 2019, Vol. 40
2019, Vol. 40

2. 江西理工大学资源与环境工程学院, 赣州 341000;
3. 湖南省宁乡市农业技术推广中心, 宁乡 410600;
4. 湖南省农业科学院土壤肥料研究所, 长沙 410125
 , CHENG Ai-wu3 , TANG Hai-ming4 , GE Ti-da1 , DENG Yang-wu2 , SU Yi-rong1 , CHEN Xiang-bi1
, CHENG Ai-wu3 , TANG Hai-ming4 , GE Ti-da1 , DENG Yang-wu2 , SU Yi-rong1 , CHEN Xiang-bi1
2. School of Resource and Environmental Engineering, Jiangxi University of Science and Technology, Ganzhou 341000, China;
3. Agricultural Technology Promotion Center, Ningxiang 410600, China;
4. Hunan Soil and Fertilizer Institute, Hunam Province Institute of Agricultural Science, Changsha 410125, China
土壤微生物在陆地生态系统碳循环过程中扮演着两个完全不同的角色:分泌胞外酶分解植物残体类有机质, 促进土壤碳向大气释放(分解代谢); 通过有机底物被微生物摄取直至死亡, 将有机碳同化并储存为难以降解的微生物源有机形态(合成代谢), 从而促进土壤碳固定[1].不同生态系统中微生物执行这两个相对独立过程的强弱程度是土壤有机碳周转的重要控制阀, 也是决定土壤碳“源”和“汇”特征的重要因素.
微生物分配给自身生长的碳量占微生物摄取总碳的比例, 即微生物碳源利用效率(carbon use efficiency, CUE), 是权衡微生物合成代谢和分解代谢强弱的重要指标[2, 3].国内外主要通过向土壤中添加13C或14C标记的某一有机底物, 如葡萄糖、秸秆等, 通过测定微生物生物量碳(MBC)和呼吸释放CO2中的13C丰度或14C放射性强度来计算微生物CUE[4].然而, 该方法得到的CUE与其实际定义不符而备受争议[5].首先, 由于微生物对不同有机底物的利用能力和代谢途径不同, 造成基于不同有机底物测定的土壤微生物CUE差异大[4].其次, 该方法只考虑了利用同位素标记碳源的微生物, 而不是所有微生物群落.再者, 添加外源有机底物改变了土壤环境, 进而干扰微生物生长和呼吸.近年来, 研究学者建立了一种基于监测微生物18O-DNA的CUE测定方法, 在一定程度上克服了以上缺陷[6].该方法通过标记土壤中水为一定丰度的H218O, 微生物生长过程中摄取H218O, 使其部分DNA被标记为18O-DNA, 基于土壤中总MBC与总DNA含量的线性关系, 计算微生物生长速率(CGrowth), 再结合土壤基础呼吸计算微生物CUE[5].
微生物群落功能多样性可以反映微生物分解代谢活动, 且土壤微生物碳源利用效率受微生物对底物碳源代谢能力的影响[7].目前, 土壤微生物对碳源代谢能力的研究方法主要有Biolog、多重底物诱导呼吸和MicroRespTM等方法. MicroRespTM方法克服了Biolog微平板法依赖土壤悬浮液提取物和细胞后续生长状况、多重底物诱导呼吸自动化程度低、操作繁琐等缺点, 且能测定全土/鲜土的微生物代谢情况, 并具有操作简单、快速等优点[8].近年来, MicroRespTM方法已应用于森林、草地和农田等旱地土壤[9~11]以及含水量较高的水稻土[12].
我国现有稻田总面积2 300万hm2, 占全国耕地总面积约25%, 且主要分布于亚热带地区, 尤以长江中下游地区(湘、鄂、赣、苏、浙、皖、沪)最为集中.许多研究已经表明, 我国水稻土的固碳效应明显, 固碳潜力较大[13], 其中以南方双季稻种植区水稻土的固碳能力最强[14].水稻土中微生物量远远高于相邻旱地土壤[15, 16], 这暗示了微生物对水稻土碳固持可能有重要贡献.长期不同施肥处理下水稻土的固碳能力不同, 如Tong等[17]的研究发现:与不施肥相比, 化肥处理有机碳含量没有显著差异, 施加有机肥显著提高土壤有机碳含量.董春华等[18]的研究也发现施用有机肥和有机无机肥配施比单施无机肥能更快速地提高土壤有机碳含量.然而, 土壤微生物合成代谢和分解代谢对不同施肥有何响应尚不明确, 这对解析不同施肥处理下水稻土固碳差异的微生物机制有重要意义.
