 2019, Vol. 40
2019, Vol. 40

2. 中国科学院亚热带农业生态研究所亚热带农业生态过程重点实验室, 长沙 410125;
3. 中南林业科技大学环境科学与工程学院, 长沙 410004
 , WANG Ji-fei1,2 , ZHU Zhen-ke2 , DENG Yang-wu1
, WANG Ji-fei1,2 , ZHU Zhen-ke2 , DENG Yang-wu1 , CHEN Shan2,3 , GE Ti-da2 , YUAN Hong-zhao2 , WU Jin-shui2
, CHEN Shan2,3 , GE Ti-da2 , YUAN Hong-zhao2 , WU Jin-shui2
2. Key Laboratory of Subtropical Agriculture Ecology, Institute of Subtropical Agriculture, Chinese Academy of Sciences, Changsha 410125, China;
3. College of Environmental Science and Engineering, Central South University of Forestry and Technology, Changsha 410004, China
土壤有机碳大约占陆地生态系统有机碳储存的四分之三, 其含量是表征土壤肥力的重要指标, 在全球碳循环中扮演重要角色[1].在稻田生态系统中, 有机碳的积累可以改善土壤的团聚体结构[2], 提高土壤质量和耕作土地作物的生产力[3], 而且有利于缓解地球温室效应[4], 其有机碳库大小取决于碳输入和碳输出的动态变化.施肥、秸秆还田和根系分泌物等是碳输入的主要方式, 而土壤碳输出的主要途径是外源有机碳的分解和土壤原有有机质的矿化[5].有机碳矿化是由微生物驱动, 相关酶介导的分解有机质释放CO2等气体的生物化学过程[6, 7].外源有机碳输入可引起土壤有机质周转短期内强烈变化的现象称为激发效应[8].激发效应与土壤有机碳的分解和积累密切联系, 可反映外源有机碳对土壤原有有机碳矿化作用的强度和方向, 是表征土壤碳循环的重要指标.目前, 关于土壤有机碳矿化及其激发效应的研究多为添加外源有机质, 如牛粪[9]、生物碳[10]、秸秆[11]等, 其添加方式和添加量因土壤和实验目的而异, 因此关于有机碳矿化及其激发效应的机制尚不明确.
在植物根系、微生物和土壤有机碳库之间的相互作用中, 微生物具有重要的介导作用; 外源碳底物添加量与土壤微生物生物量碳(MBC)存在一定的关系, 外源碳量相对于MBC的比例称为“碳负荷”[12], 同时微生物对底物碳的转换率是碳固存的重要因素[13]. Blagodatskaya等[12]探讨微生物量和微生物结构对表观激发相应和真实激发效应的机制, 发现外源添加碳数量高于、等于和低于MBC量对土壤微生物活性的影响不同, 并提出计算土壤碳底物时应该考虑土壤微生物碳量. Wang等[6]提出新鲜有机质的输入量会影响微生物对底物的利用, 以及对土壤有机质的矿化.另外, Guenet等[14]的研究证明新鲜有机质的输入与激发效应不存在线性关系.然而对于微生物、外源碳输入量以及激发相应三者之间的关系研究甚少.因此, 以土壤微生物碳量为微生物对碳的需求量大小的依据, 采用分梯度添加方式来研究微生物对土壤碳矿化和激发效应的作用规律, 可以深入揭示微生物活性碳库对外源碳输入量的响应特征.
葡萄糖是一种易利用态的糖类有机碳, 约占植物根系分泌的碳水化合物的一半, 是微生物生长的主要碳源[15, 16].此外, 微生物呼吸响应水平随葡萄糖负荷速率的增加而增加, 直到微生物的碳需求达到饱和为止[17].因此, 选择葡萄糖为典型外源有机碳, 研究在不同碳负荷梯度下, 稻田有机碳矿化及其激发效应, 进而深入探讨土壤碳转化的调控机制.
