 2019, Vol. 40
2019, Vol. 40



2. 中国地质科学院岩溶地质研究所, 自然资源部、广西岩溶动力学重点实验室, 桂林 541004
2. Key Laboratory of Karst Dynamics Laboratory, Ministry of Natural Resources & Guangxi, Institute of Karst Geology, Chinese Academy of Geological Sciences, Guilin 541004, China
地球系统科学认为地球具有一个由岩石圈、大气圈、水圈和生物圈构成的表层系统, 其中生物圈在地球表层系统中拥有不可忽视的作用[1].从17世纪CO2的发现和18世纪发现光合作用以来, 人类对植物光合作用及其对自然环境影响的探索已延续了近400年[2].
岩溶系统属于表层地球化学开放系统, 与岩石圈、大气圈、水圈和生物圈密切相关, 尤其是生物作用的介入, 使得岩溶作用不再是传统意义上的纯无机过程[3, 4].随着岩溶学研究的深入, 生物地球化学过程在岩溶作用中的地位日益突显.水生植物作为河流生态系统的重要组成部分, 是河流生态系统的基本结构框架, 对河流水化学昼夜动态变化具有重要的影响[5].早在19世纪, 人类就已经认识到生物地球化学过程的昼夜与季节变化现象及其对河流理化性质的影响[3].自20世纪90年代以来, 得益于高分辨率自动化在线监测仪器和高频率自动化取样技术的日趋完善, 越来越多的研究开始关注河流昼夜尺度的生物地球化学过程[3].河流昼夜尺度的动态变化研究一方面有助于揭示水体中相对快速的生物地球化学过程, 同时也有助于判别影响河流水化学变化的主导过程[3].章程等[6]对桂林潮田河的研究表明:pH、DO、EC(电导率)、HCO3-和Ca2+的昼夜变化动态反映了光合作用和钙沉积的影响, 水生植物光合作用DIC平均转化速率为0.64×10-5 mmol ·(L ·s)-1, 日转化量占总输入量的4%.张红波等[7]对漓江阳朔断面的监测表明:在昼夜尺度上, 水温、水生植物生理活动以及降水稀释效应共同影响河流水化学昼夜动态变化, 同时指出河流流量是影响河流碳通量的首要因素, 平均碳通量为1 525 g ·s-1.陈波等[8]对茂兰拉桥泉的研究表明:沉水植物生长量较小的泉口和下游池水的水化学和δ13CDIC昼夜变化小于沉水植物大量生长的中游池水, 有机碳通量达336 t ·(km2 ·a)-1.张陶等[9]对广西官村地下河的研究表明:岩溶溪流中DO、p(CO2)、pH和DIC的昼夜变化主要受水生植物代谢的控制, 水生植物光合作用DIC日转化量为0.94 kg ·d-1.由此可见, 前人对不同类型的岩溶水体水化学昼夜动态变化及水生生态系统岩溶碳汇量有了较为详细的研究, 但大部分研究都集中于夏季且主要为单一季节, 具有一定的偶然性而对不同季节水化学昼夜动态变化对比研究较少.因此, 探讨岩溶区河流不同季节水化学昼夜动态变化规律与水生植物光合作用对水体溶解无机碳向有机碳的转化速率有着十分重要的意义[10].本文以广西漓江流域省里断面为例, 利用高分辨率自动化在线监测和高频率取样工作, 探讨pH、电导率、溶解氧等河流物理化学参数以及水体营养元素、溶解无机碳同位素的昼夜动态变化和季节变化特征以及岩溶水体中DIC向OC转换速率, 分析了影响这些参数的驱动因素和过程.
1 材料与方法 1.1 研究区概况漓江流域位于广西壮族自治区东北部, 属于珠江一级支流桂江的上游河段, 地理坐标:109°45′05″~111°02′10″E, 24°16′07″~26°21′11″N.漓江发源于广西兴安县的越城岭老山界南侧, 由北向南依次流经兴安县、灵川县、桂林市区和阳朔县, 止于平乐县恭城河口, 全长164 km, 流域总面积12 680 km2, 漓江流域为长形多支流河系, 整个流域以漓江为轴线, 呈南北狭长带状分布[11](图 1).漓江流域属于中亚热带季风气候区, 年平均气温为16.5~20.0℃, 多年平均降雨量为1 367.5~1 932.9 mm, 雨量充沛, 雨热同期.漓江径流主要来源于大气降水, 年内各月径流分配与流域降水量年内分配相似.漓江汛期为每年4~8月, 5~6月达到最高水位, 8月汛期结束, 径流量约占全年总径流量的70%~80%, 11月至翌年2月为枯水期, 其它时间为平水期, 多年平均流量为128 m3 ·s-1, 流量、水位随降水季节变化明显[12].漓江流域上游为非岩溶区, 岩性主要为花岗岩、碎屑岩.漓江流域中南部为覆盖型及裸露型岩溶区, 广泛分布中、上泥盆统和下石炭统浅海相-滨海相碳酸盐岩, 是岩溶发育的物质基础.因流域内碳酸盐岩质纯层厚, 加之雨热同期的季风气候条件, 岩溶发育强烈, 碳酸盐岩峰丛、峰林地貌广布[11~13].省里断面位于广西灵川县省里村, 地理坐标为110°25′51″E, 25°09′53″N, 由良丰河、桃花江、潮田河和甘棠江等岩溶河流汇入, 以覆盖型和裸露型碳酸盐岩为主.省里断面水生植物生长茂盛, 水生植物以苦草、水绵和黑叶轮藻为主.省里断面以上, 漓江流经灵川县和桂林市区, 城市及城镇分布密集, 社会经济和人口发展迅速, 加之沿线旅游景区和配套设施的开发对生态环境造成了一定程度的破坏.

