 2019, Vol. 40
2019, Vol. 40



汞是一种具有很强的毒性、持久性和生物富集性并且能够在大气中长距离传输的全球性污染物.自工业革命以来, 人为活动(如化石燃料燃烧、有色金属冶炼等)加剧了全球范围内的汞污染[1].湿地是通常位于水陆交错带的一种独特的生态系统, 与森林、海洋并称为全球三大生态系统.高原湿地占我国湿地总面积的1/4, 主要分布在青藏高原、云贵高原和蒙新高原等高原区, 我国高原湿地发育典型、类型多样、功能强大, 具有无可替代的环境生态功能, 是高原区域维持生物多样性的关键地区, 也是影响及响应全球气候变化的敏感区之一.有研究表明, 湿地生态系统是汞的“活性库”和“汇”[2], 湿地中含有丰富的能与汞生成络合物的可溶性碳和腐殖酸, 可以同时吸收富集自然源和人为活动排放的汞, 沉积物中的微生物(硫酸盐还原菌、产甲烷菌、铁还原菌等)可将无机汞转化成甲基汞(MeHg), 在湿地生态系统中经食物链逐级放大, 并最终通过食用鱼类和其他水产品等对人体健康造成威胁.因此有必要针对湿地生态系统食物链中的汞污染特征进行研究.
Hg在自然界存在7种稳定同位素, 分别是196Hg、198Hg、199Hg、200Hg、201Hg、202Hg和204Hg, 同位素之间的相对质量差可达4%.本世纪初, Lauretta等[3]采用多接收体电感耦合等离子体质谱(MC-ICP-MS)法实现了汞同位素组成的精确测定, 为开展汞同位素的研究工作奠定了基础.研究表明, 自然样品(水生系统、煤炭、土壤和微生物等)中不仅存在汞同位素的质量分馏(MDF), 更存在着汞同位素的非质量分馏(MIF), 且汞是自然界少数存在同位素非质量分馏的金属元素之一.汞同位素的MIF对识别某些地球化学过程(光还原作用、挥发作用等)具有重要作用, 可以为湿地生态系统中汞的地球化学循环提供更加可靠的“源”信息.这展示了Hg同位素技术在示踪环境中汞来源和地球化学行为上的广阔应用前景.如Feng等[4]通过测定贵州省不同汞污染区表层土壤的汞同位素组成, 发现不同汞污染区土壤汞存在不同的汞同位素组成特征. Sherman等[5]分析了美国佛罗里达一个大型燃煤企业顺风区沉降汞中的同位素组成, 发现湿沉降汞的同位素组成来自于这一燃煤企业燃烧的煤炭中汞的同位素分馏.因此汞同位素技术为示踪水生生态结构、生物体汞暴露途径等提供了强有力的工具.
贵州威宁草海湿地是在碳酸盐形成的喀斯特盆地上积水发育而成的高原湿地生态系统.由于其水生动植物种类丰富, 水生生物群落生产力较高, 系统功能和结构较完整, 并且是以黑颈鹤(Grus nigricollis)为代表的珍稀鸟类的主要越冬地之一, 因此成为我国亚热带地区湿地生态系统研究的重要基地.草海曾遭受到大量生活生产废水和周围铅锌冶炼厂的污染[6~8], 重金属进入水体后一般都沉积于底泥中, 因此湖泊沉积物是水生生态系统中汞的重要汇和二次排放源, 而底栖动物充当了汞从水体和沉积物进入食物链的重要通道, 使得汞(尤其是甲基汞)随食物链的累积与放大最终造成人体的汞暴露, 且草海湿地生物链复杂, 具有易于汞甲基化及生物富集的生态系统特征.本研究测定了草海食物链中各营养级生物样品的汞同位素组成, 分析了草海食物链中汞同位素的分馏特征, 以期为我国了解西南高原湿地汞的生物地球化学循环过程提供基础数据及理论依据.