1 材料与方法 1.1 供试土壤概况供试土壤为多年种植水稻的典型红壤, 采集于中国科学院长沙农业环境观测研究站(113°19′52″E, 28°33′04″N), 该地区为亚热带季风湿润气候, 平均气温16~18℃, >10℃积温5 000~5 800℃, 无霜期260~310 d, 年降雨量1 200~1 700 mm.采用直径为5 cm不锈钢土钻采集0~20 cm的耕作层土壤, 除去可见杂质(植物残体、石头等), 然后室内自然风干, 分别过2 mm和0.149 mm筛, 用于测定土壤基本理化性质[18].剩余土样风干后过2 mm筛, 备用.供试土壤基本理化性质: pH为6.0, 黏粒含量(质量分数)14.1%, 砂粒含量6.7%, 粉粒79.2%, 土壤有机碳(SOC)18.6 g·kg-1, 土壤可溶性有机碳(DOC)18.5g·kg-1, 碳氮比(C/N)10:7, 碱解氮0.13g·kg-1, 微生物碳量(MBC)0.63g·kg-1.
1.2 土壤培养实验为研究在不同碳负荷条件下, 微生物对稻田土壤有机碳及其激发效应的作用, 本实验共设计了5个添加处理:①对照(CK), ②土+0.5×MBC, ③土+1×MBC, ④土+3×MBC, ⑤土+5×MBC, 每处理设置3个重复.将过2 mm的土壤置于50 L塑料桶中, 淹水2~3 cm, 25℃恒温预培养14 d.称取预培养后混匀的土样20 g(干土计)于500 mL厌氧瓶中, 添加上述含量的13C-葡萄糖溶液, 淹水培养(干土/水=1/2), 置于25℃恒温培养室.在培养的前15 d每天采集气体, 之后分别在第18、22、26、30、35、40、50和60 d采集气体.采气方法:选用一次性注射器采集气体30 mL到已抽真空的顶空瓶中, 利用鼓气装置将厌氧瓶中剩余气体鼓出, 待剩余气体排尽后, 重新密封并置于恒温培养室继续培养.
另称取土样200 g(干重)于培养瓶中在上述相同条件下, 每隔5 d开盖通气, 并在2 d和60 d进行破坏性采样, 测定其NH4+-N和MBN等理化指标, 并采集10 g鲜土样于-20℃保存, 短期内测定土壤酶活性指标.
1.3 测定与分析方法土壤pH测定以水为浸提剂, 水土比为2.5:1, 采用Mettler-toledo320pH计(FE20K, 瑞士)测定; 土壤机械组成采用激光粒度仪测定.土壤有机碳(SOC)和全氮(TN)采用碳氮元素分析仪(VARIO MAXC/N, 德国)测定(干烧法). NH4+-N采用流动注射仪(Fiastar5000, 瑞典福斯)测定.生物量碳(MBC)采用氯仿熏蒸提取-碳自动分析法测定.土壤溶解性有机碳(DOC, mg·kg-1)采用有机碳分析仪(岛津Vwp, 日本)测定.土壤氮转化酶活性的测定采用96微孔酶标板荧光分析法[19]. CO2气体含量用气相色谱(Shimadzu, Kyoto, 日本)测定.土壤、葡萄糖和CO2的13C丰度用MAT253同位素质谱仪(Thermo Fisher Scientific, Waltham, US)测定.
1.4 数据处理添加13C-葡萄糖于稻田土壤中培养, 土壤原有碳和外加葡萄糖矿化作用产生CO2气体, 计算公式[20]见式(1)和式(2).
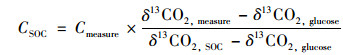
|
(1) |
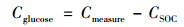
|
(2) |
式中, CSOC是来源SOC的CO2量, mg·kg-1; Cmeasure是测量的总CO2释放量, mg·kg-1; Cglucose是外源碳分解产生的CO2量, mg·kg-1; δ13CO2, measure是总CO2测量的13C丰度; δ13CO2, SOC是SOC分解释放的13C丰度(对照组释放的13C丰度), δ13CO2, glucose是添加至样品中的13C丰度.
葡萄糖-C的累积矿化量是各取样点CO2生成量的总和, 葡萄糖-C的矿化率用13C-CO2的量与加入土壤中葡萄糖-13C总量的比值表示.
激发效应(PE)计算公式[21]见式(3).
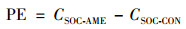
|
(3) |
式中, CSOC-AME表示添加葡萄糖处理中来源于土壤有机碳(SOC)的CO2的量, mg·kg-1; CSOC-CON表示对照处理中来源于土壤有机碳的CO2量, mg·kg-1.