|
图 1 漓江流域以及省里断面位置 Fig. 1 Map of the Lijiang River and location of the Shengli site |
监测和取样工作分别于2016年10月30日~11月1日(秋季)和2017年9月14~16日(夏季)进行.秋季监测期间出现一次降温过程, 日均温在12~20℃.夏季天气晴朗, 日均温为24~34℃.利用便携式水质分析仪(法国PONSEL)监测水温(T)、pH、溶解氧(DO)、电导率(EC)等水质参数, 精度分别为0.01℃、0.01pH单位、0.01 mg ·L-1和1 μS ·cm-1, 记录间隔设定为15 min.便携式水质分析仪在监测之前进行校正, 其中pH值用4.01和7.01两种标准溶液进行校准, 电导率用1 413μS ·cm-1标准液进行校准.
采样工作每间隔2 h进行一次, 两次监测各采集168个样品.用德国Merck公司生产的碱度计现场测定水体碱度, 计算出水体HCO3-浓度, 精度为0.1 mmol ·L-1.Ca2+质量浓度采用硬度计滴定, 精度为2mg ·L-1.用清洁的聚乙烯瓶采集水样, 采样之前用水样润洗瓶子3次, 样品采集时不留气泡并用封口膜密封.取600 mL水样存储于润洗过的高密度聚乙烯瓶中并加入2滴HgCl2溶液, 抑制微生物活动, 用于DOC含量测试, 另取600 mL水样存储于润洗过的高密度聚乙烯瓶中用于NH4+测试.取50 mL过滤水样储存于高密度聚乙烯瓶中, 用于阴离子测定.另取过滤水样装于润洗过的30 mL聚乙烯瓶中, 分别加入2~3滴浓HNO3和2滴HgCl2溶液, 用于阳离子和水体溶解无机碳同位素(δ13CDIC)测定.用于C/N、POC含量和δ13CPOC测试的玻璃纤维膜预先在马弗炉灼烧3 h(450℃)以去除无机碳.所有样品采集完成后运至室内4℃冷藏.
1.2.2 室内分析阴离子、DOC测试工作在自然资源部/广西岩溶动力学重点实验室完成, 阴离子采用ICS-900离子色谱仪进行分析, DOC含量采用德国MULTI N/C-3100总有机碳/总氮分析仪进行测定.阳离子测试工作在西南大学地理科学学院岩溶环境重庆市重点实验室完成, 采用美国Perkin Elmer公司生产的ICP-OES Optima 2100DV电感耦合等离子体发射光谱仪进行测定. NH4+测试分析由自然资源部岩溶地质资源环境监督检测中心完成. POC、C/N、δ13CPOC和δ13CDIC样品送至国家海洋局第三海洋研究所测试中心进行检测, δ13CDIC采用Gas BenchⅡ-IRMS进行测试(δ13C以V-PDB标准给出), 检测精度为±0.08‰. POC含量和δ13CPOC值利用Thermo公司生产的元素分析仪-稳定同位素质谱仪联机(Flash EA 1112 HT-Delta V Advantages)测定.
δ13CDIC和δ13CPOC测试结果采用V-PDB(Vienna Pee Dee Belemnite)进行标准化, 其值按方程(1)计算:

|
(1) |
式中, R(13C/12C)标准为国际标准物V-PDB的碳同位素丰度比值.
利用现场监测的水温(T)、pH以及实验室内分析测试出的K+、Na+、Ca2+、Mg2+、Cl-、SO42-和HCO3-, 通过WATSPEC软件计算水体二氧化碳分压[p(CO2)]和方解石饱和指数(SIC).二氧化碳分压按方程(2)计算:
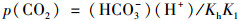
|
(2) |
式中, Kh和K1分别是H2CO3和CO2的平衡常数.
方解石饱和指数(SIC)按方程(3)计算:
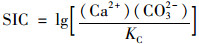
|
(3) |
文中不同监测月份平均流量数据由桂林和潮田河水文站提供.
2 结果与分析 2.1 秋季省里断面物理化学参数的昼夜变化监测期间省里断面水文参数[T、pH、EC、DO、p(CO2)、SIC]以及Ca2+、HCO3-、NO3-等离子含量和δ13CDIC均表现出明显的有规律的昼夜变化, 并且表现出两种不同的变化模式, 其中T、pH、SIC、DO、δ13CDIC表现出白天上升夜间下降的趋势, EC、Ca2+、HCO3-、p(CO2)和NO3-等离子含量则表现出白天下降夜间上升的趋势.同时DOC浓度也表现出白天上升夜间下降的变化, 但变化趋势没有其他物理化学参数明显.
水温(T)变化范围为17.66~21.49℃, 平均值为19.58℃, 昼夜变化幅度3.83℃. 10月30日最高值出现在15:00~16:00, 10月31日由于出现降温过程, 水温最高值出现在11:00~13:00. 10月31日水温最低出现在清晨07:00左右, 由于降温过程延缓了水温的回升过程, 11月1日最低温出现在09:00左右(图 2).溶解氧(DO)昼夜变化趋势与水温变化趋势基本一致, 变化范围为5.77~25.17mg ·L-1, 平均值为12.65 mg ·L-1. DO峰值出现在日落前后, 滞后于水温峰值约2 h. 10月31日受降温过程影响, DO在13:30~14:30达到最大值.夜间随着水温的降低, DO含量逐渐降低, 凌晨04:00左右至日出前后达到最低值并呈现平坦的低值宽谷(图 3).

|
图 2 秋季省里断面水温和pH的昼夜变化 Fig. 2 Diurnal variation of the temperature and pH at the Shengli site in the month representing autumn |

|
图 3 秋季省里断面溶解氧和电导率昼夜变化 Fig. 3 Diurnal variation of the DO and EC at the Shengli site in the month representing autumn |
监测期间水体pH值介于7.06~9.08之间, 平均值为7.94, 昼夜变化幅度为2.02, 呈弱碱性. pH值的变化趋势与水温相似, 在白天14:00~16:00达到最高值, 清晨08:00左右达到最低值(图 2). p(CO2)在凌晨(04:00~08:00)达到最大值, 在下午(12:00~16:00)达到最小值, 变化范围为46.77~8511.38ng ·L-1, 平均值为2145.27 ng ·L-1. SIC与p(CO2)呈相反的变化趋势, 中午(12:00~14:00)达到最大值, 而在日出前(06:00~08:00)达到最小值, 变化范围介于-0.77~0.99, 平均值为0.11(图 4).

|
图 4 秋季省里断面二氧化碳分压和方解石饱和指数昼夜变化 Fig. 4 Diurnal variation of the pCO2 and SIC at the Shengli site in the month representing autumn |
EC变化范围为201.9~258μS ·cm-1, 平均值为229.11μS ·cm-1(图 3); HCO3-含量变化范围为67.1~115.9 mg ·L-1, 平均值为96.5 mg ·L-1, Ca2+含量的变化范围为34.47~40.13 mg ·L-1, 平均值为37.17 mg ·L-1(图 5). EC、HCO3-和Ca2+的昼夜动态趋势基本一致, 监测期间EC、HCO3-和Ca2+在中午(12:00~16:00)处于相对的低值区, 而在夜间和清晨(00:00~8:00)则处于高值区, 即EC、HCO3-和Ca2+夜间高于白天.

|
图 5 秋季省里断面碳酸氢根和钙离子昼夜变化 Fig. 5 Diurnal variation of the HCO3- and Ca2+ at the Shengli site in the month representing autumn |
DOC和POC含量小时数据表现出快速变动的特点. DOC浓度变化范围为3.88~6.25 mg ·L-1, 平均值为5.06 mg ·L-1, 在下午13:00~15:00达到最大值, 而在06:00~08:00达到最小值. POC浓度介于0.21~0.76 mg ·L-1平均值为0.39 mg ·L-1, 昼夜变化趋势与DOC基本一致(图 6). δ13CPOC变化范围为-25.61‰~-22.52‰, 平均值为-24.45‰, 昼夜变化幅度为-1.93‰. δ13CDIC变化范围为-9.9‰~-7.8‰, 平均值为-8.9‰, 昼夜变化幅度为-2.1‰. δ13CDIC在白天偏重晚上偏轻, 在下午16:00左右达到最大值, 清晨05:30~08:00达到最小值与DIC峰值相反(图 7).