本研究基于亚热带红壤丘陵区典型稻田长期定位施肥试验, 采集不同施肥处理下水稻分蘖旺期根际土和非根际土, 研究根际和非根际土壤微生物CUE对施肥的响应特征, 并基于MicroRespTM方法分析土壤微生物对不同碳源的代谢特征, 其结果有助于揭示不同施肥制度下水稻土微生物碳固持的机制, 以期为完善水稻土碳固持机制提供重要理论依据.
1 材料与方法 1.1 试验区概况长期定位施肥试验点位于湖南省宁乡县农技中心(112°18′ E, 28°07′ N), 试验始于1986年.该区域为典型的水稻两熟农作物主产区, 试验点属亚热带季风湿润气候, 全年气候温暖湿润, 多年平均气温16.8℃, 年均降雨量1 553.7 mm, 年均蒸发量1 353.9 mm, 海拔36.1 m, 无霜期274 d.试验地土壤为水稻土, 河沙泥土种, 1986年~1992年作物种植制度为稻-玉米, 1993年至今为稻-稻-冬小麦[19].土壤肥力中等, 排灌条件良好.定位试验开始前0~20 cm耕层土壤的基础肥力为:有机质29.4 g·kg-1、全氮2.0 g·kg-1、全磷0.6 g·kg-1、全钾20.6 g·kg-1、碱解氮144.1 mg·kg-1、有效磷12.9 mg·kg-1、速效钾33.0 mg·kg-1和pH 6.85[20].
1.2 试验设计与土壤样品采集选择5个施肥处理. ①无肥对照:不施加任何肥料(CK)、②全化肥:施用氮、磷、钾化肥(NPK)、③秸秆还田+化肥:晚稻秸秆还田和化肥(NPKS)、④30%有机肥:30%氮素来源于有机肥(猪粪), 70%氮素来自化肥氮(LOM)、⑤60%有机肥:60%氮素来源于有机肥, 40%氮素来源于化肥氮(HOM).每个试验处理小区长10.00 m, 宽6.67 m, 总面积达66.7 m2, 各个试验小区用水泥梗分隔开, 向地下延伸100 cm, 高出试验田水平面35 cm, 以免各试验处理小区窜灌、窜排.
NPK处理在早稻季和晚稻季总施N分别为142.5 kg·hm-2和157.5 kg·hm-2; 施P2O5分别为54.0 kg·hm-2和43.2 kg·hm-2; 施K2O分别为63.0 kg·hm-2和81.0 kg·hm-2. NPKS处理的秸秆还田量在早稻季和晚稻季分别为2 775.0 kg·hm-2(秸秆中N、P2O5、K2O的含量分别为0.65%、0.13%、0.89%)和3 600.0 kg·hm-2(秸秆中N、P2O5、K2O的含量分别为0.68%、0.15%、0.91%). LOM和HOM处理的有机肥均为腐熟猪粪(有机肥的N、P2O5和K2O养分含量分别为1.77%、0.83%和1.12%), 其中LOM处理在早稻季和晚稻季的猪粪施肥量分别为2 625.0 kg·hm-2和2 670.0 kg·hm-2; HOM处理在早稻季和晚稻季的猪粪施肥量分别为5 250.0 kg·hm-2和5 340.0 kg·hm-2.各个处理以等氮量为基准, 不足的氮、磷、钾肥用化肥补足; 早稻季和晚稻季各施肥处理的P2O5、秸秆和有机肥均于稻田耕地时作基肥一次性施入; N和K2O作基肥和追肥2次施入, 基、追肥比例均按7 :3施用; 基肥在耕地时施入, 追肥在移栽后7 d施用, 各施肥处理每年所使用的肥料种类及施用量均保持一致.早稻和晚稻田间管理为在移栽后保持浅水条件, 结合施用分蘖肥进行杂草防除, 分蘖期进行晒田, 晒田后复水, 后期进行干湿交替灌溉, 水稻完熟后落干晒田、收获, 其他管理措施同常规大田生产[20].
于2017年9月5日晚稻分蘖旺期, 采集不同施肥处理的根际和非根际土壤样品, 每个处理的小区中随机采集长势具有代表性的水稻四兜, 代表四个重复样品.具体操作为:在水平方向距离水稻茎秆5 cm处采用铁锹挖出整株水稻(深度0~15 cm), 装入干净塑料袋, 带回实验室后采用镊子将水稻根上附着的土壤轻轻剥离作为根际土[21], 另采用铁锹取出水平方向距离水稻茎秆5~10 cm的表层土(深度0~15 cm)作为非根际土.将土壤样品中肉眼可见的植物残体、根系、石粒等拣出, 室温自然风干、过100目筛后备用.