1 材料与方法 1.1 样品采集分别于2014年8月、2015年的1月和5月, 在草海湿地采集了沉积物和不同种类生物样品, 包括水生植物、浮游动物、底栖动物、鱼类和鸟类羽毛.样品包含了处于不同营养级的食物链样品, 基本涵盖了草海水生生态系统中的常见水生生物.其中沉积物、水生植物、浮游动物样品带回实验室后, 置于真空冷冻干燥机(FDU-1110型, TOKYO RIKAKIKAI CO. LTD)内, 在-40℃条件下将样品进行低温干燥, 然后用研钵研磨并过100目筛, 保存待测.底栖动物以及鱼样均用去离子水洗净, 自然风干, 再次用去离子水洗净风干, 每个样品分为两部分, 一部分用三层自封袋密封编号后于-20℃冷冻保存, 用于测定总汞及甲基汞含量, 另外选取部分样品用冷冻干燥机在-40℃条件下冷冻干燥, 保证水分完全被去除, 再用碎样机磨碎, 自封袋装样保存用于碳、氮及汞同位素测定.鸟类羽毛带回实验室后, 用去离子水洗净风干后, 再次用去离子水洗净, 风干或低温烘干后, 用磨样机将羽毛粉碎装袋待测.
1.2 样品分析水生植物总汞:植物总汞含量测定参照硝酸水浴消解, 两次金汞齐富集结合冷原子荧光光谱法测定[9].
底栖动物、鱼类、浮游动物、鸟类羽毛总汞:动物总汞测定参照二酸(H2SO4:HNO3=3:7)消解, 两次金汞齐富集结合冷原子荧光光谱法测定[10].
沉积物总汞:参照王水水浴消解, 两次金汞齐富集结合冷原子荧光光谱法测定[11].
水生植物甲基汞测定:采用溶剂萃取-水相乙基化衍生, GC-CVAFS联用的方法测定[12].
底栖动物、鱼类、浮游动物、鸟类羽毛甲基汞测定:采用碱消解结合GC-CVAFS法测定[13].
沉积物甲基汞测定:参照萃取-乙基化结合GC-CVAFS法测定[14].
以上各方法消解的样品均使用CVAFS汞分析仪进行分析测定(MODEL Ⅲ型, U.S.A Brooks Rand LLC).
碳、氮同位素测定分析方法:本研究中样品碳、氮同位素送广东深圳微泰克有限公司测定, 测定同位素采用Carlo Erba EA-1110元素分析仪与Delta PlusFinnigan(Thermo Scient, Waltham, Massachusetts, USA)同位素比率质谱联用仪.分析结果表示为:

|
(1) |
式中, R代表13C或15N, X代表13C/12C或15N/14N.
汞同位素组成分析:在中国科学院地球化学研究所环境地球化学国家重点实验室完成, 所用仪器为英国Nu Instruments公司Nu plasma型MC-ICP-MS.使用连续流进样系统[15]对所有溶液样品进行同位素组成测定.同位素测定使用样品-标样交叉测定法(bracketing method), 并加入Tl标准溶液(NIST SRM 997)作为内标校正仪器测试过程中产生的质量歧视效应, 另外采用2%~3%的HNO3溶液清洗进样系统, 以消除记忆效应.根据已测THg浓度和稀释体积计算汞同位素所需称样量, 由于生物样品中THg浓度相对沉积物较低, 因此以最终稀释后浓度在1μg·mL-1, 部分低含量样品稀释到0.5 μg·mL-1左右, 且酸度 < 15%为原则.
汞同位素质量分馏(MDF, 用δxxxHg表示)和非质量分馏(MIF, 用ΔxxxHg表示)的计算公式如下[16]:
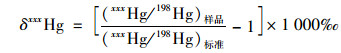
|
(2) |
式中, xxx分别指199、200、201、202、204, (xxxHg/198Hg)标准指的是汞标准溶液NIST SRM 3133汞同位素组成.
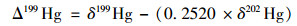
|
(3) |
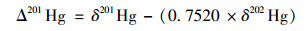
|
(4) |
总汞(THg)和甲基汞(MeHg)分析过程中, 样品处理与分析质量采用试剂空白、平行重复样和标准参考样品(总汞GSS-5、甲基汞ERM-CC580、土壤样品, TORT-2、生物样品, GSB-26、植物样品)进行控制.标准工作曲线线性回归系数R>0.998 4, 标准样品总汞和甲基汞测定次数分别为6次, 测定结果回收率([测定值/标准参考值]×100%)为90%~110%, 平行样品的相对偏差小于10%, 实验结果可靠.
汞同位素组成测定过程中, 利用标准物质BCR482(地衣)、GSS-5(土壤)和GSS-4(土壤)作为第二标准样品进行质量控制, 所测结果与报道的同位素结果差别很小(表 1), 同时对标准物质NIST SRM 3133进行测定, 当连续两次测定值的各个同位素的比值小于±0.10‰时可以接受, 在分析每一个样品前后, 分别测定汞同位素标准NIST SRM 3133, 测试结果:δ202Hg=-0.01±0.05‰, Δ199Hg=0.00±0.03‰, Δ201Hg=0.01±0.05‰(n=30), 说明测试结果可靠.
|
|
表 1 标准参考物质分析结果/‰ Table 1 Results for standard reference materials/‰ |
1.4 数据处理
本研究中, 数据均使用Microsoft Excel 2016和SPSS11.0统计软件进行数据分析, 使用Origin 8.6进行相关图件的绘制.