|
图 6 秋季省里断面溶解有机碳和颗粒有机碳昼夜变化 Fig. 6 Diurnal variation of the DOC and POC at the Shengli site in the month representing autumn |

|
图 7 秋季省里断面δ13CPOC和δ13CDIC昼夜变化 Fig. 7 Diurnal variation of the δ13CPOC and δ13CDIC at the Shengli site in the month representing autumn |
NO3-、SO42-、Cl-、Na+和K+等营养元素质量浓度的昼夜变化表现出一定的规律性, 这5种离子含量均表现为白天低夜间高, 上午离子含量有逐渐降低的趋势, 傍晚逐渐升高, 在下午14:00~16:00含量降至最低值, 凌晨04:00~06:00升至最高值. NO3-浓度变化范围为6.80~9.21 mg ·L-1, 平均值为8.03 mg ·L-1. SO42-浓度变化范围为11.12~13.03 mg ·L-1, 平均值为12.08 mg ·L-1(图 8).Cl-浓度变化范围为5.70~6.64 mg ·L-1, 平均值为5.99 mg ·L-1(图 9). Na+浓度变化范围为4.82~5.64 mg ·L-1, 平均值为5.13 mg ·L-1(图 9). K+浓度变化范围2.11~2.44 mg ·L-1,平均值为2.25 mg ·L-1. Mg2+浓度在10月30日和10月31日的变化趋势与NO3-、SO42-、Cl-、Na+和K+保持一致,而在11月1日清晨上升并保持在较高的浓度,变化范围为2.36~2.96 mg ·L-1,平均值为2.63 mg ·L-1(图 10).

|
图 8 秋季省里断面SO42-和NO3-昼夜变化 Fig. 8 Diurnal variation of the SO42- and NO3- at the Shengli site in the month representing autumn |

|
图 9 秋季省里断面Cl-和Na+昼夜变化 Fig. 9 Diurnal variation of theCl- and Na+ at the Shengli site in the month representing autumn |

|
图 10 秋季省里断面Mg2+和K+昼夜变化 Fig. 10 Diurnal variation of the Mg2+ and K+ at the Shengli site in the month representing autumn |
监测期间, 水化学昼夜变化特征与2016年10月30日~11月1日基本一致, 水温(T)、pH、DO、SIC、δ13CDIC白天呈上升趋势, 夜间呈下降趋势; HCO3-、Ca2+、NO3-等离子含量、EC和p(CO2)白天降低夜间上升, 具体结果分析如下.
水温(T)变化范围为25.46~29.88℃, 平均值为27.40℃, 昼夜变化幅度4.42℃.最高值出现在14:45~15:45, 最低值出现在06:00~08:00(图 11).溶解氧(DO)昼夜变化趋势与水温变化趋势基本一致, 变化范围为5.86~11.44 mg ·L-1, 平均值为8.38 mg ·L-1. DO峰值出现在日落前后(17:00~18:00), 滞后于水温峰值约2h.夜间随着水温的降低, DO含量逐渐降低, 午夜(00:00)至清晨(08:00)达到最低值并呈现平坦的低值宽谷, 之后随水体温度上升DO含量逐渐升高(图 12).

|
图 11 夏季省里断面水温和pH的昼夜变化 Fig. 11 Diurnal variation of the temperature and pH at the Shengli site in the month representing summer |

|
图 12 夏季省里断面溶解氧和电导率昼夜变化 Fig. 12 Diurnal variation of the DO and EC at the Shengli site in the month representing summer |
监测期间, 水体的pH值介于7.51~8.99, 平均值为8.11, 昼夜变化幅度为1.48, 呈弱碱性. pH值的变化趋势与DO相似, 下午14:45~17:45达到最高值, 午夜(00:00)至清晨(08:00)达到最低值(图 11). p(CO2)在凌晨(00:00~08:00)达到最大值, 在下午(12:00~18:00)达到最小值, 变化范围为81.10~3723.96 ng ·L-1, 平均值为1474.79 ng ·L-1. SIC峰值与p(CO2)相反, 在中午(12:00~18:00)达到最大值, 而在凌晨至清晨(00:00~08:00)达到最小值, 变化范围介于-0.32~1.12, 平均值为0.32(图 13).

|
图 13 夏季省里断面二氧化碳分压和方解石饱和指数昼夜变化 Fig. 13 Diurnal variation of the p(CO2) and SIC at the Shengli site in the month representing summer |
EC的变化范围为189.11~210.95μS ·cm-1, 平均值为201.08μS ·cm-1(图 12); HCO3-含量的变化范围为103.70~122.00 mg ·L-1, 平均值为114.49 mg ·L-1, Ca2+含量的变化范围为34.00~50.00 mg ·L-1, 平均值为38.46 mg ·L-1(图 14). EC、HCO3-和Ca2+的昼夜动态趋势基本一致, 在中午(12:00~14:00)处于相对的低值区, 而在夜间和清晨(00:00~06:00)则处于高值区. EC在9月16日夜间出现异常下降, 但总体上EC值夜间高于白天.

|
图 14 夏季省里断面碳酸氢根和钙离子昼夜变化 Fig. 14 Diurnal variation of the HCO3- andCa2+ at the Shengli site in the month representing summer |
DOC浓度变化范围为3.19~5.09 mg ·L-1, 平均值为3.87 mg ·L-1. POC浓度介于3.66~5.73 mg ·L-1平均值为4.45 mg ·L-1. DOC和POC数据表现出快速波动的特点, 但总体上白天高于夜间(图 15). δ13CPOC变化范围为-29.91‰~-21.80‰, 平均值为-24.42‰, 昼夜变化幅度为-8.11‰, 未表现出明显的昼夜变化规律. δ13CDIC变化范围为-12.15‰~-7.80‰, 平均值为-10.08‰, 昼夜变化幅度为-4.35‰. δ13CDIC在白天偏重晚上偏轻, 在上午10:00至下午16:00左右处于高值区, 夜间20:00至清晨08:00处于低值区(图 16).

|
图 15 夏季省里断面溶解有机碳和颗粒有机碳昼夜变化 Fig. 15 Diurnal variation of the DOC and POC at the Shengli site in the month representing summer |

|
图 16 夏季省里断面δ13CDIC和δ13CPOC昼夜变化 Fig. 16 Diurnal variation of the δ13CDIC and δ13CPOC at the Shengli site in the month representing summer |
NO3-、SO42-、Cl-和Mg2+等营养元素在下午14:00~18:00含量降至最低值, 夜间离子含量上升并处于高值区. K+和Na+浓度在9月14日变化趋势与NO3-、SO42-、Cl-和Mg2+保持一致(图 17和图 18), 而在9月15日夜间开始下降并在清晨08:00达到最小值, 之后呈现上升趋势, 并于9月16日稳定在较高的值域(图 19). NO3-浓度变化范围为6.73~7.44 mg ·L-1, 平均值为7.14 mg ·L-1. SO42-浓度变化范围为12.64~14.26 mg ·L-1, 平均值为13.31 mg ·L-1.Cl-浓度变化范围为3.49~3.90 mg ·L-1, 平均值为3.69 mg ·L-1. Na+浓度变化范围为2.98~3.57 mg ·L-1, 平均值为3.31 mg ·L-1. Mg2+浓度变化范围为2.39~2.90 mg ·L-1, 平均值为2.59 mg ·L-1. K+浓度变化范围为1.5~1.93 mg ·L-1,平均值为1.78 mg ·L-1.