1.3 样品分析 1.3.1 微生物碳源利用效率的测定将300 mg(以鲜土计)25℃预先培养两周的土壤样品称量到2 mL螺帽冻存管中, 每个样品2次重复.用移液枪将18O-H2O(97.0 at%, 上海化工研究院有限公司)注入到冻存管中, 使最终土壤水中的18O丰度达到20%.设置加入相同体积未标记的水作为自然丰度对照.将2 mL冻存管中的土壤样品在涡旋仪上轻轻混匀, 然后打开螺帽, 并将其立即转移到50 mL培养瓶中, 密封培养瓶.同时, 设置加入5 g土壤样品至5 mL离心管, 并转移至50 mL培养瓶, 密闭培养, 用于采集气体.每间隔4个培养瓶设置一个空白对照.在25℃恒温培养箱中培养24 h后, 取出2 mL冻存管、盖上螺帽、放入-80℃冰箱保存, 直至微生物DNA提取[22].同时, 采用三通阀注射器从5 g土壤样品的培养瓶中抽取20 mL气体, 注射至12 mL真空顶空瓶, 并采集空白对照样品.
采用Agilent 7890A气相色谱仪测定采集气体样品中的CO2含量, 并计算微生物通过呼吸作用所产生的基础呼吸碳量(CRespiration).
采用文献[5]中的方法测定土壤微生物CUE.其具体步骤为:采用土壤DNA提取试剂盒(Fast DNA SPIN Kit for Soil)提取土壤总DNA; 使用Nanodrop one超微量紫外分光光度计测定DNA质量浓度, 剩余DNA样品在60℃干燥过夜以去除水; 采用MAT 253型稳定性同位素比率质谱仪(美国热电公司)测定18O丰度.根据本研究40个土壤样本中的微生物总DNA含量与MBC含量, 建立二者的线性关系(y=4.85x+16.3267, R2=0.605, 数据未列出, 式中x为DNA含量, y为MBC含量).再根据该线性方程以及18O-DNA丰度, 获得该段时间微生物利用碳源用于自身生长的量(CGrowth), 以下公式计算微生物CUE:
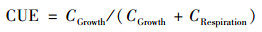
|
采用MicroRespTM方法[8], 测定土壤微生物对31种碳源的代谢能力, 并以无菌水为空白对照.所选的31种碳源包括14种羧酸类(甲酸、L-抗坏血酸、α-酮丁酸、富马酸、α-酮戊二酸、丁香酸、L-酒石酸、甘醇酸、琥珀酸、DL-苹果酸、柠檬酸、奎尼酸、D-半乳糖醛酸和DL-乳酸), 6种氨基酸(L-丙氨酸、L-天冬酰胺、L-蛋氨酸、L-缬氨酸、L-苏氨酸和L-谷氨酸), 7种复杂化合物(硫胺素、咪唑、尿刊酸、肉桂、香豆素、儿茶素和槲皮素), 4种碳水化合物(糊精、D-甘露糖醇、L-鼠李糖和葡萄糖).碳源配制参考文献[23]中的方法, 具体为:称取难溶性碳源(香豆素、丁香酸、DL-儿茶素、尿刊酸、琥珀酸、肉桂、槲皮素、硫胺素和L-谷氨酸)0.69 g、可溶性碳源(其余22种)2.75 g, 分别定容至50 mL, 并调pH至5.5~6.0, 存于-20℃冰箱备用.
检测板制备步骤[8]:将7.5 mg甲酚红、5.366 4 g氯化钾、0.100 8 g碳酸氢钠溶于水中, 定容至200 mL, 即为甲酚红溶液; 配制3%的琼脂溶液100 mL, 121℃高压灭菌20 min使之溶解; 将甲酚红溶液与3%的琼脂溶液按体积比(1 :2)混合并搅拌至少1.5 h, 置于60℃水浴锅中, 一直搅拌防止凝固; 用八孔电动连续加样器向96孔透明酶标板中每孔加入混合液150 μL, 待检测板冷却后用封口膜封好, 置于干燥器中备用.
微生物碳源代谢测定:称取50 g经预培养的土壤样品于乐扣盒中, 加水20 g, 转速150 r·min-1振荡30 min使之成为匀浆, 用电动移液器吸取350 μL匀浆至预先加入30 μL碳源的96孔深孔板中, 将密封垫片压于深孔板上, 再将检测板扣于垫片上, 保证其密闭性.用铁板将深孔板和检测板固定, 置于25℃水平振荡恒温培养箱(减少水稻土上覆水对CO2的溶解)中培养4 h, 取出显色板采用多功能酶标仪在590 nm波长下测定吸光值.基于CO2产生率和指示剂对特定波长光吸收值变化的校准曲线拟合为指数衰减模型, 计算4 h内土壤微生物对不同碳源的代谢能力(CO2产生率).指数衰减模型为:
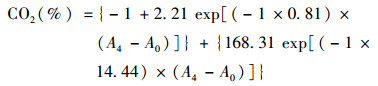
|
式中, A4为添加某一碳源并培养4 h后检测板的吸光值; A0为培养前未添加碳源的检测板吸光值.