土壤中易被微生物迅速分解利用的那部分活性碳为快库(a1), 而暂时固定于微生物体内以及被土壤矿物或团聚体稳定的相对难以利用的惰性碳为慢库(a2).双库模型计算公式见式(4).
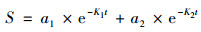
|
(4) |
式中, a1和a2分别为快库和慢库的比例(%); K1和K2分别为快库和慢库的矿化速率常数(d-1); S为累积矿化量; t为培养时间.在双库模型中假设a1+a2=100%, 利用60 d内累积矿化量进行双库模型的拟合得出参数a1、a2、K1、K2.
土壤有机碳的半衰期(T1/2/d)计算:
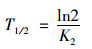
|
(5) |
土壤快库的平均滞留时间(MRT/d)计算如下:
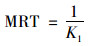
|
(6) |
测定结果均为3次重复的平均值, 所得数据采用Microsoft Excel 2010处理、采用Origin 8.5绘图, 采用Sigma plot 12.5软件进行双库模型拟合, 运用SPSS 16.0软件进行各因子间相关性分析, 通过Canoco5实现环境因子的RDA(冗余分析).
2 结果与分析 2.1 不同碳负荷梯度下土壤外源碳的矿化有机碳累积矿化总量随着葡萄糖-C负荷的增加而增加, 其矿化率在60 d培养期内呈两个阶段变化趋势:在0~20 d快速分解, 而后逐渐达到稳定(图 1).葡萄糖添加显著增加了土壤碳的矿化, 相比于不添加外源碳的处理(CK, 587.8mg·kg-1), 葡萄糖-C的添加促进了CO2释放, 分别提高了1.4倍(0.5×MBC)、2.0倍(1×MBC)、3.2倍(3×MBC)和4.2倍(5×MBC)[图 1(a)].土壤中葡萄糖-C快速矿化生成13CO2; 相比于高负荷处理, 低负荷处理快速矿化, 10 d左右达到稳定状态; 高负荷下13CO2-C累积排放量在20d左右达到稳定[图 1(b)].负荷为0.5×MBC处理有852.2mg·kg-1的13CO2-C生成, 而1×MBC、3×MBC和5×MBC处理分别是0.5×MBC处理的2.7、9.8、16倍.

|
图 1 不同碳负荷梯度下葡萄糖矿化产生CO2和13CO2-C规律 Fig. 1 Cumulative CO2 and 13CO2-C production from mineralization of glucose relative to carbon loading |
土壤添加的易利用态碳快速分解, 在60 d培养实验结束时, 葡萄糖-C矿化为34.9%~56.7%[图 2(a)]; 葡萄糖-C累积矿化率随添加负荷量的增加而呈指数增加[图 2(b)].负荷为0.5×MBC和1×MBC处理中矿化率平稳在30%和40%左右, 而负荷3×MBC和5×MBC的处理中矿化率可达53%左右[图 2(b)].同样, 在60 d培养结束时, 低负荷的矿化率比高负荷的矿化率要更低, 而且在负荷过高时葡萄糖-C矿化率不再变化, 如图 2(b)所示, 不同负荷葡萄糖-C累积矿化率符合一级动力学方程的变化规律(R2=0.99, P < 0.05).

|
图 2 不同碳负荷梯度下的葡萄糖-C矿化率和葡萄糖60 d累积矿化率与易利用态碳负荷之间的关系 Fig. 2 Cumulative glucose mineralization rate at the end of 60d incubation and relationships of carbon loading with the cumulative glucose-C mineralization rate |
葡萄糖的输入负荷不仅可以改变其矿化率, 而且改变了其在土壤中快库和慢库间的分配.利用一级动力学双指数模型拟合葡萄糖-C在土壤碳库中分配的动态变化[22], 由图 3可知, 随着易利用态碳负荷的增加, 葡萄糖-C向土壤快库分配的比例逐渐由26%增加至56%;而向慢库分配的比例由72%降低至42%, 土壤中葡萄糖-C快库、慢库的比例和不同负荷易利用态碳之间呈现指数关系(R2=0.99, P < 0.05和R2=0.99, P < 0.05), 当葡萄糖-C添加量大于3×MBC时, 其分配比例不再受输入负荷的影响(图 3).另外, 由表 1可以看出, 碳滞留时间在低负荷条件下差异不大, 但在高负荷条件下有明显的差异.