|
图 17 夏季省里断面SO42-和NO3-昼夜变化 Fig. 17 Diurnal variation of the SO42- and NO3- at the Shengli site in the month representing summer |

|
图 18 夏季省里断面Cl-和Mg2+昼夜变化 Fig. 18 Diurnal variation of the Cl- and Mg2+ at the Shengli site in the month representing summer |

|
图 19 夏季省里断面Na+和K+昼夜变化 Fig. 19 Diurnal variation of the Na+ and K+ at the Shengli site in the month representing summer |
白天, 水生植物光合作用消耗DIC, 促进碳酸钙沉积; 夜间, 水生植物呼吸作用取代光合作用而占据主导地位, 呼吸作用产生CO2, 加速碳酸钙溶解[3].此过程中发生了以下反应:
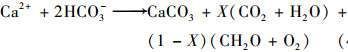
|
(4) |
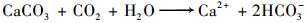
|
(5) |

|
(6) |

|
(7) |
白天, 在有光照的条件下[方程(4)], 水生植物消耗水中的HCO3-, 合成有机质, 释放O2, 加速了碳酸钙沉积, Ca2+和HCO3-含量以及EC值下降, DO含量、pH值升高.一部分HCO3-被水生植物通过利用PEPC(烯醇丙酮酸磷酸羧激酶)经CCM(CO2浓缩机制)途径固定下来, 再通过碳酸酐酶(CA)把HCO3-转化成CO2, 最后再由卡尔文循环固碳[14~16] [方程(6)]; 一部分CO2通过溶解CaCO3被固定[方程(5)], 剩余部分释放进入空气中.夜间, 水生植物呼吸作用成为主导的生理活动, 呼吸作用消耗有机质和溶解氧, 产生CO2[方程(7)], 一部分CO2重新溶解沉淀的碳酸钙[方程(5)], 转化为HCO3-, 剩余部分释放到大气中, Ca2+和HCO3-含量以及EC值升高, DO含量、pH值降低.
受气温影响, 秋季省里断面水温整体上呈下降趋势, 但仍显示出白天升高夜间下降的昼夜变化.夏季天气晴朗且监测期间气温波动不大, 省里断面水温表现出明显的昼夜动态变化, 且具有很好的重现性.省里断面水化学明显的昼夜变化与水生植物光合作用和呼吸作用过程表现出一致性(由溶解氧的变化体现), 说明省里断面水体昼夜变化主要受水生植物生理活动的影响.
省里断面p(CO2)昼夜变幅大, 两次昼夜监测变异系数分别为1.23和0.89, 说明水生植物光合作用消耗水体DIC以及伴随的钙沉降能显著影响方解石饱和指数(SIC)和p(CO2).白天光合作用利用DIC导致p(CO2)下降, SIC上升, 水体处于过饱和状态, p(CO2)处于与大气平衡的临界线附近(389ng ·L-1), 水体CO2脱气量减小, 说明水生植物光合作用将水体DIC以有机碳的形式存储, 同时减少水气界面CO2逸出量, 从而提高了岩溶碳汇的稳定性; 夜间呼吸作用产生CO2导致p(CO2)快速上升, SIC下降, 水体处于不饱和状态, 水体p(CO2)远高于大气, 可能发生潜在的脱气作用[17].
虽然pH、DO、SIC与水温(T)呈现一致的变化趋势, 但其最高值与最低值的出现并不与水温(T)同步. pH、DO、SIC最高值的出现要滞后于水温1~2 h, 例如2016年10月31日水温最高值出现在12:00~13:00, 而pH、DO、SIC在14:00左右达到最高值.最低值要提前水温最低值大约1 h, 例如2016年10月31日pH、DO、SIC在清晨05:00~06:00就已经达到最低值并呈现平坦的低值宽谷而水温最低值出现在07:00左右. pH、DO、SIC最高值和最低值与水温(T)不同步的原因可能是:白天水温上升到最高值后随气温的下降而降低, 但光照和温度条件仍能满足以水生植物光合作用占主导的新陈代谢过程, 光合作用仍然在进行, 水生植物光合作用的主要表征DO和pH值继续上升.与光合作用的积累过程不同, 呼吸作用是一个相对快速消耗过程, 夜间随着水生植物呼吸作用消耗水体溶解氧, 释放二氧化碳, DO和pH值逐步降低, 并且随着水温(T)降低, 消耗速率逐步降低, 在清晨05:00~06:00就已经达到最低值并趋于稳定.同时, 与秋季相比, 夏季的光照时间长, 水生植物能够进行光合作用的时间也相应要长, 作为表征水生植物生理过程的主要指标, DO和pH值在18:00到达峰值, 明显晚于秋季(14:00).
水生植物通过光合作用合成有机质, 可能发生了以下两个反应[3, 18]:
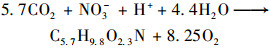
|
(8) |
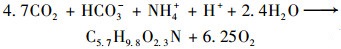
|
(9) |
通过夏季昼夜监测的取样和测试发现, 省里断面NH4+浓度多数时段低于检测限(<0.02mg ·L-1, 图 20), 而NO3-浓度表现出白天下降夜间上升的昼夜变化.两次昼夜监测期间NO3-与DO的相关性分别为-0.672和-0.516, 呈现明显的负相关关系, 即随着光合作用的进行[方程(8)], NO3-逐渐被消耗, 并不断产生O2, 使得水体中溶解氧含量上升.同时两次昼夜监测期间, SO42-、Cl-、Na+、K+和Mg2+等营养元素与DO表现出负相关性, 相关系数分别为-0.303、-0.602、-0.488、-0.659、-0.427(表 1)和-0.443、-0.649、-0.672、-0.491、-0.412(表 2), 说明水生植物光合作用在利用NO3-合成有机质的同时SO42-、Cl-、Na+、K+和Mg2+等营养元素也参与其中, 受生物代谢过程(同化作用)控制[17].通常情况下夏季温度和太阳辐射量高于秋季, 水生植物光合作用更加活跃, 即生物量越高, NO3-、Cl-、Na+、K+和Mg2+等营养元素消耗量与DO生成量越高; 反之则越低.对比秋季和夏季两次昼夜监测数据发现, 夏季省里断面水温变化范围为25.46~29.88℃, 日变化幅度为4.42℃, 均高于秋季(17.66~21.49℃,3.83℃); 而水体DO含量和SO42-、NO3-、Cl-、Na+、Mg2+等营养元素消耗量均低于秋季, 尤其是DO变化幅度(5.86~11.44mg ·L-1)和均值(8.38mg ·L-1)远低于秋季(变化幅度为5.77~25.17mg ·L-1, 平均值为12.65mg ·L-1), 说明夏季省里断面的光合作用活跃程度低于秋季.分析原因可能是:一方面, 2017年8月中旬漓江水位暴涨, 洪水对省里断面水生植物群落和生态系统产生较大的破坏作用, 至9月中旬仍未得到恢复; 另一方面, 夏季水体浊度较高, 削弱一部分太阳辐射, 使得进入水生生态系统的辐射量减少, 因此在天气晴朗且水温较高的条件下产生了较低的净光合作用.两次昼夜监测期间省里断面水化学显著的昼夜变化过程与水生植物生理活动表现出一致性(由DO的变化表征), 说明省里断面水化学的昼夜变化主要受水生植物生理过程控制, 同时季节变化和水生生态系统的变迁也对省里断面水化学昼夜变化产生一定程度的影响.