CO2产生速率[μg·(g·h)-1]计算公式:
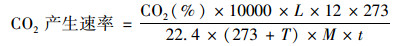
|
式中, T为培养温度(25℃), L为深孔板每孔体积(945 μL), M为干土重(g), t为培养时间(4 h).
1.3.3 基本理化性质的测定用风干法测定土壤含水率(water content, WC), 采用pH计测定土壤pH(水土比为2.5 :1), 且供试土壤样品的pH<7.0, 可以把土壤全碳视作有机碳.采用碳氮元素分析仪(VARIO MAX C/N, Germany)测定(干烧法)土壤有机碳(soil organic carbon, SOC)和全氮(soil total nitrogen, TN); 土壤铵态氮和硝态氮含量用0.5 mol·L-1K2SO4溶液浸提, 采用流动注射仪(Fiastar 5000, 瑞典福斯)测定; 土壤微生物量碳氮(MBC, MBN)采用氯仿熏蒸提取, MBC采用有机碳分析仪(岛津Vwp, 日本)测定[24]; MBN采用流动注射仪(Fiastar 5000, 瑞典福斯)测定[25].
1.4 数据统计分析采用SPSS 19.0软件进行差异显著性和Pearson相关分析.同一施肥处理下根际土与非根际土之间采用配对T检验、不同施肥处理间采用One-way ANOVA(单因素方差分析)进行差异显著性检验; 采用Canoco 5.0对土壤微生物碳源利用与环境因子进行RDA分析.采用Origin 8.5软件进行数据处理及绘图.
2 结果与分析 2.1 微生物碳源利用效率及其影响因素不同施肥处理下微生物量碳含量范围为817.0~1 807.4 mg·kg-1, 且在根际土和非根际土中以HOM处理最高, CK处理最低(图 1).除NPKS处理的MBC含量根际土显著高于非根际土(P < 0.01)外, 其余处理在根际与非根际土之间无显著差异.

|
不同小写字母表示根际或者非根际土在不同施肥处理下该指标在(P < 0.05)水平下差异显著, *和* *分别表示在同一施肥处理下的根际与非根际土之间该指标在(P < 0.05)和(P < 0.01)水平下差异显著 图 1 不同施肥处理根际和非根际土壤微生物碳分配 Fig. 1 Soil microbial distribution in rhizosphere and non-rhizosphere soil of different fertilization treatments |
土壤微生物基础呼吸速率(以CO2-C计)范围为251.4~418.1 ng·(g·h)-1, 且该速率在根际土中以HOM处理最高, CK和NPK处理最低, 在非根际土中不同处理无显著差异(P>0.05).总体上, 根际土的微生物基础呼吸速率高于非根际土, 且在NPKS、LOM和HOM处理下差异显著(P < 0.01).
基于该方程和18O-DNA丰度计算微生物生长速率CGrowth.结果表明:不同施肥处理下土壤微生物生长速率(CGrowth)的范围为216.0~231.7 ng·(g·h)-1; 总体上, 该指标以非根际土高于根际土, 且在LOM和HOM处理下差异达极显著水平(P < 0.01);不同施肥处理下以HOM和LOM高于其它处理.
不同施肥处理下土壤微生物碳源利用效率(CUE)的变化范围为0.38~0.47.总体上, CUE以非根际土高于根际土, 且在NPKS、LOM和HOM处理下差异显著(P < 0.01).不同施肥处理下非根际土CUE无显著差异, 根际土中以NPK最高, LOM和HOM最低.
2.2 微生物对不同类型外源碳源的代谢特征总体上, 各施肥处理下土壤微生物对外源碳源的代谢能力(分解为CO2)以非根际土高于根际土(图 2).与CK和NPK相比, 施用有机物料(秸秆还田和有机肥)均提高根际(氨基酸类除外)与非根际土壤中微生物对外源碳源的利用, 但根际土中微生物对复杂化合物的利用在不同施肥处理下无显著差异.各处理土壤微生物对羧酸类的平均利用速率最高, 对复杂化合物的平均利用速率最低.冗余分析(RDA)表明:环境因子对土壤微生物碳源利用情况的第一轴解释了50.0%的变异, 第二轴解释了6.5%的变异(图 3).微生物对外源碳源利用情况的聚类总体以根际土与非根际土分开; 不同施肥处理下, CK与施肥处理分开, 且NPK与NPKS相对聚集, LOM与HOM相对聚集, NPK、NPKS与LOM、HOM分开, 表明不同施肥处理显著改变土壤微生物对外源碳源的利用特征.蒙特卡罗检验表明, 影响土壤微生物对外源碳源利用能力的因子主要为C/N、MBC、NO3--N、NH4+-N、TN和pH(P < 0.05).