|
图 3 土壤中葡萄糖-C快库与慢库的比例及其与碳负荷之间的关系 Fig. 3 Relationship of carbon loading with the ratio of readily available and slowly available C in 13C glucose |
|
|
表 1 不同碳负荷易梯度下的生物可利用标记的碳库、碳持留时间和CO2中13C的半衰期1) Table 1 Bioavailable labeled substrate C pool, mean residence time (MRT), and half-life of cumulative 13C recovery in CO2 under different gradient carbon loading |
2.3 不同碳负荷梯度下的累积激发效应
在60 d的培养过程中, 所有输入负荷下, 葡萄糖对土壤原有有机碳的矿化均表现为正累积激发效应(图 4).在实验初期(前15 d), 激发效应随葡萄糖负荷的升高而增强.随着培养时间, 累积激发效应逐渐增加. 20 d后, 5×MBC处理出现轻微负激发效应, 其他处理中激发效应依然为正; 负荷为3×MBC虽然比1×MBC处理增加了两倍, 但是60 d累积激发效应却下降了20.7%.培养60 d时, 1×MBC处理累积激发效应最大, (325.1 mg·kg-1), 而负荷为0.5×MBC处理累积激发效应最小(160.0 mg·kg-1).

|
图 4 不同碳负荷梯度下的累积激发效应 Fig. 4 Cumulative priming effect of the initial 60 days |
冗余分析(RDA)表明, 前期其累积激发效应主要是受MBC、MBN以及DOC影响, 后期主要是受β-葡糖苷酶、几丁质酶以及铵态氮的影响(图 5).这与不同负荷累积激发效应前期呈正相关和后期出现差异结果一致(图 4).同时, 对2 d和60 d的不同碳负荷梯度的累积矿化率和累积激发效应与土壤基本指标进行Pearson相关分析发现(表 2和表 3), 在2 d时累积矿化率和累积激发效应与MBC呈正相关(R=0.24, P < 0.01; R=0.36, P < 0.01), 而累积激发效应与DOC呈显著正相关关系(R=0.96, P < 0.01), 在2d累积矿化率与β-葡糖苷酶和几丁质酶活性呈显著负相关(R=-0.76, P < 0.01; R=-0.87, P < 0.01), 在60 d累积矿化率与β-葡糖苷酶和几丁质酶活性呈显著正相关(R=0.78, P < 0.01; R=0.72, P < 0.01).

|
图 5 影响累积激发效应因素的RDA分析 Fig. 5 RDA of influencing factors on the cumulative priming effect |
|
|
表 2 d累积激发强度和土壤基本指标的Pearson相关分析1) Table 2 Pearson correlation analysis of priming effect intension and soil basic index in 2 days |
|
|
表 3 60 d累积激发强度和土壤基本指标的Pearson相关分析 Table 3 Pearson correlation analysis of priming effect intension and soil basic index in 60 days |
3 讨论 3.1 不同碳负荷梯度下外源易利用态碳矿化特征
在稻田土壤中, 葡萄糖矿化产生CO2和13CO2量随着负荷的增加而增大(图 1), 这表明碳源供应增加能有效促进微生物活性.这与Blagodatskaya[23]等的结论:土壤溶液中的葡萄糖促进微生物从休眠期或潜在活跃期过渡到活跃阶段现象一致; 而这种激活进一步促进了胞外酶的产生, 从而加速了土壤有机质的分解和养分(主要是N、P和S)的释放.土壤中碳储存量通常是新鲜有机质输入量的线性函数[24, 25], 而新鲜有机质输入会影响土壤有机质矿化速率, 可以通过新鲜有机质的输入来增加或减少SOM的矿化速率[14].