2 结果与分析 2.1 草海湿地营养级关系和食物网结构很多研究表明, δ15N随着食物链营养级的提高而增大[19~21], 为研究草海湿地中水生生物在食物链中的营养级及食物链构成情况, 采用稳定同位素δ13C和δ15N追踪食物源和确定营养级[22].由于初级生产者水生植物和浮游植物的δ15N值无论是在一个系统内还是系统之间都有很大的变化, 利用其做基准值会引起较大的误差[23], 本研究采用初级消费者浮游动物体内的氮同位素值(δ15N=1.08‰)作为基准值, 营养级计算公式如下:
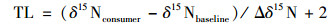
|
(5) |
式中, TL表示所计算生物的营养级, δ15Nconsumer表示消费者的氮同位素值, δ15Nbaseline表示该生态系统中基准线生物中的氮同位素值(1.08‰), Δδ15N表示相邻营养级之间氮同位素的富集程度(3.8‰).而鸟类与水生生物不同, 所以它与被捕食鱼类的δ15N富集值和水生食物网的富集因子不同[24], Mizutani等[25]经过23 a观察得出鸟捕食鱼类的δ15N富集因子为2.4‰.因此可以得出鸟类捕食生物的营养等级关系公式:
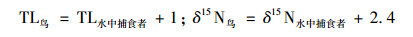
|
(6) |
故鸟类δ15N与营养等级关系为:
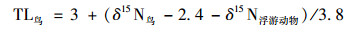
|
(7) |
选取的水生生物包括水生植物、浮游动物、底栖动物、鱼类和鸟类, 基本涵盖了草海湿地中的所有类别水生生物, 可反映出草海湿地生态系统中的汞沿食物链的富集情况, 如表 2所示.
|
|
表 2 草海食物链中碳和氮同位素测定结果 Table 2 Results for δ15N and δ13C of the food web at the Caohai Lake |
由表 2可知, 所有食物链样品δ13C和δ15N的范围分别为-27.40‰~-13.25‰和-1.88‰~15.14‰, 这与孟梅[26]对渤海食物链中的研究结果相一致.与其他种类相比, 白骨顶、蜻蜓幼虫的δ15N值(15.14‰、13.55‰)最大, 营养级也最高(6.07、5.28), 处于食物链的上层; 而水生植物穗状狐尾藻的δ15N值最小(-1.88‰), 处于食物链的底层.由于本文中的大部分鸟类和鱼类都属于杂食性物种, 在捕食过程既会捕食浮游动物, 也捕食浮游植物, 故其通过氮同位素计算出的营养级别相当; 而蜻蜓幼虫是以捕食水生昆虫为主的肉食性动物, 所以其营养级比鱼类和部分鸟类更高.
2.2 草海食物链中汞同位素组成特征表 3列出了草海沉积物样品、食物链不同种群生物中总汞(THg)、甲基汞(MeHg)、甲基汞比例(%MeHg)和汞同位素比值.可以看出, 所有样品的总汞含量范围在6.71~2 733.41 ng·g-1之间, 甲基汞含量范围在0.08~112.63 ng·g-1之间, 可见食物链中汞含量的变化范围较大.从表 3可看出, 所有食物链样品均发生了汞同位素质量分馏(MDF)和非质量分馏(MIF).所有样品中δ202Hg的范围在-2.26‰~-0.26‰之间, Δ199Hg的范围在0.03‰~1.52‰之间, Δ201Hg的范围在0.09‰~1.54‰之间, 表明草海食物链中汞同位素组成的变化范围相对较小, δ202Hg的变化幅度为2‰.低于孟梅对渤海食物链中δ202Hg的研究结果(5.1‰)[26], 也低于Perrot等[27]对俄罗斯贝加尔湖食物链中δ202Hg测定的变化幅度(3.5‰), 这很可能与草海食物链中整体汞污染水平较低, 总汞和甲基汞含量相对渤海和贝加尔湖较低有关, 且草海位于高海拔地区, 强烈的紫外辐射引起的去甲基化作用会导致甲基汞和甲基汞比例降低, 使得草海食物链中δ202Hg的变化幅度低于已有的研究结果..