|
不同小写字母表示根际或者非根际土在不同施肥处理下该指标在(P < 0.05)水平下差异显著, 根际土中的*表示在同一施肥处理下的根际与非根际土之间该指标在(P < 0.05)水平下差异显著 图 2 不同施肥处理下根际和非根际土壤微生物对不同类型外源碳源的代谢特征 Fig. 2 Characteristics of utilization of different types of exogenous carbon sources by rhizosphere and non-rhizosphere soil microorganisms under different fertilization treatments |

|
实心图标代表根际土, 空心图标代表非根际土 图 3 微生物碳源利用速率与环境因子的RDA分析 Fig. 3 RDA analysis of microbial carbon source utilization rate and environmental factors |
本研究中, 施用有机物料(秸秆和有机肥)显著增加水稻根际和非根际土中总MBC和CGrowth.与不施肥相比, 施用有机物料提高了土壤速效养分, 为微生物生长和繁殖提供了碳底物和养分, 进而提高土壤微生物生长速率和微生物量[26].有机肥相比秸秆还田的效果更好, 其原因可能是秸秆还田后土壤C/N升高, 且多是缓效养分, 微生物生长受到一定的限制[27].不同施肥处理下非根际土基础呼吸无显著差异, 但施用有机物料显著增加根际土基础呼吸.这可能因为易分解态有机碳是微生物利用的主要碳源, 而不同施肥处理下非根际土中易分解态有机碳的含量无显著差异[28].在根际土中, 水稻根系分泌物的输入会引起土壤有机碳分解的正激发效应, 进而增加微生物呼吸[29].水稻根系分泌物的数量受施肥影响, 如施用有机物料增加水稻根系生物量, 进而增加根系分泌物[30].与CK相比, NPK处理的CGrowth和基础呼吸均无显著差异, 其原因是本研究中土壤的有机碳含量无显著差异[17], 可能造成微生物可利用的碳源与CK处理无显著差异.
本研究水稻土中微生物CUE的范围为0.38~0.47, 略低于土壤生态系统平均值(0.55).这可能是由于以往研究多采用13C底物标记法, 其高估了土壤微生物CUE[3, 31].不同施肥处理下非根际土CUE无显著差异, 其原因可能是非根际土的生理环境相对稳定, Dijkstra等[32]认为CUE的高低与生理环境的稳定有强烈的关系.根际土壤微生物CUE以NPK最高, LOM和HOM最低.有机物料输入提高了土壤环境中的C :养分比值, 在这种营养限制的条件下微生物通过溢流呼吸(overflow respiration)释放更多CO2, 使C :养分的比例更有利于满足自身对营养的需求[33], 降低微生物CUE[5]. NPK处理的CUE最高, 可能是由于NPK的输入, 参与N的芳香化合物降解的氧化酶受到抑制, 土壤微生物对能量的需求降低[5].总体上, 根际土CUE低于非根际土, 一方面由于本试验CRespiration的值相对要高一些, 微生物分配的呼吸碳高于生长碳; 另一方面可能是植物生长过程中根系吸收养分, 进而增加了根际土中C :养分, CUE随C :养分比的增加而降低[5].
3.2 不同施肥处理下微生物碳源代谢特征土壤微生物群落对碳源代谢特征可以反映生物有效性和土壤微生物功能多样性[23], 平均CO2产生率可作为微生物整体活性的有效指标, 也可反映微生物群落对碳源利用的能力[11].本研究中, 施用有机物料(秸秆和有机肥)均比不施肥(对照)和施用化肥显著增加土壤微生物对外源碳源(复杂化合物除外)的代谢能力.秸秆还田处理碳源代谢能力略低于施用有机肥处理, 其原因可能是秸秆还田后土壤C/N升高, 微生物分解慢, 微生物对碳源的代谢能力降低[27].有机肥本身含有大量活的微生物、活性有机碳源, 起到“接种”作用[34], 其施入土壤后促进微生物对碳源的代谢能力.本研究中, 土壤微生物对羧酸类的平均利用速率最高, 复杂化合物最低.羧酸类碳源仅占土壤溶解性有机碳的极少部分, 但它们却是土壤微生物生长和代谢的重要能源[35].复杂化合物的降解需要多种不同胞外酶的共同作用, 因此微生物生长过程中需将更多碳源和能量投入到胞外酶的合成, 进而降低微生物利用复杂化合物速率[36]. RDA分析表明, CK处理的土壤微生物对外源碳源的代谢能力与施肥处理分开, 说明施肥大大改变了土壤环境, 进而显著改变土壤微生物对外源碳源的利用特征. NPK与NPKS相对聚集、LOM与HOM相对聚集, 且NPK、NPKS与LOM、HOM分开, 这可能是因为NPK和NPKS处理通过水稻根系分泌物或秸秆还田引入了与稻田土壤原有有机质性质相同的外源有机物料, 而有机粪肥的施用可能引入了异源有机质和微生物, 进而显著影响土壤微生物对外源碳源的利用.