|
图 20 夏季省里断面NH4+昼夜变化 Fig. 20 Diurnal variation of the NH4+ at the Shengli site in the month representing summer |
|
|
表 1 秋季省里断面营养元素与溶解氧相关性系数1) Table 1 Coefficients of correlation between the nutrients and dissolved oxygen at the Shengli site in the month representing autumn |
|
|
表 2 夏季省里断面营养元素与溶解氧相关性系数1) Table 2 Coefficients of correlation between the nutrients and dissolved oxygen at the Shengli site in the month representing summer |
分析发现省里断面夏季水体总有机碳(TOC=POC+DOC)和POC质量浓度均高于秋季, 尤其是POC质量浓度(3.66~5.73 mg ·L-1)远高于秋季(0.21~0.76 mg ·L-1), 这与前文分析秋季省里断面具有更高的净初级生产力并不一致.一般来说水体中的有机碳主要来源于陆生生物和水生生物的初级生产力, 可以利用C/N计算内源有机碳占总有机碳的比例.典型藻类光合作用形成产物为C5.7H9.8O2.3N, 其C/N应为5.7, 以C/N值5.7为内源端元, 20.69为外源端元, 可以利用端元混合模型计算内源有机碳占总有机碳的比例[19], 方程如下:
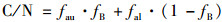
|
(10) |
式中, C/N为样品的C/N值; fau为C/N值的内源端元, 取5.7; fal为C/N值的外源端元, 取20.69; fB为内源有机碳占总有机碳的比例.
夏季监测期间, 省里断面C/N值介于9.57~10.88, 平均为10.09;秋季C/N值介于6.56~7.68, 平均为7.04.计算可得省里断面夏季内源有机碳占总有机碳的比例介于65.44%~74.22%, 平均为70.71%;秋季内源有机碳占总有机碳的比例介于86.79%~94.25%, 平均为91.07%, 表明秋季省里断面内源有机碳占总有机碳的比例高于夏季, 这与秋季水生植物具有更高的净初级生产力一致.因此, 夏季省里断面较高的TOC和POC质量浓度可能与陆源有机碳的补给有关.夏季监测取样前一周内, 漓江流域有中到大雨, 地表径流携带大量的陆源有机碳进入漓江, 导致夏季水体TOC和POC质量浓度高于秋季.
水生植物光合作用利用HCO3-占无机碳源的比例, 可以利用吴沿友[20]等人的专利计算.模型中两个端元分别为:不利用HCO3-的悬铃木叶片13C值为-31.56‰, 极少利用CO2的小球藻13C值为-22.74‰.计算方程如下:
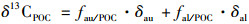
|
(11) |
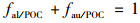
|
(12) |
式中, δ13CPOC为样品的碳同位素值; δau为极少利用CO2作为碳源而以HCO3-作为无机碳源的端元值, 为-22.74‰; δal为基本不利用HCO3-作无机碳源端元组分的碳同位素值, 为-31.56‰; fau/POC表示植物光合作用利用HCO3-生成的有机碳在内源有机碳中所占的比例, fal/POC表示植物光合作用利用CO2生成的有机碳在内源有机碳中所占的比例.
秋季省里断面δ13CPOC值变化范围介于-25.61‰~-22.52‰, 平均值为-24.45‰, fau/POC介于67.42%~99.75%, 平均值为79.54%.夏季省里断面δ13CPOC值变化范围介于-27.11‰~-22.44‰, 平均值为-24.83‰, fau/POC介于50.43%~99.81%, 平均值为76.59%.表明夏秋季节省里断面水生植物光合作用以HCO3-作为主要的无机碳源.
3.2 水生生物作用对δ13CDIC的控制岩溶地区溶解无机碳(DIC)主要来自碳酸盐岩风化和土壤CO2淋滤.不同来源的DIC其溶解无机碳同位素(δ13CDIC)组成不同, 通过测定水中DIC同位素(δ13CDIC)可以了解水中物理、化学和生物过程的变化[21, 22].在昼夜时间尺度上, 水生植物光合作用和呼吸作用是控制岩溶河流中溶解无机碳(DIC)含量和溶解无机碳同位素(δ13CDIC)变化的主要因素, 而温度变化和脱气作用影响相对较小[22].省里断面δ13CDIC在夏季和秋季两次昼夜监测取样期间, 都表现出最高值出现在16:00左右, 然后晚上逐渐偏轻, 白天偏重的变化规律(图 7和图 16).水生植物光合作用消耗DIC, 优先利用低能量的12C, 导致河流DIC含量降低和13C累积, 因此随着光合作用的进行, 水体δ13CDIC逐渐偏正.夜间, 水生植物生理活动以呼吸作用为主, 呼吸作用释放CO2, 因为加入了同位素较轻的生物成因的CO2, 导致δ13CDIC值偏轻[23~25].清晨06:00~08:00左右δ13CDIC值降至最低, 这一现象指示了省里断面水生植物呼吸效应累积达到最大.与秋季相比, 夏季省里断面δ13CDIC值偏负, 夏季和秋季δ13CDIC平均值分别为-10.08‰和-8.9‰, 表明夏季水生生物的光合作用强度低于秋季, 这与前文地球化学特征昼夜变化表明水生生物光合作用强度夏季低于秋季一致.主要的原因也是由于2017年8月中旬漓江洪水对水生植物群落的破坏以及夏季较高的水体浊度削弱太阳辐射的影响.而秋季漓江处于枯水季, 河水比较清澈, 适合于浮游植物和藻类生长, 光合作用大于呼吸作用, DIC中的12C被优先利用并以有机质的形式储存, 使δ13CDIC值正向偏移.同时, 枯水期的漓江水位低而流速缓, 有利于河流DIC与大气CO2之间建立同位素平衡, 从而使δ13CDIC值正向偏移.此外, 夏季, 土壤根系和土壤微生物的呼吸作用强, 土壤呼吸产生较多的CO2, 溶解后导致河流δ13CDIC偏负; 秋季, 土壤根系和土壤微生物的呼吸作用比夏季弱, 土壤呼吸产生的CO2量减少, 溶解后导致河流δ13CDIC比夏季偏正[25~27].夏季省里断面pH、DO、HCO3-、EC、DOC以及营养元素的昼夜变化量均小于秋季的昼夜变化量, 变化量分别为1.48、5.58mg ·L-1、18.3mg ·L-1、21.84μS ·cm-1、1.9mg ·L-1和2.02、19.4mg ·L-1、48.8mg ·L-1、56.1μS ·cm-1、2.37mg ·L-1, 尤其是作为衡量水生植物活跃程度的DO和pH明显偏小, 说明秋季水生植物生理活动对省里断面水化学特征的影响程度更高, 而夏季省里断面δ13CDIC昼夜变化幅度(-4.35‰)大于秋季(昼夜变化幅度为-2.1‰).通过计算亨利常数发现:两次昼夜监测期间温度对省里断面水化学的影响平均比例分别为10.90%和7.37%(表 3), 显示温度昼夜变化差异对δ13CDIC昼夜变化的影响程度存在一定的差别, 但影响比例差别不大, 水生植物生理活动仍然是控制水化学及同位素昼夜变化的主导因素[26~33].因此, 出现δ13CDIC昼夜变化幅度与水化学参数及营养元素昼夜变化幅度不一致的现象值得进一步深入探究.
|
|
表 3 温度变化对河流p(CO2)的影响比例 Table 3 Influence of temperature onCO2 partial pressure |
3.3 DIC-OC转化速率
岩溶动力系统中碳酸盐岩在传统的无机岩溶作用和生物作用协同下积极参与到全球碳循环中, 成为大气CO2循环的重要环节, 其中陆地水生植物光合作用发挥了重要作用.水生植物光合作用利用水体中DIC, 将岩溶作用风化溶解的DIC转化为有机碳并以有机碳的形式储存, 使陆地水生生态系统也具有与海洋过程相类似的“生物碳泵”作用[34~39].
针对岩溶水生植物光合作用的特点, 可通过与光合作用密切相关的溶解氧为计算指标, 计算水生植物光合作用对岩溶水体中溶解无机碳的固定量.在陆地水生生态系统中, 白天水生植物初级生产力(GPP)即为光合作用固定岩溶水体中DIC的数量, 而水生生态系统的总呼吸作用(ER)为以有机碳(OC)形式固定下来的DIC消耗量.白天, 水中DO的变化主要来自水生植物光合作用放氧、呼吸作用耗氧及与大气的氧交换; 夜间, 水中DO的变化主要来自呼吸作用耗氧及与大气的氧交换.水生植物光合作用产生的O2与呼吸作用消耗的CO2是等摩尔的, 即每消耗1 mol的CO2产生1 mol的O2[方程(13)], 而岩溶水中1 mol的HCO3-分解产生1 mol的CO2.因此, 可以通过计算水体溶解氧的净生产量来估算水生植物净固碳量[方程(14)][39].