有研究显示Glu-13C累积矿化率符合一级动力学方程的变化规律(R2=0.99, P < 0.05), 通过调节土壤能源物质和微生物活性进而影响土壤碳库的易利用性, 这与土壤生态系统中有机碳的迁移、固持以及CO2、CH4的释放有密切联系, 成为土壤有机碳分解矿化的动力和土壤质量演变的关键指标[26].基于葡萄糖-C矿化率的一级双指数模型, 得到土壤中葡萄糖-C主要是以微生物易利用态碳快速转化的碳库(快库)形式存在, 其比例最后平稳在45%左右, 且平稳节点大致在碳负荷2×MBC, 这与激发效应在负荷3×MBC出现短暂的负激发效应和负荷为5×MBC后期出现持续的负激发效应结果一致.且两种碳库的分配比例均与易利用态碳添加量表现出显著的指数关系(R2=0.99, P < 0.05和R2=0.99, P < 0.05).负荷为3×MBC和5×MBC都具有最高且相近(55%左右)的生物可利用碳库, 并且负荷为0.5×MBC, 1×MBC具有最低且相近的碳滞留时间(4 d), 这表明在3×MBC已经满足了微生物的能量需求, 更多的葡萄糖以稳定碳储存; 而负荷5×MBC同时具有最高生物可利用碳库以及最长的碳滞留时间, 这表明负荷5×MBC的处理完全超过了微生物的碳需求量, 这也与Glu-C矿化率的结果一致[27, 28].在这些研究中, 初始时葡萄糖快速分解是由于在添加的底物中添加了易利用态的化合物.在加入的底物中消耗掉易利用的碳后, 添加的基质与天然矿物稳定的SOC(如角质和木质素)一起被利用[29].本研究显示, 起初CO2和13CO2矿化矿化速率加快, 后期矿化量逐渐平缓, 这种转变可能涉及物种优势的改变, 即在分解的早期阶段, 迅速繁殖的细菌使用更多的易利用态化合物, 而增长缓慢的真菌分解较难降解的有机质[29, 30].
3.2 不同碳负荷梯度对激发效应的影响微生物活性是土壤有机质(SOM)分解的重要因素, 由于微生物生长及其活性很大程度上取决于底物的可利用程度, 这些底物间接地影响了SOM分解[7].而SOM分解的加速或延缓主要取决于土壤的营养状况和微生物在增长阶段时土壤中的碳氮比[17, 31].在此研究中, 累积激发效应在实验初期随着负荷增加而增强, 这可能是前期微生物代谢生长而产生的表观激发效应, 这与Blagodatskaya等[12]提出表观激发效应理论一致, 即随着微生物代谢的激活和微生物生物量的增加, 二氧化碳的加速演化与SOM的代谢无关, 主要是通过微量的底物加速内部微生物的新陈代谢, 并立即(几分钟至几小时)增加呼吸活动; 但负荷为3×MBC在实验中期出现了短暂的负激发效应, 负荷为5×MBC的处理在后期持续的微弱负激发效应.在负荷为3×MBC处理中可能是与微生物群落结构的变化有关, 土壤有机碳主要用于微生物生长[17]; 而负荷为5×MBC的处理主要是由于添加的易利用态碳满足了微生物生长以及繁殖的需求而出现持续的负激发效应[31]. 1×MBC处理的碳累积激发是最高的, 并且保持持续的正激发效应, 可能在实验初期激发效应随易利用态碳量增加而增加, 而后期由于有机底物的输入促进了微生物活性, 使胞外酶分泌增加, 引起SOC持续的共代谢[32].因此, 负荷1×MBC的处理可能刚好满足了微生物的呼吸利用, 但生长繁殖的要求还没有达到, 因此需要分泌胞外酶分解SOM来维持生长繁殖.综上也证明对于不同梯度碳负荷和激发效应与微生物碳量有一个碳需求饱和关系.此外, 本实验初期累积激发效应与DOC显著正相关, 也证明了表观激发效应的理论[12], 以及β-葡糖苷酶和几丁质酶与矿化率呈显著负相关说明初期胞外酶对碳源的矿化没有影响; 而铵态氮和MBN与累积激发效应呈负相关关系, 这可能是由于实验处理添加的外源碳而导致的氮限制, 当添加过量外源碳而造成了氮限制时, 激发效应不明显[17], 这也与碳负荷为5×MBC后期出现的微弱的激发现象表现一致. β-葡糖苷酶和几丁质酶在微生物碳源获取中有重要意义, 其活性受碳源有效性的影响[33, 34].在激发效应的后期, β-葡糖苷酶与累积激发效应的负相关性增强, 而β-葡糖苷酶通过将纤维二糖转化为葡萄糖完成纤维素水解的最后一步, 为中枢代谢提供葡萄糖[35]; 而几丁质酶与其的负相关性变弱, 几丁质酶的主要作用是分解代谢和几丁质作为碳和能源的利用.这也说明在低碳负荷处理中激发效应为正时主要分解土壤中较难分解的有机质; 同时, 这两种酶活性的变化可能与矿化培养时期微生物利用策略的转换相关.