基于18O-H2O方法发现, 非根际土中微生物CUE和基础呼吸在不同施肥处理下无显著差异, 但有外源碳源输入(如根系分泌物)的情况下, 施用有机物料(秸秆或有机肥)处理增加基础呼吸、降低CUE.该结果也可以由基于MicroRespTM方法获得的碳代谢数据所证实, 即采用31种碳源模拟根系分泌物的各种有机物组分, 施用有机物料处理可显著增加微生物对外源碳的代谢能力(释放为CO2).此外, 水稻非根际土土壤微生物对外源碳源的代谢能力高于根际土(图 2), 这与基于18O-H2O方法的土壤基础呼吸结果(根际土高于非根际土)不一致. MicroRespTM方法主要监测微生物对外源碳的分解情况, 但无法区分激发效应, 18O-H2O主要监测微生物对土壤原有碳源的分解情况.根际土长期处于外源碳源输入(根系分泌物)的环境中, 对外源碳源输入有一定的适应能力, 而非根际土中外源碳源的输入会引起原有有机碳分解的正激发效应, 进而微生物对有机碳的矿化作用增强[29].
4 结论(1) 与不施肥和施用化肥相比, 施用有机物料(秸秆和有机肥)能显著增加土壤微生物量碳和微生物生长速率(CGrowth); 施肥不改变微生物基础呼吸和CUE(非根际土), 但在根际土中, 受根际分泌物的影响, 施用有机物料增加微生物基础呼吸、降低其CUE.
(2) 非根际土微生物对外源碳源的代谢能力高于根际土; 施用有机物料均能在一定程度上提高微生物对羧酸类、氨基酸、碳水化合物的利用速率; 单施化肥和秸秆还田+化肥处理的外源碳代谢特征相似, 其与施用高量或低量有机肥的碳源代谢特征有明显差别.
| [1] | Liang C, Schimel J P, Jastrow J D. The importance of anabolism in microbial control over soil carbon storage[J]. Nature Microbiology, 2017, 2(8): 17105. DOI:10.1038/nmicrobiol.2017.105 |
| [2] | Manzoni S, Taylor P, Richter A, et al. Environmental and stoichiometric controls on microbial carbon-use efficiency in soils[J]. New Phytologist, 2012, 196(1): 79-91. DOI:10.1111/j.1469-8137.2012.04225.x |
| [3] | Sinsabaugh R L, Manzoni S, Moorhead D L, et al. Carbon use efficiency of microbial communities:stoichiometry, methodology and modelling[J]. Ecology Letters, 2013, 16(7): 930-939. DOI:10.1111/ele.12113 |
| [4] | Frey S D, Lee J, Melillo J M, et al. The temperature response of soil microbial efficiency and its feedback to climate[J]. Nature Climate Change, 2013, 3(4): 395-398. DOI:10.1038/nclimate1796 |
| [5] | Spohn M, Pötsch E M, Eichorst S A, et al. Soil microbial carbon use efficiency and biomass turnover in a long-term fertilization experiment in a temperate grassland[J]. Soil Biology and Biochemistry, 2016, 97: 168-175. DOI:10.1016/j.soilbio.2016.03.008 |
| [6] | Schwartz E. Characterization of growing microorganisms in soil by stable isotope probing with H218O[J]. Applied and Environmental Microbiology, 2007, 73(8): 2541-2546. DOI:10.1128/AEM.02021-06 |
| [7] | Preston-Mafham J, Boddy L, Randerson P F. Analysis of microbial community functional diversity using sole-carbon-source utilisation profiles-a critique[J]. FEMS Microbiology Ecology, 2002, 42(1): 1-14. |
| [8] | Campbell C D, Chapman S J, Cameron C M, et al. A rapid microtiter plate method to measure carbon dioxide evolved from carbon substrate amendments so as to determine the physiological profiles of soil microbial communities by using whole soil[J]. Applied and Environmental Microbiology, 2003, 69(6): 3593-3599. DOI:10.1128/AEM.69.6.3593-3599.2003 |
| [9] |
鲁顺保, 郭晓敏, 芮亦超, 等. 澳大利亚亚热带不同森林土壤微生物群落对碳源的利用[J]. 生态学报, 2012, 32(9): 2819-2826. Lu S B, Guo X M, Rui Y C, et al. Utilization of carbon sources by the soil microbial communities of different forest types in subtropical Australia[J]. Acta Ecologica Sinica, 2012, 32(9): 2819-2826. |
| [10] | Macdonald C A, Thomas N, Robinson L, et al. Physiological, biochemical and molecular responses of the soil microbial community after afforestation of pastures with Pinus radiata[J]. Soil Biology and Biochemistry, 2009, 41(8): 1642-1651. DOI:10.1016/j.soilbio.2009.05.003 |
| [11] |
陈晓娟, 吴小红, 刘守龙, 等. 不同耕地利用方式下土壤微生物活性及群落结构特性分析:基于PLFA和MicroRespTM方法[J]. 环境科学, 2013, 34(6): 2375-2382. Chen X J, Wu X H, Liu S L, et al. Microbial activity and community structure analysis under the different land use patterns in farmland soils:based on the methods PLFA and microRespTM[J]. Environmental Science, 2013, 34(6): 2375-2382. |