|
|
表 3 草海食物链样品中总汞含量、甲基汞含量、甲基汞百分比、汞同位素测定结果 Table 3 Results for the total mercury (THg), methylmercury (MeHg), methylmercury ratios (%MeHg), and Hg isotopic compositions of the food web at the Caohai Lake |
所有样品均表现出偏负的δ202Hg(-0.93‰±1.32‰, n=14)以及偏正的Δ199Hg(0.79‰±0.82‰, n=14), 见图 1, 其中浮游动物表现出最低的δ202Hg值(-2.26‰), 这与已有的研究结果一致, 如Perrot等[27]报道的俄罗斯贝加尔湖食物链中浮游生物具有最低的δ202Hg(-1.09‰±0.31‰), 孟梅报道的渤海食物链中浮游植物的δ202Hg也最低(-3.22‰)[26]; 鱼类具有最高的δ202Hg值(-0.26‰)和最高的Δ199Hg值(1.52‰), 且均表现出偏负的δ202Hg和偏正的Δ199Hg, 这与花秀兵对我国西藏地区, 三峡水库, 小浪底水库鱼类的研究结果一致[28], 在其他的研究中也发现绝大多数淡水鱼类均表现出偏负的δ202Hg和偏正的Δ199Hg[29, 30].不同的栖息环境会影响水生生物体内的汞同位素分馏特征:Jackson等的[31]研究发现食性相同的鱼类体内的汞同位素特征随着生活环境(深处温度低的冷环境和表面温度高的环境)不同而变化; Gantner等[30]的研究表明鱼体内的MDF与纬度存在着负相关的关系.

|
图 1 草海食物链不同样品的汞同位素组成 Fig. 1 Hg isotopic compositions of different samples of the food web at the Caohai Lake |
由表 2可知, 草海食物链中整体汞污染水平较低, 甲基汞及甲基汞比例(%MeHg范围0.15%~74.59%, 均值11.86%)均低于其他文献报道结果, 其中初级生产者水生植物%MeHg均值为1.13%, 杂食性鸟类羽毛%MeHg均值为2.6%, 鱼类%MeHg均值为18.9%.草海整个食物链较低的甲基汞及甲基汞比例可能与草海处于较高的海拔水平, 紫外线强烈而具有较高的去甲基化作用有关[32].
图 2给出了草海食物链中各生物种群的总汞、甲基汞含量, 可以看出, 能量从初级生产者水生植物到高级消费者鸟类羽毛传递的同时, 甲基汞含量也在逐级增大, 而总汞含量却没有逐级增大的趋势, 这可能与不同生物具有不同的汞富集途径有关.底栖动物和植物除了通过水或其它摄食途径富集汞以外, 还从沉积物吸收富集了大量无机汞, 故具有较高的总汞含量.从图 3也可以看出, δ15N与%MeHg之间虽然没有显著相关性(r=0.45, P > 0.05), 但也表现出%MeHg随着δ15N值升高而增大的趋势, 这也表明甲基汞能通过水生食物链进行富集和生物放大, 进而威胁到食物网较为上层的消费者.

|
图 2 草海各生物种群总汞、甲基汞含量 Fig. 2 Concentration of THg and MeHg in different aquatic organisms in Caohai |

|
图 3 δ15N与%MeHg的关系 Fig. 3 Relationship between %MeHg and δ15N |
已有研究表明:在实验室的喂养实验中, 被喂养鱼类的同位素组成接近食物的同位素组成, 所以鱼类在营养转移的过程中并不会发生MDF和MIF[33, 34].但也有研究表明:δ15N与鱼体的质量分馏(MDF)存在着正相关的关系[31]; 花秀兵等[35]的研究指出我国小浪底水库中鱼体甲基汞在营养转移的过程中可能发生了MDF, 从而导致MDF随着δ15N的升高而增加; Perrot等[27]的研究发现贝加尔湖食物链中δ202Hg与δ15N之间有显著相关性, 且质量分馏(MDF)主要与食物链中的营养传递过程有关.为研究草海湿地食物链中汞同位素质量分馏与营养级之间的关系, 图 4给出了δ15N和δ202Hg之间的相关性, 可见除穗状狐尾藻(δ15N=-1.88‰)以外, δ15N和δ202Hg之间表现出显著正相关性(r=0.58, P < 0.05), 表明汞在食物链的生物富集过程中相对富集较重的汞同位素, 这与已有的研究结果一致[27, 36].从图 4也可以看出, δ202Hg和营养级之间虽然没有相关性, 但也表现出类似的趋势(r=0.59, P=0.057).从图 5可以看出, 草海食物链中各样品甲基汞比例(%MeHg)与δ202Hg之间并不存在显著相关性(r=0.1, P=0.73), 表明草海食物链中汞同位素质量分馏(MDF)差异不是来自于水体甲基汞富集程度差异, 而很可能是甲基汞在营养传递过程中产生了质量分馏.