4 结论(1) 土壤有机碳的矿化随着葡萄糖-C负荷的增加而增加, 主要表现为在前期快速矿化而后逐渐达到稳定.碳负荷的增加显著增加了葡萄糖-C向土壤快速转化碳库中的分配, 促进土壤碳的累积矿化率.
(2) 易利用态碳源的输入促进了土壤有机碳的矿化, 表现为持续的正激发效应; 而且正激发效应随碳负荷的增加而逐渐增加.
(3) 不同碳负荷条件下, 矿化前期的累积激发效应主要受土壤微生物生物量(MBC)的影响, 而在后期主要受土壤酶活性(β-葡糖苷酶、几丁质酶)的影响.
| [1] |
冯嘉仪, 储双双, 王婧, 等. 华南地区几种典型人工林土壤有机碳密度及其与土壤物理性质的关系[J]. 华南农业大学学报, 2018, 39(1): 83-90. Feng J Y, Chu S S, Wang J, et al. Soil organic carbon density and its relationship with soil physical properties of typical plantations in South China[J]. Journal of South China Agricultural University, 2018, 39(1): 83-90. |
| [2] | Angst G, Mueller K E, Kögel-Knabner I, et al. Aggregation controls the stability of lignin and lipids in clay-sized particulate and mineral associated organic matter[J]. Biogeochemistry, 2017, 132(3): 307-324. DOI:10.1007/s10533-017-0304-2 |
| [3] | Pump J, Conrad R. Rice biomass production and carbon cycling in 13CO2 pulse-labeled microcosms with different soils under submerged conditions[J]. Plant and soil, 2014, 384(1-2): 213-229. DOI:10.1007/s11104-014-2201-y |
| [4] |
叶功富, 尤龙辉, 卢昌义, 等. 全球气候变化及森林生态系统的适应性管理[J]. 世界林业研究, 2015, 28(1): 1-6. Ye G F, You L H, Lu C Y, et al. Global climate change and adaptive management of forest ecosystem[J]. World Forestry Research, 2015, 28(1): 1-6. |
| [5] |
徐嘉晖, 孙颖, 高雷, 等. 土壤有机碳稳定性影响因素的研究进展[J]. 中国生态农业学报, 2018, 26(2): 222-230. Xu J H, Sun Y, Gao L, et al. A review of the factors influencing soil organic carbon stability[J]. Chinese Journal of Eco-Agriculture, 2018, 26(2): 222-230. |
| [6] | Wang H, Boutton T W, Xu W H, et al. Quality of fresh organic matter affects priming of soil organic matter and substrate utilization patterns of microbes[J]. Scientific Reports, 2015, 5: 10102. DOI:10.1038/srep10102 |
| [7] | Blagodatsky S, Blagodatskaya E, Yuyukina T, et al. Model of apparent and real priming effects:linking microbial activity with soil organic matter decomposition[J]. Soil Biology and Biochemistry, 2010, 42(8): 1275-1283. DOI:10.1016/j.soilbio.2010.04.005 |
| [8] | Kuzyakov Y, Friedel J K, Stahr K. Review of mechanisms and quantification of priming effects[J]. Soil Biology and Biochemistry, 2000, 32(11-12): 1485-1498. DOI:10.1016/S0038-0717(00)00084-5 |
| [9] |
戚瑞敏, 赵秉强, 李娟, 等. 添加牛粪对长期不同施肥潮土有机碳矿化的影响及激发效应[J]. 农业工程学报, 2016, 32(S2): 118-127. Qi R M, Zhao B Q, Li J, et al. Effects of cattle manure addition on soil organic carbon mineralization and priming effects under long-term fertilization regimes[J]. Transactions of the Chinese Society of Agricultural Engineering, 2016, 32(S2): 118-127. |
| [10] |
葛晓改, 周本智, 肖文发, 等. 生物质炭输入对土壤碳排放的激发效应研究进展[J]. 生态环境学报, 2016, 25(2): 339-345. Ge X G, Zhou B Z, Xiao W F, et al. Priming effect of biochar addition on soil carbon emission:a review[J]. Ecology and Environmental Sciences, 2016, 25(2): 339-345. |
| [11] |
苗淑杰, 乔云发, 张福韬. 黑土团聚体碳矿化和小麦秸秆对其的激发效应[J]. 水土保持学报, 2014, 28(4): 218-221. Miao S J, Qiao Y F, Zhang F T. Priming effect of wheat straw addition on organic carbon mineralization in various aggregates of black soil[J]. Journal of Soil and Water Conservation, 2014, 28(4): 218-221. |
