 2019, Vol. 40
2019, Vol. 40

2. 中国科学院亚热带农业生态研究所, 亚热带农业生态过程重点实验室, 长沙 410125
 , XIAO Mou-liang2 , ZHANG Yun1
, XIAO Mou-liang2 , ZHANG Yun1 , YUAN Hong-zhao2 , ZHU Zhen-ke2 , GE Ti-da2 , WU Jin-shui2 , ZHANG Guang-cai1 , GAO Xiao-dan1
, YUAN Hong-zhao2 , ZHU Zhen-ke2 , GE Ti-da2 , WU Jin-shui2 , ZHANG Guang-cai1 , GAO Xiao-dan1
2. Key Laboratory of Subtropical Agriculture Ecology, Institute of Subtropical Agriculture, Chinese Academy of Sciences, Changsha 410125, China
土壤有机碳库是陆地生态系统最大的碳库, 有机碳碳储量相当于大气碳库和植物碳库的2~3倍[1].由于体量巨大, 土壤碳库的微小变动都会对大气CO2浓度和全球气候变化产生重要影响[2]. Wu[3]的研究表明, 在1980~2000年, 我国耕层土壤有机碳量平均每年增加5.78 t·hm-2, 其中稻田土壤贡献了70%的增量.近年30来, 我国南方稻田土壤有机碳含量明显升高[4], 但相关机制仍不清楚.水稻光合碳是土壤有机碳库的最主要的来源之一.通过根际沉积作用, 水稻将约2%~10%的光合碳输入到土壤中[5, 6].作为高活性的有机碳组分, 根际沉积碳对微生物生长代谢有显著的促进作用, 直接影响土壤碳库周转过程.光合碳进入土壤碳库后, 以存在形态的不同可分为颗粒态有机碳(POC)和矿物结合态有机碳(MOC). POC是以游离态存在的有机碳组分, 粒度较大(>53 μm), 易被微生物矿化, 周转速度较快, 一般为几年到几十年[7]. MOC是土壤矿物吸附或包埋的有机碳组分, 粒度小(<53 μm), 生物可接触性差, 周转慢[8].光合碳在POC和MOC间的分配对光合碳在土壤中的稳定有决定性作用, 进而通过调节微生物活动对土壤原有碳库的周转产生重要影响.研究光合碳在水稻植株-土壤间及土壤不同碳库间的分配对土壤有机碳固持与稳定有重要意义.然而其相关研究仍较少, 限制了对稻田土壤碳库累积机制的认识.
磷是水稻生长的必需营养元素, 对水稻光合作用和光合碳在地上部、根部的分配有重要调控作用, 是影响根际沉积碳数量和成分的最主要因素之一[9].热带、亚热带地区是水稻的主产区, 由于长期风化作用, 该区域土壤磷含量普遍较低[10].为了维持较高的作物产量, 磷肥在该区域大量施用.研究发现, 施磷有利于提高叶片中CO2同化率, 增加光合碳在水稻植株中的积累.与不施磷相比, 磷肥(P2O5)施用量在42.0~126.0 kg·hm-2之间, 分蘖期和抽穗期增施磷肥叶片净光合速率分别增加15.17%~29.88%和6.03%~22.38%[10].缺磷胁迫能促进根系发育, 使光合碳向根部的分配比例增大, 提高水稻根冠比[11].施磷条件下, 光合碳向土壤的输送比例下降, 然而由于总光合作用强度的增加, 光合碳向土壤输入的绝对量可能会升高.同时, 与缺磷条件相比, 施磷后, 根系分泌物中对土壤矿物有强吸附性的有机酸含量降低, 对光合碳在土壤碳库中的分配产生潜在影响[12].然而, 到目前为止, 施磷对光合碳在水稻-土壤体系分配影响的系统研究仍很缺乏, 不同磷水平下光合碳在土壤碳库中分配情况的差异鲜见报道.
本文在前人研究的基础上选择低磷红壤水稻土, 采用13CO2连续标记技术来区分光合碳和土壤原有有机碳, 探究施磷对水稻生长过程中光合碳在植株-土壤体系中的分配及其稳定性的影响, 通过进一步揭示稻田生态系统光合碳分配规律, 以期为深入认识农田土壤有机碳累积机制提供科学依据.