|
图 4 δ202Hg与δ15N和营养级的关系 Fig. 4 Relationship between δ15N and the trophic level and δ202Hg |

|
图 5 甲基汞比例(%MeHg)与δ202Hg的关系 Fig. 5 Relationship between%MeHg and δ202Hg |
从表 3可以看出, 所有的食物链样品都呈现出不同程度的偏正的非质量分馏特征, Δ199Hg的范围在0.03‰~1.52‰之间, 平均值为0.79‰, Δ201Hg的范围在0.09‰~1.54‰之间, 平均值为0.74‰.浮游动物不仅具有最低的δ202Hg值(-2.26‰), 还具有最低的Δ199 Hg值(0.03‰).关于水生生物富集奇数汞同位素的现象已经大量报道, Δ199Hg变化达7‰[29, 30, 32, 36~39].然而有研究表明, 在水生环境中, 水柱和表层沉积物中的生物过程(包括生物甲基化, 去甲基化和还原过程)不太可能产生汞同位素的非质量分馏[40~42], 光化学反应才是导致全球汞同位素非质量分馏的主要原因[43~46], 水体中汞同位素的非质量分馏主要来自于水柱中无机汞的光致还原作用和甲基汞的光致降解过程, 并且这些过程会引起剩余的汞反应化合物中奇数汞同位素的富集[47, 48], 从而导致水生生物存在明显的奇数汞同位素异常的现象. Bergquist等[36]的研究发现, 可以用Δ199Hg/Δ201Hg的比值来判断Hg2+的光化学还原过程(Δ199Hg/Δ201Hg≈1.0)和甲基汞的光降解过程(Δ199Hg/Δ201Hg≈1.36).从图 6可以看出, 草海食物链中Δ199Hg与Δ201Hg线性拟合方程为y=1.1177x-0.0238, 呈现出Δ199Hg/Δ201Hg的比值为1.1177, 可见大部分样品的Δ199Hg与Δ201Hg都位于y=1.36x和y=1.0x这两条直线之间, 表明草海食物链中的汞同位素非质量分馏可能同时受到甲基汞的光致去甲基化过程和无机汞的光致还原过程的影响.

|
图 6 草海食物链中的汞同位素非质量分馏特征 Fig. 6 Characteristics of the Hg isotopic mass-independent fractionation of the food web at the Caohai Lake |
草海湿地食物链中汞同位素非质量分馏与营养级之间的关系如图 7所示, 可见Δ199Hg和δ15N之间表现出显著正相关性(r=0.67, P < 0.05), 表明非质量分馏(MIF)可能与食物链的营养传递过程有关, 也可能与水体中非质量分馏过程产生的富集奇汞同位素的汞反应物化合物(如甲基汞)在生物体中的不同富集程度有关; 如前所述, 很多研究已经证明, 在水生环境中, 水柱和表层沉积物中的生物过程(包括生物甲基化, 去甲基化和还原过程)不太可能产生汞同位素的非质量分馏, 光化学反应才是导致全球汞同位素非质量分馏的主要原因.鱼类在营养转移的过程中也并不会发生非质量分馏[33, 34].从图 8可以看出, 草海食物链中Δ199Hg与甲基汞比例(%MeHg)呈显著正相关性(r=0.58, P < 0.05), 表明水生生物体内吸收富集的MeHg含量越多, 无机汞含量越少, 则其发生汞同位素非质量分馏的程度就会越高, 这与Li等[49]对大米中甲基汞的非质量分馏研究结果一致.由此可以推断, 水体中MeHg的光致去甲基化过程可能是导致食物链生物非质量分馏随营养级升高而增大的主要原因.水体中MeHg的光致降解过程产生了非质量分馏, 导致水体中的MeHg富集奇同位素, 而这些富集了奇同位素的甲基汞在生物体中不同程度地累积导致了不同营养级生物具有不同的非质量分馏数值.浮游动物的非质量分馏程度最低(Δ199Hg=0.03‰), 很可能就是因为其体内具有最低的%MeHg(0.15%)而导致, 鱼体中较高程度的非质量分馏(Δ199Hg范围为0.67‰~1.52‰, 均值1.28‰)可能是因为鱼类通过食物链富集了更高比例的甲基汞.随着各级食物链样品中甲基汞比例的提高, 非质量分馏程度也越高, 表明草海食物链中非质量分馏(MIF)主要与水体中甲基汞的光致降解过程和营养传递过程中甲基汞在生物体中的积累程度有关.