| [12] |
王伟华, 刘毅, 唐海明, 等. 长期施肥对稻田土壤微生物量、群落结构和活性的影响[J]. 环境科学, 2018, 39(1): 430-437. Wang W H, Liu Y, Tang H M, et al. Effects of long-term fertilization regimes on microbial biomass, community structure and activity in a paddy soil[J]. Environmental Science, 2018, 39(1): 430-437. |
| [13] | 潘根兴, 李恋卿, 郑聚锋, 等. 土壤碳循环研究及中国稻田土壤固碳研究的进展与问题[J]. 土壤学报, 2008, 45(5): 901-914. DOI:10.3321/j.issn:0564-3929.2008.05.017 |
| [14] | Huang S, Sun Y N, Zhang W J. Changes in soil organic carbon stocks as affected by cropping systems and cropping duration in China's paddy fields:a meta-analysis[J]. Climatic Change, 2012, 112(3-4): 847-858. DOI:10.1007/s10584-011-0255-x |
| [15] | Li Y, Wu J S, Liu S L, et al. Is the C:N:P stoichiometry in soil and soil microbial biomass related to the landscape and land use in southern subtropical China?[J]. Global Biogeochemical Cycles, 2012, 26(4): GB4002. |
| [16] | Chen X B, Wang A H, Li Y, et al. Fate of 14C-labeled dissolved organic matter in paddy and upland soils in responding to moisture[J]. Science of the Total Environment, 2014, 488-489: 268-274. DOI:10.1016/j.scitotenv.2014.04.071 |
| [17] | Tong C L, Xiao H A, Tang G Y, et al. Long-term fertilizer effects on organic carbon and total nitrogen and coupling relationships of C and N in paddy soils in subtropical China[J]. Soil and Tillage Research, 2009, 106(1): 8-14. DOI:10.1016/j.still.2009.09.003 |
| [18] |
董春华, 高菊生, 曾希柏, 等. 长期有机无机肥配施下红壤性稻田水稻产量及土壤有机碳变化特征[J]. 植物营养与肥料学报, 2014, 20(2): 336-345. Dong C H, Gao J S, Zeng X B, et al. Effects of long-term organic manure and inorganic fertilizer combined application on rice yield and soil organic carbon content in reddish paddy fields[J]. Journal of Plant Nutrition and Fertilizer, 2014, 20(2): 336-345. |
| [19] | Hao X H, Liu S L, Wu J S, et al. Effect of long-term application of inorganic fertilizer and organic amendments on soil organic matter and microbial biomass in three subtropical paddy soils[J]. Nutrient Cycling in Agroecosystems, 2008, 81(1): 17-24. DOI:10.1007/s10705-007-9145-z |
| [20] |
唐海明, 肖小平, 汤文光, 等. 长期施肥对双季稻田甲烷排放和关键功能微生物的影响[J]. 生态学报, 2017, 37(22): 7668-7678. Tang H M, Xiao X P, Tang W G, et al. Effects of long-term fertilizer treatments on CH4 fluxes and key functional microorganisms in a double-cropping paddy field[J]. Acta Ecologica Sinica, 2017, 37(22): 7668-7678. |
| [21] |
孙小霞, 王海斌, 何海斌, 等. 田间旱育条件下不同化感潜力水稻根际土壤酚酸类和萜类物质分析[J]. 中国生态农业学报, 2014, 22(7): 806-812. Sun X X, Wang H B, He H B, et al. Analysis of phenolic acids and terpenoids in rhizosphere soils of different allelopathic rice varieties under dry field conditions[J]. Chinese Journal of Eco-Agriculture, 2014, 22(7): 806-812. |
| [22] | Spohn M, Klaus K, Wanek W, et al. Microbial carbon use efficiency and biomass turnover times depending on soil depth-implications for carbon cycling[J]. Soil Biology and Biochemistry, 2016, 96: 74-81. DOI:10.1016/j.soilbio.2016.01.016 |
| [23] | Banning N C, Lalor B M, Cookson W R, et al. Analysis of soil microbial community level physiological profiles in native and post-mining rehabilitation forest:which substrates discriminate?[J]. Applied Soil Ecology, 2012, 56: 27-34. DOI:10.1016/j.apsoil.2012.01.009 |
