): 428-436. Zhao F K, Yang L, Qiao M, et al. Environmental behavior and distribution of antibiotics in soils:a review[J]. Soils, 2017, 49(3): 428-436. |
| [9] |
刘款, 孙明明, 刘满强, 等. 土壤反硝化对磺胺嘧啶及抗性基因消减的影响[J]. 土壤, 2017, 49(3): 482-491. Liu K, Sun M M, Liu M Q, et al. Effects of anaerobic denitrification on the dissipation of sulfadiazine and resistance genes in soil[J]. Soils, 2017, 49(3): 482-491. |
| [10] |
杨腾飞, 张小寒, 黄铄淇, 等. 磺胺甲恶唑和甲氧苄啶对生物脱氮过程的影响规律[J]. 环境化学, 2018, 37(3): 471-479. Yang T F, Zhang X H, Huang S Q, et al. Effect of sulfamethoxazole and trimethoprim on biological nitrogen removal[J]. Environmental Chemistry, 2018, 37(3): 471-479. |
| [11] |
王娜, 王昝畅, 郭欣妍, 等. 兽药抗生素环境风险控制管理政策探析[J]. 生态与农村环境学报, 2017, 33(7): 586-591. Wang N, Wang Z C, Guo X Y, et al. Management policies for control of environmental risks of veterinary antibiotics[J]. Journal of Ecology and Rural Environment, 2017, 33(7): 586-591. |
| [12] | Fang H, Han Y L, Yin Y M, et al. Variations in dissipation rate, microbial function and antibiotic resistance due to repeated introductions of manure containing sulfadiazine and chlortetracycline to soil[J]. Chemosphere, 2014, 96: 51-56. DOI:10.1016/j.chemosphere.2013.07.016 |
| [13] | Tasho R P, Cho J Y. Veterinary antibiotics in animal waste, its distribution in soil and uptake by plants:a review[J]. Science of the Total Environment, 2016, 563-564: 366-376. DOI:10.1016/j.scitotenv.2016.04.140 |
| [14] | Everts S. Drugs in the environment:Stakeholders consider ways to reduce the IMPACT of DRUGS that get past sewage treatment plants and into nature[J]. Chemical & Engineering News, 2010, 88(13): 23-24. |
| [15] | Ma J W, Lin H, Sun W C, et al. Soil microbial systems respond differentially to tetracycline, sulfamonomethoxine, and ciprofloxacin entering soil under pot experimental conditions alone and in combination[J]. Environmental Science and Pollution Research, 2014, 21(12): 7436-7448. DOI:10.1007/s11356-014-2685-2 |
| [16] | Ding G C, Radl V, Schloter-Hai B, et al. Dynamics of soil bacterial communities in response to repeated application of manure containing sulfadiazine[J]. PLoS One, 2014, 9(3): e92958. DOI:10.1371/journal.pone.0092958 |
| [17] |
邰义萍, 罗晓栋, 莫测辉, 等. 广东省畜牧粪便中喹诺酮类和磺胺类抗生素的含量与分布特征研究[J]. 环境科学, 2011, 32(4): 1188-1193. Tai Y P, Luo X D, Mo C H, et al. Occurrence of quinolone and sulfonamide antibiotics in swine and cattle manures from large-scale feeding operations of Guangdong province[J]. Environmental Science, 2011, 32(4): 1188-1193. |
| [18] |
刘锋, 廖德润, 李可, 等. 畜禽养殖基地磺胺类喹诺酮类和大环内酯类抗生素污染特征[J]. 农业环境科学学报, 2013, 32(4): 847-853. Liu F, Liao D R, Li K, et al. Pollution characteristics of the sulfonamides, quinolones and macrolides in the samples collected from livestock and poultry feedlots[J]. Journal of Agro-Environment Science, 2013, 32(4): 847-853. |
| [19] | Ji X L, Shen Q H, Liu F, et al. Antibiotic resistance gene abundances associated with antibiotics and heavy metals in animal manures and agricultural soils adjacent to feedlots in Shanghai; China[J]. Journal of Hazardous Materials, 2012, 235-236: 178-185. DOI:10.1016/j.jhazmat.2012.07.040 |
| [20] | Huang X F, Feng Y, Hu C, et al. Mechanistic model for interpreting the toxic effects of sulfonamides on nitrification[J]. Journal of Hazardous Materials, 2016, 305: 123-129. DOI:10.1016/j.jhazmat.2015.11.037 |
| [21] | Ahmad M, Vithanage M, Kim K, et al. Inhibitory effect of veterinary antibiotics on denitrification in groundwater:a microcosm approach[J]. The Scientific World Journal, 2014, 2014: 879831. |