1 材料与方法 1.1 供试土壤供试土壤为长期种植水稻的典型水稻土, 由第四纪红色黏土母质发育而成, 有效磷含量<5 mg·kg-1, 根据中国土壤养分分类标准判定为缺磷土壤.土样采自中国科学院长沙农业环境观测研究站(28°33′04″N, 113°19′52″E), 该区海拔92.2~125.3 m, 降雨量1 300 mm, 年平均气温17.5℃, 日照时长1 663 h, 属典型亚热带湿润气候.土样采集时间为2015年4月, 使用不锈钢土钻(直径为5 cm)采集0~20 cm耕作层土壤(13C自然丰度为-29.11‰).将采回的土壤样品运回实验室后, 取1.0 kg土样在室内进行风干, 人工剔除残根和大颗粒物质, 分别过0.250 mm和0.149 mm筛, 用于测定土壤基本理化性质:pH为5.43, 土壤有机质(SOC)14.26 g·kg-1, C/N为9.80, 全氮(TN)为1.45 g·kg-1, 全磷(TP)0.75 g·kg-1, 可溶性磷(Olsen-P)4.96 mg·kg-1, 黏粒(Clay)6.80%, 粉粒(Silt)65.50%, 砂粒(Sand)27.80%;阳离子交换量(CEC)为7.71 cmol·kg-1.取剩余土样过2.00 mm筛, 加超纯水(ddH2O)淹水2~3 cm, 在25℃下预培养14 d, 备用.供试品种为籼型常规双季早稻中早39.
1.2 试验方案盆栽试验采用直径17.2 cm、高20 cm的PVC盆.向预培养后的土壤中加入160 mg·kg-1的KCl(以K计)和250 mg·kg-1的(NH4)2SO4(以N计)作为基肥, 并加入硝化抑制剂DCD(25 mg·kg-1), 充分混匀后平均分为两份, 一份加入NaH2PO4溶液, 添加量以P计为80 mg·kg-1(施磷处理, P80), 另一份加入等量蒸馏水(对照, P0), 分别混匀.将网孔孔径为25 μm的尼龙根际袋(15 cm×17 cm)置于PCV盆中心, 均匀地加入0.5 kg干土当量的P0或P80土壤, 并将另外1.0 kg相同土壤均匀填充在根际袋周围, 作为非根际土.取秧龄20d的水稻幼苗移栽在根际袋中央, 每盆3株, 每个处理4个重复, 在人工气候箱中培养.培养装置和方法参考文献[13, 14], 水稻生长期间用去离子水浇灌, 并及时除草.培养20 d后, 于2015年6月29日将盆栽移入标记箱(80 cm×250 cm ×120 cm)进行标记.13CO2通过13C-NaHCO3(50% atom13C, 1mol·L-1)在CO2发生器中与过量的H2SO4(0.5 mol·L-1)反应产生.通过控制13C-NaHCO3加入速度使标记箱内CO2浓度维持在350~400 μL·L-1.标记箱内外分别安置两个温度传感器(智能型精密数显温度控制器, SNT-96S, 杭州), 箱内安置控温空调, 维持标记过程中标记箱内外温度控制在±1℃[15].与此同时, 另设置2个施肥处理的盆栽水稻在离标记培养箱10 m以外的区域培养, 作为13C的自然丰度对照.
1.3 样品采集在持续淹水条件下(2~3 cm)连续标记14 d, 标记结束后进行破坏性采样:轻轻从PVC盆土壤中取出完整植株, 把收获的水稻植株按根系、地上部分开后, 根系先用自来水冲洗, 再用0.5 mol·L-1 CaCl2(pH 6.2)浸泡1 min, 最后用去离子水冲洗, 以去除吸附在植株表面的土壤, 然后在105℃杀青30 min, 在60℃下烘干至恒重.采集根际袋和根际袋外的土壤分别混匀作为根际土和非根际土, 其余自然风干, 室温保存.