| [24] | Wu J, Joergensen R G, Pommerening B, et al. Measurement of soil microbial biomass C by fumigation-extraction-an automated procedure[J]. Soil Biology and Biochemistry, 1990, 22(8): 1167-1169. DOI:10.1016/0038-0717(90)90046-3 |
| [25] | 吴金水, 林启美, 黄巧云, 等. 土壤微生物生物量测定方法及其应用[M]. 北京: 气象出版社, 2006: 65-71. |
| [26] | Ebhin Masto R, Chhonkar P K, Singh D, et al. Changes in soil biological and biochemical characteristics in a long-term field trial on a sub-tropical inceptisol[J]. Soil Biology and Biochemistry, 2006, 38(7): 1577-1582. DOI:10.1016/j.soilbio.2005.11.012 |
| [27] |
罗希茜, 郝晓晖, 陈涛, 等. 长期不同施肥对稻田土壤微生物群落功能多样性的影响[J]. 生态学报, 2009, 29(2): 740-748. Luo X Q, Hao X H, Chen T, et al. Effects of long-term different fertilization on microbial community functional diversity in paddy soil[J]. Acta Ecologica Sinica, 2009, 29(2): 740-748. DOI:10.3321/j.issn:1000-0933.2009.02.024 |
| [28] |
于维水, 李桂花, 王碧胜, 等. 不同施肥制度下我国东部典型土壤易分解与耐分解碳的组分特征[J]. 植物营养与肥料学报, 2015, 21(3): 675-683. Yu W S, Li G H, Wang B S, et al. Component characteristics of soil labile and recalcitrant carbon under long-term different fertilization systems in eastern China[J]. Journal of Plant Nutrition and Fertilizer, 2015, 21(3): 675-683. |
| [29] | Kuzyakov Y. Review:factors affecting rhizosphere priming effects[J]. Journal of Plant Nutrition and Soil Science, 2002, 165(4): 382-396. DOI:10.1002/1522-2624(200208)165:4<382::AID-JPLN382>3.0.CO;2-# |
| [30] |
吴林坤, 林向民, 林文雄. 根系分泌物介导下植物-土壤-微生物互作关系研究进展与展望[J]. 植物生态学报, 2014, 38(3): 298-310. Wu L K, Lin X M, Lin W X. Advances and perspective in research on plant-soil-microbe interactions mediated by root exudates[J]. Chinese Journal of Plant Ecology, 2014, 38(3): 298-310. |
| [31] | Geyer K M, Kyker-Snowman E, Grandy A S, et al. Microbial carbon use efficiency:accounting for population, community, and ecosystem-scale controls over the fate of metabolized organic matter[J]. Biogeochemistry, 2016, 127(2-3): 173-188. DOI:10.1007/s10533-016-0191-y |
| [32] | Dijkstra P, Salpas E, Fairbanks D, et al. High carbon use efficiency in soil microbial communities is related to balanced growth, not storage compound synthesis[J]. Soil Biology and Biochemistry, 2015, 89: 35-43. DOI:10.1016/j.soilbio.2015.06.021 |
| [33] | Spohn M, Chodak M. Microbial respiration per unit biomass increases with carbon-to-nutrient ratios in forest soils[J]. Soil Biology and Biochemistry, 2015, 81: 128-133. DOI:10.1016/j.soilbio.2014.11.008 |
| [34] |
郭芸, 孙本华, 王颖, 等. 长期施用不同肥料塿土PLFA指纹特征[J]. 中国农业科学, 2017, 50(1): 94-103. Guo Y, Sun B H, Wang Y, et al. PLFA fingerprint characteristics of an anthropogenic loess soil under long-term different fertilizations[J]. Scientia Agricultura Sinica, 2017, 50(1): 94-103. |
| [35] | Klimek B, Chodak M, Jaźwa M, et al. Functional diversity of soil microbial communities in boreal and temperate scots pine forests[J]. European Journal of Forest Research, 2016, 135(4): 731-742. DOI:10.1007/s10342-016-0968-5 |
| [36] | van Groenigen K J, Forristal D, Jones M, et al. Using metabolic tracer techniques to assess the impact of tillage and straw management on microbial carbon use efficiency in soil[J]. Soil Biology and Biochemistry, 2013, 66: 139-145. DOI:10.1016/j.soilbio.2013.07.002 |