|
(13) |
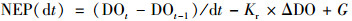
|
(14) |
式中, t为时间(min), DOt-DOt-1为单位时间内溶解氧浓度变化量, ΔDO为氧亏量(mg ·L-1), 即水体中溶解氧饱和时的浓度与监测值之差; Kr为复氧系数, 是水气界面O2交换系数; G为地表水体与地下河水交换对监测点水体溶解氧的影响量[40~44].不同温度(T)下, 饱和溶解氧计算方法如式(15):
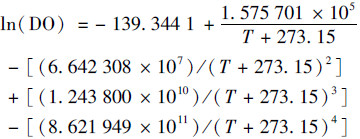
|
(15) |
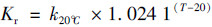
|
(16) |
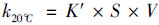
|
(17) |
式中, T为实时监测温度(℃), k20℃为水体温度为20℃时氧气复氧率, 当河流流量大于0.56 m3·s-1时, K′取值为1.53×104, S为河道比降(‰), 取值0.44‰, V为流速(m ·s-1), 2016年10月和2017年9月省里断面平均流速为0.032 5 m ·s-1和0.092 m ·s-1.
通过计算, 实验期间秋季和夏季省里断面水生植物光合作用岩溶碳汇固定量日平均值为2.12mmol ·L-1和0.94mmol ·L-1, 其中秋季显示出更高的固碳效率.以夏季为例, 水生植物光合作用DIC转化过程可以分为4个部分(图 21).第一部分为清晨07:00至上午10:30, 随着太阳辐射增强, 水生植物光合作用开始利用DIC, 导致DO浓度上升, DIC浓度和p(CO2)降低.由于较低的太阳辐射和水温, 光合作用强度小于呼吸作用, 水生植物固碳量为负值.第二部分为上午10:30至下午17:30, 随着太阳辐射增强和水体温度上升, 水生植物光合作用强度大于呼吸作用, 水生植物固碳量为正值并且达到最大值.第三部分为下午17:30至夜间22:00, 随着太阳辐射的减弱, 水生植物光合作用强度逐渐减小, 水生植物利用DIC量减小, 河流DIC浓度和p(CO2)上升, 水生植物固碳量重新成为负值.第四部分为夜间22:00至清晨08:00, 水生植物固碳量仍为负值, 由于缺乏太阳辐射, 水生植物净固碳量等于呼吸作用消耗量, 在此期间河流DIC浓度和p(CO2)达到最大值[44].