1.4 指标测定土壤基本理化性质:可溶性有机碳(DOC)采用0.5 mol·L-1 K2SO4浸提[16], TOC分析仪测定; 微生物量碳(MBC)采用氯仿熏蒸-TOC分析仪测定法[17].以上指标的测定均使用鲜土.全氮(TN)和有机碳(SOC)的测定采用干烧法-碳氮元素分析仪(VARIO MAX C/N, 德国); 土壤pH值采用Mettler-toledo 320 pH计测定(水土比为2.5:1).以上指标的测定使用风干土样.
土壤有机质采用Canarini等[18]的分级方法, 将土壤有机质分为颗粒态有机碳(POC)和矿物结合态有机碳(MOC).称取10 g风干土于50 mL离心管中, 加入30 mL 5 g·L-1的六偏磷酸钠, 置于摇床中转速180 r·min-1, 振荡15 h, 过53 μm的筛子, 用少量去离子水反复冲洗(4~6次)筛子, 留在筛子上的即为颗粒有机质(>53 μm, POC); 筛子下方的泥浆3 100 g离心后沉淀, 即为矿物态有机物(< 53 μm, MOC).将两种有机质组分60℃烘干, 称重, 计算各组分的比例, 测定全碳和13C.
水稻植株(地上部、根系)与土壤(根际土、非根际土)样品的13C丰度以及植物碳含量测定采用同位素比率质谱仪(MAT253型, 美国热电公司).
1.5 计算公式植株(根系和地上部)和土壤中13C含量的测定由标记样和非标记样中13C的丰度来确定[4]:
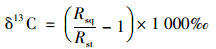
|
(1) |
式中, Rsq表示样品的同位素比值; Rst表示标准物质(PDB)的同位素比值.
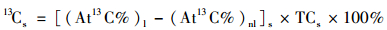
|
(2) |
式中, 13Cs表示样品中13C含量; At13C%表示样品中含13C的丰度; l和nl分别表示标记和非标记样品; TCs表示样品中总碳含量.
生育期水稻光合13C的分配比例计算公式:
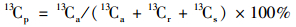
|
(3) |
式中, 13Cp是指水稻地上部光合13C在水稻-土壤系统中的分配比例(%); 13Ca、13Cr和13Cs分别指地上部、地下部和土壤中的13C含量.其他部分的分配比例计算方法类似.
水稻光合13C各部分光合13C含量的总和(13Ca+13Cr+13Cs), 假设同位素分馏效应可以忽视, 用13C光合碳的分配情况代表总光合碳的分配.
1.6 数据处理数据处理和统计分析采用Microsoft Excel 2013和SPSS 20软件.绘图采用Origin8.5软件.不同处理差异显著性用单因素方差分析(One-way ANOVA)、t检验.
2 结果与分析 2.1 水稻生物量和根冠比施磷对水稻地上部和根系的生物量无显著影响, 但在连续标记14 d后, 不施磷的水稻总生物量(3.19 g·pot-1)显著高于施磷处理(3.70 g·pot-1), 施磷显著降低了水稻生物量的根冠比, 降幅达34%(P < 0.05, 图 1).不施磷的水稻植株全碳含量为1.22 g·pot-1, 施磷的全碳含量为1.53 g·pot-1, 施磷使水稻地上部全碳含量, 增幅为25%;不施磷的根系全碳含量为0.55 g·pot-1, 施磷的根系全碳含量为0.40 g·pot-1, 施磷显著降低了根系的全碳含量, 降幅为27%.因此, 施磷显著增加了地上部的生物量和全碳含量, 减少了地下部的全碳含量, 显著降低了水稻根冠比(图 1).

|
图 1 水稻干物质量和生物量碳 Fig. 1 Dry weight and biomass carbon of rice |
施磷显著增加水稻地上部13C的分配, 降低13C在水稻根系和根际土壤的比例, 但对非根际土壤的13C分配无显著影响.施磷处理光合碳向地上部13C分配与不施磷相比增加了39%, 从而使根际13C分配降低了32%(P < 0.05, 图 2).与不施磷相比, 施磷使13C向地上部的分配比例增加了24%;向根系和根际土壤的分配比例分别减少了42%和43%(P < 0.05, 图 2).综上, 施磷促进光合碳在地上部的分配, 而降低光合碳向根系和根际土壤的传输(P < 0.05, 图 2), 但对其在非根际土壤的分配无显著性影响.