| [12] | Blagodatskaya E, Kuzyakov Y. Mechanisms of real and apparent priming effects and their dependence on soil microbial biomass and community structure:critical review[J]. Biology and Fertility of Soils, 2008, 45(2): 115-131. DOI:10.1007/s00374-008-0334-y |
| [13] |
程璞, 张慧, 陈健. 进水碳负荷浓度对垂直潜流式人工湿地中植物根系微生物动态的影响[J]. 环境工程学报, 2014, 8(5): 2006-2012. Cheng P, Zhang H, Chen J. Effects of influent carbon loading on rhizosphere microbial diversity in vertical subsurface-flow constructed wetland[J]. Chinese Journal of Environmental Engineering, 2014, 8(5): 2006-2012. |
| [14] | Guenet B, Neill C, Bardoux G, et al. Is there a linear relationship between priming effect intensity and the amount of organic matter input?[J]. Applied Soil Ecology, 2010, 46(3): 436-442. DOI:10.1016/j.apsoil.2010.09.006 |
| [15] | Hütsch B W, Augustin J, Merbach W. Plant rhizodeposition-an important source for carbon turnover in soils[J]. Journal of Plant Nutrition and Soil Science, 2002, 165(4): 397-407. DOI:10.1002/1522-2624(200208)165:4<397::AID-JPLN397>3.0.CO;2-C |
| [16] | Yevdokimov I, Ruser R, Buegger F, et al. Microbial immobilisation of 13C rhizodeposits in rhizosphere and root-free soil under continuous 13C labelling of oats[J]. Soil Biology and Biochemistry, 2006, 38(6): 1202-1211. DOI:10.1016/j.soilbio.2005.10.004 |
| [17] | Blagodatskaya E V, Blagodatsky S A, Anderson T H, et al. Priming effects in Chernozem induced by glucose and N in relation to microbial growth strategies[J]. Applied Soil Ecology, 2007, 37(1-2): 95-105. DOI:10.1016/j.apsoil.2007.05.002 |
| [18] | 鲍士旦. 土壤农化分析[M]. (第三版). 北京: 中国农业出版社, 2000. |
| [19] | Loeppmann S, Blagodatskaya E, Pausch J, et al. Substrate quality affects kinetics and catalytic efficiency of exo-enzymes in rhizosphere and detritusphere[J]. Soil Biology and Biochemistry, 2016, 92: 111-118. DOI:10.1016/j.soilbio.2015.09.020 |
| [20] | Tian Q X, Yang X L, Wang X G, et al. Microbial community mediated response of organic carbon mineralization to labile carbon and nitrogen addition in topsoil and subsoil[J]. Biogeochemistry, 2016, 128(1-2): 125-139. DOI:10.1007/s10533-016-0198-4 |
| [21] | Chen R R, Senbayram M, Blagodatsky S, et al. Soil C and N availability determine the priming effect:microbial N mining and stoichiometric decomposition theories[J]. Global Change Biology, 2014, 20(7): 2356-2367. DOI:10.1111/gcb.2014.20.issue-7 |
| [22] | Sinsabaugh R L, Manzoni S, Moorhead D L, et al. Carbon use efficiency of microbial communities:stoichiometry, methodology and modelling[J]. Ecology Letters, 2013, 16(7): 930-939. DOI:10.1111/ele.12113 |
| [23] | Blagodatskaya E, Kuzyakov Y. Active microorganisms in soil:critical review of estimation criteria and approaches[J]. Soil Biology and Biochemistry, 2013, 67: 192-211. DOI:10.1016/j.soilbio.2013.08.024 |