|
图 21 省里断面夏季和秋季溶解氧净生产量昼夜变化 Fig. 21 Diurnal variation of the dissolved oxygen net production at the Shengli site in the months representing summer and autumn |
(1) 高分辨率在线监测和高频率取样结果表明:受水生植物光合作用和呼吸作用控制, 夏秋季节漓江流域省里断面河流理化性质有着明显的昼夜变化, 且表现出不同的变化模式:水温、DO、pH、SIC白天升高, 夜晚降低, p(CO2)、EC、HCO3-、Ca2+则表现出白天降低夜间上升的变化趋势, 且秋季昼夜变化幅度大于夏季.
(2) SO42-、NO3-、Cl-、Na+、K+和Mg2+等营养元素的昼夜变化主要受水生植物代谢过程(同化作用)控制, 表现出白天下降夜间上升的变化趋势.由于2017年8月中旬洪水对省里断面水生植物群落的破坏和水体浊度的影响, 营养元素的昼夜变化幅度表现为秋季大于夏季, 这与水生植物光合作用的主要表征DO和pH值变化幅度相一致.
(3) δ13CDIC昼夜变化主要受水生植物光合作用和呼吸作用控制, 表现出白天偏重夜间偏轻的昼夜变化规律.受土壤根系和土壤微生物呼吸作用以及河流水文要素季节变化的影响, 夏季δ13CDIC值偏轻于秋季.
(4) 通过以溶解氧为计算指标, 得到秋季和夏季省里断面水生植物光合作用对岩溶碳汇固定量日平均值为2.12mmol ·L-1和0.94mmol ·L-1, 秋季显示出更高的固碳效率.
| [1] |
袁道先. 现代岩溶学和全球变化研究[J]. 地学前缘, 1997, 4(1-2): 17-25. Yuan D X. Modern karstology and global change study[J]. Earth Science Frontiers, 1997, 4(1-2): 17-25. |
| [2] |
袁道先. 地质作用与碳循环研究的回顾和展望[J]. 科学通报, 2011, 56(26): 2157. Yuan D X. Foreword for the special topic "Geological Processes in Carbon Cycle"[J]. Chinese Science Bulletin, 2011, 56(35): 3741-3742. |
| [3] |
章程, 汪进良, 蒲俊兵. 地下河出口河流水化学昼夜动态变化——生物地球化学过程的控制[J]. 地球学报, 2015, 36(2): 197-203. Zhang C, Wang J L, Pu J B. Diel aqueous chemical cycling in a typical karst spring-fed stream:controls of biogeochemical processes[J]. Acta Geoscientia Sinica, 2015, 36(2): 197-203. |
| [4] |
张强. 岩溶地质碳汇的稳定性——以贵州草海地质碳汇为例[J]. 地球学报, 2012, 33(6): 947-952. Zhang Q. The stability of carbon sink effect related to carbonate rock dissolution:a case study of the Caohai lake geological carbon sink[J]. Acta Geoscientica Sinica, 2012, 33(6): 947-952. |
| [5] |
苏睿丽, 李伟. 沉水植物光合作用的特点与研究进展[J]. 植物学报, 2005, 22(S1): 128-138. Su R L, Li W. Advances in research on photosynthesis of submerged macrophytes[J]. Chinese Bulletin of Botany, 2005, 22(S1): 128-138. |
| [6] |
章程, 汪进良, 肖琼. 桂林潮田河溶解无机碳来源与昼夜动态变化[J]. 第四纪研究, 2017, 37(6): 1283-1292. Zhang C, Wang J L, Xiao Q, et al. The sources and diurnal changes of dissolved inorganic carbon in chaotian river, Guilin, China[J]. Quaternary Sciences, 2017, 37(6): 1283-1292. |
| [7] |
张红波, 何师意, 于奭, 等. 夏季岩溶区河流的水化学及碳汇动态变化——以桂林漓江为例[J]. 西南师范大学学报(自然科学版), 2013, 38(3): 55-61. Zhang H B, He S Y, Yu S, et al. Dynamic changes to water chemical and carbon sinks of karst river in summer:a case study in the Li River of Guilin[J]. Journal of Southwest China Normal University (Natural Science Edition), 2013, 38(3): 55-61. DOI:10.3969/j.issn.1000-5471.2013.03.012 |
| [8] |
陈波, 杨睿, 刘再华, 等. 水生光合生物对茂兰拉桥泉及其下游水化学和δ13CDIC昼夜变化的影响[J]. 地球化学, 2014, 43(4): 375-385. Chen B, Yang R, Liu Z H, et al. Effects of aquatic phototrophs on diurnal hydrochemical and δ13CDIC variations in an epikarst spring and two spring-fed ponds of Laqiao, Maolan, SW China[J]. Geochimica, 2014, 43(4): 375-385. |
| [9] |
张陶, 蒲俊兵, 袁道先, 等. 广西官村岩溶溪流中生物地球化学控制的DIC和NO3-昼夜变化研究[J]. 地质学报, 2016, 90(8): 1965-1976. Zhang T, Pu J B, Yuan D X, et al. Biogeochemical controls on daily cycling of DIC and NO3-of Guancun karst stream in Guangxi[J]. Acta Geologica Sinica, 2016, 90(8): 1965-1976. DOI:10.3969/j.issn.0001-5717.2016.08.024 |
| [10] |
章程. 岩溶区河流水化学昼夜变化与生物地球化学过程[J]. 中国岩溶, 2015, 34(1): 1-8. Zhang C. Diel aqueous chemistry and biogeochemical processes in streams of karst areas[J]. Carsologica Sinica, 2015, 34(1): 1-8. |
| [11] |
邓自强, 林玉石, 张美良, 等. 桂林地质构造与岩溶地貌发育的时序关系[J]. 中国岩溶, 1986, 5(4): 289-296. Deng Z Q, Lin Y S, Zhang M L, et al. Time-sequence relationship between geological structures and development of karst features in Guilin area[J]. Carsologica Sinica, 1986, 5(4): 289-296. |
| [12] | 韩耀全, 周解, 吴祥庆. 漓江的自然地理与水质调查[J]. 广西水产科技, 2007(2): 8-16. |
| [13] | 黄家城. 桂林漓江志[M]. 南宁: 广西人民出版社, 2004. |
| [14] | Reiskind J B, Berg R H, Salvucci M E, et al. Immunogold localization of primary carboxylases in leaves of aquatic and A C3-C4 intermediate species[J]. Plant Science, 1989, 61(1): 43-52. DOI:10.1016/0168-9452(89)90117-9 |
| [15] | Magnin N C, Cooley B A, Reiskind J B, et al. Regulation and localization of key enzymes during the induction of kranz-less, C4-type photosynthesis in Hydrilla verticillata[J]. Plant Physiology, 1997, 115(4): 1681-1689. DOI:10.1104/pp.115.4.1681 |
| [16] | Liu Y, Liu Z H, Zhang J L, et al. Experimental study on the utilization of DIC by Oocystis solitaria Wittr and its influence on the precipitation of calcium carbonate in karst and non-karst waters[J]. Carbonates and Evaporites, 2010, 25(1): 21-26. DOI:10.1007/s13146-009-0002-9 |
| [17] |
章程, 肖琼, 苗迎, 等. 广西桂林漓江典型河段水化学昼夜动态变化及其对岩溶碳循环的影响[J]. 地球学报, 2018, 39(5): 613-621. Zhang C, Xiao Q, Miao Y, et al. Day and night aqueous chemical changes and their impact on karst carbon cycle at typical monitoring sites of the Lijiang river, Guilin, Guangxi[J]. Acta Geoscientica Sinica, 2018, 39(5): 613-621. |
| [18] | Gammons C H, Babcock J N, Parker S R, et al. Diel cycling and stable isotopes of dissolved oxygen, dissolved inorganic carbon, and nitrogenous species in a stream receiving treated municipal sewage[J]. Chemical Geology, 2011, 283(1-2): 44-55. |
| [19] |
原雅琼.水生光合生物对漓江流域水化学和岩溶碳汇的影响[D].重庆: 西南大学, 2016. Yuan Y Q. Impacts of aquatic organisms on hydrochemical characteristics and karst carbon sink in Lijiang basin[D]. Chongqing: Southwest University, 2016. http://cdmd.cnki.com.cn/Article/CDMD-10635-1016767037.htm |
| [20] |
吴沿友, 邢德科, 刘莹. 植物利用碳酸氢根离子的特征分析[J]. 地球与环境, 2011, 39(2): 273-277. Wu Y Y, Xing D K, Liu Y. The characteristics of bicarbonate used by plants[J]. Earth and Environment, 2011, 39(2): 273-277. |
| [21] | Han G L, Tang Y, Wu Q X. Hydrogeochemistry and dissolved inorganic carbon isotopic composition on karst groundwater in Maolan, southwest China[J]. Environmental Earth Sciences, 2010, 60(4): 893-899. DOI:10.1007/s12665-009-0226-y |