|
图 2 13C在水稻-土壤系统中的含量及分配比例 Fig. 2 Amount and proportion of 13C in the rice-soil system |
如图 3所示, 在连续标记14 d后, 施磷对根际土壤13C-MBC含量无显著影响, 但使光合碳在根际土壤MBC分配比例显著降低了30%(P < 0.05).另外, 施磷条件下, 根际土壤13C-MBC含量和分配比例均显著高于非根际土壤, 增幅达0.92 mg·kg-1和79%(P < 0.05).在非根际土壤中, 施磷使13C-MBC含量显著提高了78%(P < 0.05);而在根际土壤中, 施磷处理显著降低了13C-MBC的含量0.99 mg·kg-1, 分配比例降低了30%(P < 0.05).

|
图 3 根际和非根际土壤中13C-MBC的含量及其分配比例 Fig. 3 Amount and proportion of 13C-MBC in the rhizosphere and non-rhizosphere soil |
施磷对根际和非根际土壤的13C-MOC含量和分配比例均无显著影响.施磷显著降低根际土壤13C-POC含量和分配比例, 分别降低了1.43 mg·kg-1和52 %(P < 0.01, 图 4), 但对非根际土壤的13C-POC含量和分配比例无显著性差异.在施磷条件下, 根际土壤的13C-MOC含量和分配比例比非根际土壤分别提高了0.67 mg·kg-1和230%, 不施磷处理分别提高了0.67 mg·kg-1和200%.在施磷条件下, 根际土壤13C-POC含量和分配比例与13C-MOC相比显著性提高1.07 mg·kg-1和110%, 不施磷处理分别提高了2.46 mg·kg-1和240%(P < 0.01, 图 4).相关性分析表明, 13C在土壤的各碳库(SOC、POC、MOC和MBC)中的分配具有显著相关性(表 1).方差分析结果表明, 施磷和根际/非根际的效应(R/B)显著影响13C在SOC、POC和MBC中的分配比例(表 2).

|
图 4 13C在颗粒态有机碳和矿物结合态有机碳中的含量及其分配比例 Fig. 4 Content of 13C in the POC and MOC and its distribution ratio |
|
|
表 1 13C-MOC、13C-POC、13C-SOC和13C-MBC含量间的相关相关性分析 Table 1 Correlation analyses of the 13C-MOC, 13C-POC, 13C-SOC, and 13C-MBC content |
|
|
表 2 施磷及根际与非根际(R/B)对13C在土壤各碳库中的分配影响的方差分析1) Table 2 ANOVA analyses for P fertilization and the rhizosphere effect for the incorporation of assimilated 13C in the soil C pools |
3 讨论 3.1 水稻生物量及其根冠比对施磷的响应
磷素是水稻生长必需的大量元素, 水稻的生长发育受到磷供给的限制, 土壤磷素供应量的变化通常可引起水稻生物量的变化, 并影响植物体内光合碳的分配格局[19].本研究发现, 在水稻拔节期, 施磷处理的植株全碳含量显著高于不施磷组, 说明施磷有利于水稻光合碳的累积.然而, 施磷条件下光合碳的累积主要集中在地上部, 根部全碳含量显著下降.光合碳在地上部和根部的分配情况对土壤磷水平的响应是水稻应对缺磷胁迫的重要手段之一.土壤是植物磷素的唯一来源, 根系是土壤磷素进入植物体内的唯一途径.缺磷条件下, 植株将大量的光合碳分配到地下部, 用于根尖伸长和细根发育, 有效增大了根系与土壤的接触面积, 有利于提高对磷素的获取能力.与本研究结果相似, Mehra等[20]发现施用磷肥对根系生长有抑制作用, 另外与缺磷条件相比, 磷素供应充足时, 水稻根冠比显著降低.