| [24] | Petersen B M, Jensen L S, Hansen S, et al. CN-SIM:a model for the turnover of soil organic matter. Ⅱ. Short-term carbon and nitrogen development[J]. Soil Biology and Biochemistry, 2005, 37(2): 375-393. DOI:10.1016/j.soilbio.2004.08.007 |
| [25] | Grace P R, Ladd J N, Robertson G P, et al. SOCRATES-a simple model for predicting long-term changes in soil organic carbon in terrestrial ecosystems[J]. Soil Biology and Biochemistry, 2006, 38(5): 1172-1176. DOI:10.1016/j.soilbio.2005.09.013 |
| [26] | Dharmakeerthi R S, Hanley K, Whitman T, et al. Organic carbon dynamics in soils with pyrogenic organic matter that received plant residue additions over seven years[J]. Soil Biology and Biochemistry, 2015, 88: 268-274. DOI:10.1016/j.soilbio.2015.06.003 |
| [27] | Lu Y H, Watanabe A, Kimura M. Contribution of plant-derived carbon to soil microbial biomass dynamics in a paddy rice microcosm[J]. Biology and Fertility of Soils, 2002, 36(2): 136-142. DOI:10.1007/s00374-002-0504-2 |
| [28] | Gunina A, Kuzyakov Y. Sugars in soil and sweets for microorganisms:review of origin, content, composition and fate[J]. Soil Biology and Biochemistry, 2015, 90: 87-100. DOI:10.1016/j.soilbio.2015.07.021 |
| [29] | Baumann K, Marschner P, Smernik R J, et al. Residue chemistry and microbial community structure during decomposition of eucalypt, wheat and vetch residues[J]. Soil Biology and Biochemistry, 2009, 41(9): 1966-1975. DOI:10.1016/j.soilbio.2009.06.022 |
| [30] | Brant J B, Sulzman E W, Myrold D D. Microbial community utilization of added carbon substrates in response to long-term carbon input manipulation[J]. Soil Biology and Biochemistry, 2006, 38(8): 2219-2232. DOI:10.1016/j.soilbio.2006.01.022 |
| [31] | Kuzyakov Y. Review:factors affecting rhizosphere priming effects[J]. Journal of Plant Nutrition and Soil Science, 2002, 165(4): 382-396. DOI:10.1002/1522-2624(200208)165:4<382::AID-JPLN382>3.0.CO;2-# |
| [32] | Zhu Z K, Ge T D, Luo Y, et al. Microbial stoichiometric flexibility regulates rice straw mineralization and its priming effect in paddy soil[J]. Soil Biology and Biochemistry, 2018, 121: 67-76. DOI:10.1016/j.soilbio.2018.03.003 |
| [33] |
李猛, 张恩平, 张淑红, 等. 长期不同施肥设施菜地土壤酶活性与微生物碳源利用特征比较[J]. 植物营养与肥料学报, 2017, 23(1): 44-53. Li M, Zhang E P, Zhang S H, et al. Comparison of soil enzyme activities and microbial C metabolism in installed vegetable fields under long-term different fertilization[J]. Journal of Plant Nutrition and Fertilizer, 2017, 23(1): 44-53. |
| [34] | Ali R S, Kandeler E, Marhan S, et al. Controls on microbially regulated soil organic carbon decomposition at the regional scale[J]. Soil Biology and Biochemistry, 2018, 118: 59-68. DOI:10.1016/j.soilbio.2017.12.007 |
| [35] |
王霏, 王绍琛, 曹明明, 等. 土壤微生物中新型β-葡萄糖苷酶的挖掘与鉴定[J]. 微生物学通报, 2018, 45(1): 71-80. Wang F, Wang S C, Cao M M, et al. A novel β-glucosidase from soil microbes[J]. Microbiology China, 2018, 45(1): 71-80. |