| [22] | Vaclavik E, Halling-Sørensen B, Ingerslev F. Evaluation of manometric respiration tests to assess the effects of veterinary antibiotics in soil[J]. Chemosphere, 2004, 56(7): 667-676. DOI:10.1016/j.chemosphere.2004.02.018 |
| [23] | Huang S H, Pant H K, Lu J. Effects of water regimes on nitrous oxide emission from soils[J]. Ecological Engineering, 2007, 31(1): 9-15. DOI:10.1016/j.ecoleng.2007.04.001 |
| [24] |
贺纪正, 张丽梅. 土壤氮素转化的关键微生物过程及机制[J]. 微生物学通报, 2013, 40(1): 98-108. He J Z, Zhang L M. Key processes and microbial mechanisms of soil nitrogen transformation[J]. Microbiology China, 2013, 40(1): 98-108. |
| [25] | Wang Y S, Wang Y H. Quick measurement of CH4, CO2 and N2 O emissions from a short-plant ecosystem[J]. Advances in Atmospheric Sciences, 2003, 20(5): 842-844. DOI:10.1007/BF02915410 |
| [26] | Ovreås L, Forney L, Daae F L, et al. Distribution of bacterioplankton in meromictic lake saelenvannet, as determined by denaturing gradient gel electrophoresis of PCR-amplified gene fragments coding for 16S rRNA[J]. Applied and Environmental Microbiology, 1997, 63(9): 3367-3373. |
| [27] | Tourna M, Freitag T E, Nicol G W, et al. Growth, activity and temperature responses of ammonia-oxidizing archaea and bacteria in soil microcosms[J]. Environmental Microbiology, 2008, 10(5): 1357-1364. DOI:10.1111/j.1462-2920.2007.01563.x |
| [28] | Stephen J R, Chang Y J, Macnaughton S J, et al. Effect of toxic metals on indigenous soil β-subgroup proteobacterium ammonia oxidizer community structure and protection against toxicity by inoculated metal-resistant bacteria[J]. Applied and Environmental Microbiology, 1999, 65(1): 95-101. |
| [29] | Henry S, Baudoin E, López-Gutiérrez J C, et al. Quantification of denitrifying bacteria in soils by nirK gene targeted real-time PCR[J]. Journal of Microbiological Methods, 2004, 59(3): 327-335. DOI:10.1016/j.mimet.2004.07.002 |
| [30] | Braker G, Fesefeldt A, Witzel K P. Development of PCR primer systems for amplification of nitrite reductase genes (nirK and nirS) to detect denitrifying bacteria in environmental samples[J]. Applied and Environmental Microbiology, 1998, 64(10): 3769-3775. |
| [31] | Scala D J, Kerkhof L J. Nitrous oxide reductase (nosZ) gene-specific PCR primers for detection of denitrifiers and three nosZ genes from marine sediments[J]. FEMS Microbiology Letters, 1998, 162(1): 61-68. DOI:10.1111/fml.1998.162.issue-1 |
| [32] |
李彦文, 莫测辉, 赵娜, 等. 高效液相色谱法测定水和土壤中磺胺类抗生素[J]. 分析化学, 2008, 36(7): 954-958. Li Y W, Mo C H, Zhao N, et al. Determination of sulfonamides antibiotics in water and soil using high performance liquid chromatography[J]. Chinese Journal of Analytical Chemistry, 2008, 36(7): 954-958. DOI:10.3321/j.issn:0253-3820.2008.07.017 |
| [33] | Pan M, Chu L M. Adsorption and degradation of five selected antibiotics in agricultural soil[J]. Science of the Total Environment, 2016, 545-546: 48-56. DOI:10.1016/j.scitotenv.2015.12.040 |
| [34] |
许静, 王娜, 孔德洋, 等. 有机肥源磺胺类抗生素在土壤中的降解规律及影响因素分析[J]. 环境科学学报, 2015, 35(2): 550-556. Xu J, Wang N, Kong D Y, et al. Dynamics and degradation mechanism of antibiotic sulfonamides in soils with organic manure amendment[J]. Acta Scientiae Circumstantiae, 2015, 35(2): 550-556. |
| [35] | 付袁芝.磺胺二甲基嘧啶降解菌的筛选及降解特性研究[D].哈尔滨: 哈尔滨工业大学, 2018. |
| [36] | Shan J, Yang P P, Rahman M M, et al. Tetracycline and sulfamethazine alter dissimilatory nitrate reduction processes and increase N2 O release in rice fields[J]. Environmental Pollution, 2018, 242: 788-796. DOI:10.1016/j.envpol.2018.07.061 |
| [37] | Hou L J, Yin G Y, Liu M, et al. Effects of sulfamethazine on denitrification and the associated N2 O release in estuarine and coastal sediments[J]. Environmental Science & Technology, 2015, 49(1): 326-333. |