3.2 施磷对光合碳在土壤-水稻系统分配的影响水稻植株主要通过叶片进行光合作用将13C固定在植物体内, 并在水稻-土壤系统中进行再分配[21, 22], 植株和土壤13C含量的变化清晰地反映了光合碳的转移路径.光合碳的分配受到作物生长环境的影响, 而施磷作为水稻生长的重要调控因子, 对光合碳在水稻-土壤系统中的分配有重要意义.本研究发现施磷显著提高了光合碳在水稻植株中的分配比例, 降低了其向土壤的转移(图 2).有研究表明, 水稻根际沉积碳的输入与根系生物量显著正相关[23].本研究中, 施用磷肥后使根系生物量减少, 从而显著降低了水稻光合碳向根际土壤的输入.根际沉积作用是植物实现土壤磷活化的主要的机制之一.磷素供应不足时, 根系柠檬酸、乙酸、琥珀酸等小分子有机酸和磷酸酶的分泌量显著增加[24], 以促进对无机磷的溶出和有机磷的矿化, 提高土壤磷素的生物有效性[25].除了少量光合碳通过真菌根毛等传输外[26], 根际土壤是光合碳向土壤释放的主要场所.从而使施磷和不施磷处理中根际土13C含量均显著高于非根际土13C含量.13C在根际土壤MBC中施磷带来的影响并不显著, 然而在非根际土壤中, 施磷后的13C-MBC显著高于不施磷处理(图 3), 这可能与微生物代谢底物的有效性有关.根系分泌物多数是一些易利用态的小分子物质, 优先被微生物利用.在土壤养分不易于被根系吸收的非根际土壤中, 微生物通将光合碳同化自身物质保存于土壤中.然而当养分缺乏时, 微生物偏向于通过降解、矿化光合碳, 以获得维持生长所需能量[27].从而不利于光合碳在非根际土壤的固持.
3.3 光合碳在土壤中的稳定及其对施磷的响应土壤有机碳与矿物结合是使其稳定化的重要机制[28], 提高土壤中MOC比例被认为是抑制土壤有机碳分解、缓解大气CO2浓度上升的重要途径[29].光合碳在根际土壤中主要以POC形态存在, 而非根际土中, POC和MOC的13C含量差异较小.这主要由根际沉积物的传输性所致, 尤其是粒径较大的颗粒有机物, 在土壤中的传输受限, 造成其含量在根际和非根际土壤中显著的差异.施磷后, 光合碳在根际土壤POC的分配显著降低, 这与王玲莉等[30]和刘骅等[31]的研究结果一致.该现象可能由两方面原因导致:其一, 施磷后根际沉积碳总量减少, 向POC的分配随之减少; 其二, 施肥使根系分泌物的组分发生改变[32], 使其更利于被微生物降解.特别是在根际热区, 微生物与根系对于养分的竞争尤为激烈[33].本试验施肥是通过基肥一次加入, 在养分后续不足的情况下, 通过激发效应, 前期被激活的微生物大量降解土壤有机质以获取养分, 此过程也包括来源于光合碳的已被利用的POC.佟小刚等[34]和樊廷录等[35]报道长期施有机肥条件下红壤、黑垆土矿物结合态有机碳含量有所增加, 原因在于施有机肥为微生物提供了营养, 微生物代谢分泌物增多, 这些分泌物可以直接转移至细黏粒, 导致土壤矿物结合态有机碳积累.这表明微生物在POC转化为MOC的过程中发挥重要的作用, 有利于光合碳在土壤中的稳定.
4 结论缺磷稻田土壤施磷后显著促进水稻地上部分生物量的增加, 却限制了根系的生长, 降低了水稻根冠比, 使水稻光合碳向地下部(根系+土壤)的分配显著减小.可见, 虽然施磷提高了水稻植株, 尤其是地上部的生长, 但降低了光合碳在土壤中的积累.光合碳在根际土壤微生物生物量碳(MBC)、颗粒态有机碳(POC)和矿物结合态有机碳(MOC)的分配均显著高于非根际土壤, 表明水稻根际土壤是光合碳输入、周转和积累的主要区域.此外, 施磷对光合碳向非根际土壤POC和MOC的分配无显著影响, 却显著降低其在根际土壤POC中的分配, 表明磷肥的施用降低了根际沉积碳的周转潜力, 有利于水稻光合碳在土壤中的稳定.