| [22] | Aravena R, Schiff S L, Trumbore S E, et al. Evaluating dissolved inorganic carbon cycling in a forested lake watershed using carbon isotopes[J]. Radiocarbon, 1992, 34(3): 636-645. DOI:10.1017/S003382220006392X |
| [23] | De Montety V, Martin J B, Cohen M J, et al. Influence of diel biogeochemical cycles on carbonate equilibrium in a karst river[J]. Chemical Geology, 2011, 283(1-2): 31-43. |
| [24] | Tobias C, Böhlke J K. Biological and geochemical controls on diel dissolved inorganic carbon cycling in a low-order agricultural stream:implications for reach scales and beyond[J]. Chemical Geology, 2011, 283(1-2): 18-30. |
| [25] | Falkowski P G, Raven J A. Aquatic photosynthesis[M]. Princeton: Princeton University Press, 2007. |
| [26] | Pawellek F, Veizer J. Carbon cycle in the upper Danube and its tributaries:δ13CDIC constraints[J]. Israel Journal of Earth Sciences, 1994, 43(3): 187-194. |
| [27] |
贾国东, 陈法锦, 邓文锋. 北江河水溶解无机碳同位素的季节变化[J]. 地球科学-中国地质大学学报, 2012, 37(2): 365-369. Jia G D, Chen F J, Deng W F. Seasonal variations of dissolved inorganic carbon isotope in the Beijiang river[J]. Earth Science-Journal of China University of Geosciences, 2012, 37(2): 365-369. |
| [28] | Blum J D, Gazis C A, Jacobson A D, et al. Carbonate versus silicate weathering in the Raikhot watershed within the High Himalayan Crystalline series[J]. Geology, 1998, 26(5): 411-414. DOI:10.1130/0091-7613(1998)026<0411:CVSWIT>2.3.CO;2 |
| [29] | Horton T W, Chamberlain C P, Fantle M, et al. Chemical weathering and lithologic controls of water chemistry in a high-elevation river system:Clark's Fork of the Yellowstone River, Wyoming and Montana[J]. Water Resources Research, 1999, 35(5): 1643-1655. DOI:10.1029/1998WR900103 |
| [30] | Barth J A C, Cronin A A, Dunlop J, et al. Influence of carbonates on the riverine carbon cycle in an anthropogenically dominated catchment basin:evidence from major elements and stable carbon isotopes in the Lagan River (N. Ireland)[J]. Chemical Geology, 2003, 200(3-4): 203-216. DOI:10.1016/S0009-2541(03)00193-1 |
| [31] |
于正良, 杨平恒, 赵瑞一, 等. 春季生物作用对山地岩溶池水地球化学特征的影响[J]. 环境科学, 2015, 36(4): 1263-1269. Yu Z L, Yang P H, Zhao R Y, et al. Influences of biological processes on geochemical characteristics:an example of a mountain karst pool in spring season[J]. Environmental Science, 2015, 36(4): 1263-1269. |
| [32] |
莫雪, 蒲俊兵, 袁道先, 等. 亚热带典型岩溶区地表溪流溶解无机碳昼夜变化特征及其影响因素[J]. 第四纪研究, 2014, 34(4): 873-880. Mo X, Pu J B, Yuan D X, et al. Diel variation and influence factors of dissolved inorganic carbon in a surface creek fed by a karst subterranean stream in subtropical area, SW China[J]. Quaternary Sciences, 2014, 34(4): 873-880. DOI:10.3969/j.issn.1001-7410.2014.04.20 |
| [33] |
吕保樱, 刘再华, 廖长君, 等. 水生植物对岩溶水化学日变化的影响——以桂林岩溶水文地质试验场为例[J]. 中国岩溶, 2006, 25(4): 335-340. Lu B Y, Liu Z H, Liao C J, et al. The influence of aquatic plants on diurnal variations of hydrochemistry in karst system-a case in the Guilin karst experimental site[J]. Carsologica Sinica, 2006, 25(4): 335-340. DOI:10.3969/j.issn.1001-4810.2006.04.014 |
| [34] |
袁道先. "岩溶作用与碳循环"研究进展[J]. 地球科学进展, 1999, 14(5): 425-432. Yuan D X. Progress in the study on karst processes and carbon cycle[J]. Advance in Earth Sciences, 1999, 14(5): 425-432. DOI:10.3321/j.issn:1001-8166.1999.05.001 |
| [35] | Liu Z, Zhao J. Contribution of carbonate rock weathering to the atmospheric CO2 sink[J]. Environmental Geology, 2000, 39(9): 1053-1058. DOI:10.1007/s002549900072 |
| [36] | Liu Z H, Dreybrodt W, Wang H J. A new direction in effective accounting for the atmospheric CO2 budget:considering the combined action of carbonate dissolution, the global water cycle and photosynthetic uptake of DIC by aquatic organisms[J]. Earth-Science Reviews, 2010, 99(3-4): 162-172. DOI:10.1016/j.earscirev.2010.03.001 |
| [37] | Liu Z H, Dreybrodt W. Significance of the carbon sink produced by H2O-carbonate-CO2-aquatic phototroph interaction on land[J]. Science Bulletin, 2015, 60(2): 182-191. DOI:10.1007/s11434-014-0682-y |
| [38] |
李亮, 胡刚, 曹建华, 等. 潮田河典型河流断面水化学日变化及光合作用固定HCO3-碳量估算[J]. 工业安全与环保, 2016, 42(9): 86-90. Li L, Hu G, Cao J H, et al. Hydrochemical daily variations of the typical river section in Chaotian river and the carbon sequestration estimation of HCO3- on photosynthesis[J]. Industrial Safety and Environmental Protection, 2016, 42(9): 86-90. DOI:10.3969/j.issn.1001-425X.2016.09.027 |
| [39] |
王培, 曹建华, 邵景力. 典型水生植物对岩溶水生生态系统无机碳稳定性影响研究[J]. 地球学报, 2017, 38(S1): 51-54. Wang P, Cao J H, Shao J L, et al. Effects of typical aquatic plants on the stability of inorganic carbon in karst aquatic ecosystem[J]. Acta Geoscientica Sinica, 2017, 38(S1): 51-54. DOI:10.3975/cagsb.2017.s1.14 |
| [40] | Odum H T. Primary production in flowing waters[J]. Limnology and Oceanography, 1956, 1(2): 102-117. DOI:10.4319/lo.1956.1.2.0102 |
| [41] | Caffrey J M. Production, respiration and net ecosystem metabolism in U.S. estuaries[J]. Environmental Monitoring and Assessment, 2003, 81(1-3): 207-219. |
| [42] | Marcarelli A M, Van Kirk R W, Baxter C V. Predicting effects of hydrologic alteration and climate change on ecosystem metabolism in a western U.S. river[J]. Ecological Applications, 2010, 20(8): 2081-2088. DOI:10.1890/09-2364.1 |
| [43] | Demars B O L, Thompson J, Manson J R. Stream metabolism and the open diel oxygen method:principles, practice, and perspectives[J]. Limnology and Oceanography Methods, 2015, 13(7): 356-374. DOI:10.1002/lom3.10030 |
| [44] | Pu J B, Li J H, Khadka M B, et al. In-stream metabolism and atmospheric carbon sequestration in a groundwater-fed karst stream[J]. Science of the Total Environment, 2017, 579: 1343-1355. DOI:10.1016/j.scitotenv.2016.11.132 |