| [38] | 鲍陈燕.猪粪对抗生素在农田系统中行为的影响[D].杭州: 浙江大学, 2016. |
| [39] | Hao X, Xu S, Larney F J, et al. Inclusion of antibiotics in feed alters greenhouse gas emissions from feedlot manure during composting[J]. Nutrient Cycling in Agroecosystems, 2011, 89(2): 257-267. DOI:10.1007/s10705-010-9391-3 |
| [40] |
国彬, 姚丽贤, 刘忠珍, 等. 磺胺类兽药对土壤生化功能及氮素的影响[J]. 土壤, 2012, 44(4): 596-600. Guo B, Yao L X, Liu Z Z, et al. Effects of sulfonamide veterinary drugs on soil biochemical function and nitrogen[J]. Soils, 2012, 44(4): 596-600. DOI:10.3969/j.issn.0253-9829.2012.04.011 |
| [41] |
国彬, 姚丽贤, 刘忠珍, 等. 磺胺类兽药对土壤微生物数量的影响[J]. 环境化学, 2012, 31(7): 1009-1015. Guo B, Yao L X, Liu Z Z, et al. The impact of sulfonamide veterinary drugs on soil microbes quantity[J]. Environmental Chemistry, 2012, 31(7): 1009-1015. |
| [42] | Awad Y M, Ok Y S, Igalavithana A D, et al. Sulphamethazine in poultry manure changes carbon and nitrogen mineralisation in soils[J]. Chemistry and Ecology, 2016, 32(10): 899-918. DOI:10.1080/02757540.2016.1216104 |
| [43] |
牛建平, 吴泽辉, 石起增. 磺胺二甲嘧啶在土壤中的降解动态研究[J]. 安徽农业科学, 2009, 37(4): 1767-1769. Niu J P, Wu Z H, Shi Q Z. Research on the degradation dynamic of sulfamethazine in the soil[J]. Journal of Anhui Agricultural Sciences, 2009, 37(4): 1767-1769. DOI:10.3969/j.issn.0517-6611.2009.04.142 |
| [44] | Smith J M, Ogram A. Genetic and functional variation in denitrifier populations along a short-term restoration chronosequence[J]. Applied and Environmental Microbiology, 2008, 74(18): 5615-5620. DOI:10.1128/AEM.00349-08 |
| [45] | Yoshida M, Ishii S, Otsuka S, et al. nirK-harboring denitrifiers are more responsive to denitrification-inducing conditions in rice paddy soil than nirS-harboring bacteria[J]. Microbes and Environments, 2010, 25(1): 45-48. DOI:10.1264/jsme2.ME09160 |
| [46] | Szukics U, Hackl E, Zechmeister-Boltenstern S, et al. Contrasting response of two forest soils to nitrogen input:rapidly altered NO and N2 O emissions and nirK abundance[J]. Biology and Fertility of Soils, 2009, 45(8): 855-863. DOI:10.1007/s00374-009-0396-5 |
| [47] | Morales S E, Cosart T, Holben W E. Bacterial gene abundances as indicators of greenhouse gas emission in soils[J]. The ISME Journal, 2010, 4(6): 799-808. DOI:10.1038/ismej.2010.8 |
| [48] |
曾希柏, 王亚男, 王玉忠, 等. 施肥对设施菜地nirK型反硝化细菌群落结构和丰度的影响[J]. 应用生态学报, 2014, 25(2): 505-514. Zeng X B, Wang Y N, Wang Y Z, et al. Effects of different fertilization regimes on abundance and community structure of the nirK-type denitrifying bacteria in greenhouse vegetable soils[J]. Chinese Journal of Applied Ecology, 2014, 25(2): 505-514. |
| [49] | Kleineidam K, Sharma S, Kotzerke A, et al. Effect of sulfadiazine on abundance and diversity of denitrifying bacteria by determining nirK and nirS genes in two arable soils[J]. Microbial Ecology, 2010, 60(4): 703-707. DOI:10.1007/s00248-010-9691-9 |
| [50] | Kotzerke A, Klemer S, Kleineidam K, et al. Manure contaminated with the antibiotic sulfadiazine impairs the abundance of nirK-and nirS-type denitrifiers in the gut of the earthworm eisenia fetida[J]. Biology and Fertility of Soils, 2010, 46(4): 415-418. DOI:10.1007/s00374-009-0434-3 |
| [51] | Li S Q, Song L N, Jin Y G, et al. Linking N2 O emission from biochar-amended composting process to the abundance of denitrify (nirK and nosZ) bacteria community[J]. AMB Express, 2016, 6: 37. DOI:10.1186/s13568-016-0208-x |
| [52] |
卢静, 刘金波, 盛荣, 等. 短期落干对水稻土反硝化微生物丰度和N2 O释放的影响[J]. 应用生态学报, 2014, 25(10): 2879-2884. Lu J, Liu J B, Sheng R, et al. Effect of short-time drought process on denitrifying bacteria abundance and N2 O emission in paddy soil[J]. Chinese Journal of Applied Ecology, 2014, 25(10): 2879-2884. |