| [1] | Pan G X, Li L Q, Wu L S, et al. Storage and sequestration potential of topsoil organic carbon in China's paddy soils[J]. Global Change Biology, 2004, 10(1): 79-92. DOI:10.1111/gcb.2004.10.issue-1 |
| [2] |
陈春梅, 谢祖彬, 朱建国. 大气CO2浓度升高对土壤碳库的影响[J]. 中国生态农业学报, 2008, 16(1): 217-222. Chen C M, Xie Z B, Zhu J G. Effects of elevated atmospheric CO2 concentration on soil carbon[J]. Chinese Journal of Eco-Agriculture, 2008, 16(1): 217-222. |
| [3] | Wu J. Carbon accumulation in paddy ecosystems in subtropical China:evidence from landscape studies[J]. European Journal of Soil Science, 2011, 62(1): 29-34. |
| [4] |
任逸文, 肖谋良, 袁红朝, 等. 水稻光合碳在植物-土壤系统中的分配及其对CO2升高和施氮的响应[J]. 应用生态学报, 2018, 29(5): 1397-1404. Ren Y W, Xiao M L, Yuan H C, et al. Allocation of rice photosynthates in plant-soil system in response to elevated CO2 and nitrogen fertilization[J]. Chinese Journal of Applied Ecology, 2018, 29(5): 1397-1404. |
| [5] | Ge T D, Yuan H Z, Zhu H H, et al. Biological carbon assimilation and dynamics in a flooded rice-soil system[J]. Soil Biology and Biochemistry, 2012, 48(4): 39-46. |
| [6] |
陈珊, 祝贞科, 袁红朝, 等. 拔节期水稻光合碳输入的动态变化及其对施氮的响应:13C-CO2脉冲标记[J]. 环境科学, 2018, 39(1): 331-338. Chen S, Zhu Z K, Yuan H Z, et al. Dynamics of rice photosynthesized carbon input and its response to nitrogen fertilization at the jointing stage:13C-CO2 Pulse-labeling[J]. Environmental Science, 2018, 39(1): 331-338. |
| [7] | Cambardella C A, Elliott E T. Carbon and nitrogen distribution in aggregates from cultivated and native grassland soils[J]. Soil Science Society of America Journal, 1993, 57(4): 1071-1076. DOI:10.2136/sssaj1993.03615995005700040032x |
| [8] | Watson J R, Parsons J W. Studies of soil organo-mineral franctions:I. Isolation by ultrasonic dispersion[J]. Journal of Soil Science, 1974, 25(1): 1-8. |
| [9] |
柏菁, 李奕霏, 刘守龙, 等. 添加磷素对低磷稻田根际土壤固碳自养微生物数量的影响[J]. 环境科学, 2018(12): 1-12. Bai J, Li Y F, Liu S L, et al. Effect of phosphorus addition on the abundance of autotrophic CO2 fixation microorganisms in the rhizospheric soil from a phosphorus limited paddy field[J]. Environmental Science, 2018(12): 1-12. DOI:10.3969/j.issn.1673-1212.2018.12.001 |
| [10] |
王婷婷, 祝贞科, 朱捍华, 等. 施氮和水分管理对光合碳在土壤-水稻系统间分配的量化研究[J]. 环境科学, 2017, 38(3): 1227-1234. Wang T T, Zhu Z K, Zhu H H, et al. Input and distribution of photosynthesized carbon in soil-rice system affected by water management and nitrogen fertilization[J]. Environmental Science, 2017, 38(3): 1227-1234. |
| [11] |
刘玉槐, 魏晓梦, 魏亮, 等. 水稻根际和非根际土磷酸酶活性对碳、磷添加的响应[J]. 中国农业科学, 2018, 51(9): 1653-1663. Liu Y H, Wei X M, Wei L, et al. Responses of extracellular enzymes to carbon and phosphorus additions in rice rhizosphere and bulk soil[J]. Scientia Agricultura Sinica, 2018, 51(9): 1653-1663. |
| [12] | Pinheiro, érika Flávia Machado, et al. Tillage systems effects on soil carbon stock and physical fractions of soil organic matter[J]. Agricultural Systems, 2015, 132: 35-39. DOI:10.1016/j.agsy.2014.08.008 |
| [13] | Ge T, Chen X, Yuan H, et al. Microbial biomass, activity, and community structure in horticultural soils under conventional and organic management strategies[J]. European Journal of Soil Biology, 2013, 58: 122-128. DOI:10.1016/j.ejsobi.2013.07.005 |
| [14] | Ge T D, Wu X H, Chen X J, et al. Microbial phototrophic fixation of atmospheric CO2 in China subtropical upland and paddy soils[J]. Geochimica et Cosmochimica Acta, 2013, 113: 70-78. DOI:10.1016/j.gca.2013.03.020 |
| [15] | Chapin Ⅲ F S, Bloom A J, Field C B, et al. Plant responses to multiple environmental factors:physiological ecology provides tools for studying how interacting environmental resources control plant growth[J]. Bioscience, 1987, 37(1): 49-57. DOI:10.2307/1310177 |
| [16] |
张瑞, 张贵龙, 姬艳艳, 等. 不同施肥措施对土壤活性有机碳的影响[J]. 环境科学, 2013, 34(1): 277-282. Zhang R, Zhang G L, Ji Y Y, et al. Effects of different fertilizer application on soil active organic carbon[J]. Environmental Science, 2013, 34(1): 277-282. |
| [17] | Yang J, Kang Y, Sakurai K, et al. Fixation of carbon dioxide by chemoautotrophic bacteria in grassland soil under dark conditions[J]. Acta Agriculturae Scandinavica, Section B-Soil & Plant Science, 2017, 67(4): 362-371. |
| [18] | Canarini A, Dijkstra F A. Dry-rewetting cycles regulate wheat carbon rhizodeposition, stabilization and nitrogen cycling[J]. Soil Biology and Biochemistry, 2015, 81: 195-203. DOI:10.1016/j.soilbio.2014.11.014 |
| [19] |
祝贞科, 沈冰洁, 葛体达, 等. 农田作物同化碳输入与周转的生物地球化学过程[J]. 生态学报, 2016, 36(19): 5987-5997. Zhu Z K, Shen B J, Ge T D, et al. Biogeochemical processes underlying the input and turnover of crop assimilative carbon in farmland ecosystems[J]. Acta Ecologica Sinica, 2016, 36(19): 5987-5997. |
| [20] | Mehra P, Pandey B K, Giri J. Comparative morphophysiological analyses and molecular profiling reveal pi-efficient strategies of a traditional rice genotype[J]. Frontiers in Plant Science, 2016, 6: 1184. |
| [21] | Johnson D, Leake J R, Ostle N P, et al. In situ 13CO2 pulse-labelling of upland grassland demonstrates a rapid pathway of carbon flux from arbuscular mycorrhizal mycelia to the soil[J]. New Phytologist, 2002, 153(2): 327-334. DOI:10.1046/j.0028-646X.2001.00316.x |
| [22] | Kuzyakov Y, Domanski G. Model for rhizodeposition and CO2 efflux from planted soil and its validation by 14 C pulse labelling of ryegrass[J]. Plant and Soil, 2002, 239(1): 87-102. DOI:10.1023/A:1014939120651 |
| [23] | Ge T D, Liu C, Yuan H Z, et al. Tracking the photosynthesized carbon input into soil organic carbon pools in a rice soil fertilized with nitrogen[J]. Plant and Soil, 2015, 392(1-2): 17-25. DOI:10.1007/s11104-014-2265-8 |
| [24] | Richardson A E, Barea J, McNeill A M, et al. Acquisition of phosphorus and nitrogen in the rhizosphere and plant growth promotion by microorganisms[J]. Plant and Soil, 2009, 321(1-2): 305-339. DOI:10.1007/s11104-009-9895-2 |
| [25] | Nuruzzaman M, Lambers H, Bolland M D A, et al. Distribution of carboxylates and acid phosphatase and depletion of different phosphorus fractions in the rhizosphere of a cereal and three grain legumes[J]. Plant and Soil, 2006, 281(1-2): 109-120. DOI:10.1007/s11104-005-3936-2 |
| [26] | Hafner S, Wiesenberg G L B, Stolnikova E, et al. Spatial distribution and turnover of root-derived carbon in alfalfa rhizosphere depending on top-and subsoil properties and mycorrhization[J]. Plant and Soil, 2014, 380(1-2): 101-115. DOI:10.1007/s11104-014-2059-z |
| [27] | Yin L M, Dijkstra F A, Wang P, et al. Rhizosphere priming effects on soil carbon and nitrogen dynamics among tree species with and without intraspecific competition[J]. New Phytologist, 2018, 218(3): 1036-1048. DOI:10.1111/nph.2018.218.issue-3 |
| [28] |
马欣, 魏亮, 唐美玲, 等. 长期不同施肥对稻田土壤有机碳矿化及激发效应的影响[J]. 环境科学, 2018(12): 1-12. Ma X, Wei L, Tang M L, et al. Effects of different long-term fertilization on organic carbon mineralization and priming effect of paddy soil[J]. Environmental Science, 2018(12): 1-12. DOI:10.3969/j.issn.1673-1212.2018.12.001 |
| [29] |
袁颖红, 李辉信, 黄欠如, 等. 不同施肥处理对红壤性水稻土微团聚体有机碳汇的影响[J]. 生态学报, 2004, 24(12): 2961-2966. Yuan Y H, Li H X, Huang Q R, et al. Effects of different fertilization on soil organic carbon distribution and storage in micro-aggregates of red paddy topsoil[J]. Acta Ecologica Sinica, 2004, 24(12): 2961-2966. DOI:10.3321/j.issn:1000-0933.2004.12.045 |
| [30] |
王玲莉, 韩晓日, 杨劲峰, 等. 长期施肥对棕壤有机碳组分的影响[J]. 植物营养与肥料学报, 2008, 14(1): 79-83. Wang L L, Han X R, Yang J F, et al. Effect of long-term fertilization on organic carbon fractions in a brown soil[J]. Plant Nutrition and Fertilizer Science, 2008, 14(1): 79-83. DOI:10.3321/j.issn:1008-505X.2008.01.013 |
| [31] |
刘骅, 佟小刚, 马兴旺, 等. 长期施肥下灰漠土矿物颗粒结合有机碳的含量及其演变特征[J]. 应用生态学报, 2010, 21(1): 84-90. Liu H, Tong X G, Ma X W, et al. Content and evolution characteristics of organic carbon associated with particle-size fractions of grey desert soil under long-term fertilizaton[J]. Chinese Journal of Applied Ecology, 2010, 21(1): 84-90. |
| [32] | Kaštovská E, Edwards K, Šantrůč ková H. Rhizodeposition flux of competitive versus conservative graminoid:contribution of exudates and root lysates as affected by N loading[J]. Plant and Soil, 2017, 412(1-2): 331-344. DOI:10.1007/s11104-016-3066-z |
| [33] | Kuzyakov Y, Blagodatskaya E. Microbial hotspots and hot moments in soil:Concept & review[J]. Soil Biology and Biochemistry, 2015, 83: 184-199. DOI:10.1016/j.soilbio.2015.01.025 |
| [34] |
佟小刚, 徐明岗, 张文菊, 等. 长期施肥对红壤和潮土颗粒有机碳含量与分布的影响[J]. 中国农业科学, 2008, 41(11): 3664-3671. Tong X G, Xu M G, Zhang W J, et al. Influence of long-term fertilization on content and distribution of organic carbon in particle-size fractions of red soil and fluvo-aquic soil in China[J]. Scientia Agricultura Sinica, 2008, 41(11): 3664-3671. DOI:10.3864/j.issn.0578-1752.2008.11.029 |
| [35] |
樊廷录, 王淑英, 周广业, 等. 长期施肥下黑垆土有机碳变化特征及碳库组分差异[J]. 中国农业科学, 2013, 46(2): 300-309. Fan T L, Wang S Y, Zhou G Y, et al. Effects of long-term fertilizer application on soil organic carbon change and fraction in cumulic haplustoll of loess plateau in China[J]. Scientia Agricultura Sinica, 2013, 46(2): 300-309. DOI:10.3864/j.issn.0578-1752.2013.02.009 |