|
图 7 Δ199Hg和δ15N的关系 Fig. 7 Relationship between Δ199Hg and δ15N |

|
图 8 Δ199Hg和甲基汞比例(%MeHg)的关系 Fig. 8 Relationship between Δ199Hg and %MeHg |
(1) 草海水生生态系统中的汞和甲基汞在食物链中的流动会随着营养级别的增高而被富集放大.
(2) 草海湿地食物链中所有样品都具有明显的质量分馏(MDF)与非质量分馏特征(MIF), δ202Hg的范围在-2.26‰~-0.26‰之间, 平均值为-0.93‰; Δ199Hg的范围在0.03‰~1.52‰之间, 平均值为0.79‰, Δ201Hg的范围在0.09‰~1.54‰之间, 平均值为0.74‰.
(3) 除穗状狐尾藻(δ15N=-1.88‰)以外, δ15N和δ202Hg之间表现出显著正相关性(r=0.58, P < 0.05), 表明汞在食物链的生物富集过程中相对富集偏重的汞同位素, 且质量分馏(MDF)很可能与甲基汞在营养级中的传递过程有关.
(4)Δ199Hg/Δ201Hg≈1.12, 表明草海食物链中的汞同位素非质量分馏可能同时受到甲基汞的光致去甲基化过程和无机汞的光致还原过程的影响. Δ199Hg和δ15N之间表现出显著相关性(r=0.67, P < 0.05), 且Δ199Hg与甲基汞比例(%MeHg)之间也呈显著正相关性(r=0.58, P < 0.05), 表明草海食物链中的汞同位素非质量分馏可能主要来源于水体MeHg的光致去甲基化过程, 汞非质量分馏程度随食物链的升高而变大, 是由于营养传递过程中生物体内甲基汞的比例提高所致.
| [1] | Selin N E. Global biogeochemical cycling of mercury:a review[J]. Annual Review of Environment and Resources, 2010, 34: 43-63. |
| [2] | Lindqvist O, Johansson K, Bringmark L, et al. Mercury in the Swedish environment-recent research on causes, consequences and corrective methods[J]. Water, Air, and Soil Pollution, 1991, 55(1-2): 1-261. |
| [3] | Lauretta D S, Klaue B, Blum J D, et al. Mercury abundances and isotopic compositions in the Murchison (CM) and Allende (CV) carbonaceous chondrites[J]. Geochimica et Cosmochimica Acta, 2001, 65(16): 2807-2818. DOI:10.1016/S0016-7037(01)00630-5 |
| [4] | Feng X B, Yin R S, Yu B, et al. Mercury isotope variations in surface soils in different contaminated areas in Guizhou Province, China[J]. Chinese Science Bulletin, 2013, 58(2): 249-255. DOI:10.1007/s11434-012-5488-1 |
| [5] | Sherman L S, Blum J D, Keeler G J, et al. Investigation of local mercury deposition from a coal-fired power plant using mercury isotopes[J]. Environmental Science & Technology, 2012, 46(1): 382-390. |
| [6] |
张建利, 吴华, 喻理飞, 等. 贵州草海湿地流域典型喀斯特森林植物群落结构特征[J]. 南方农业学报, 2013, 44(3): 471-477. Zhang J L, Wu H, Yu L F, et al. Structure feature of typical Karst forest plant community in Caohai wetland watershed of Guizhou Province[J]. Journal of Southern Agriculture, 2013, 44(3): 471-477. DOI:10.3969/j:issn.2095-1191.2013.3.471 |
| [7] | 李广辉.贵州省土法炼锌中环境汞污染研究[D].重庆: 西南农业大学, 2004. http://cdmd.cnki.com.cn/Article/CDMD-10625-2004077626.htm |
| [8] | 张槐安, 雷吉华.贵州草海湿地保护措施研究[C].见: 2013中国环境科学学会学术年会论文集(第六卷).昆明: 中国环境科学学会, 2013. |
| [9] | 郑伟, 冯新斌, 李广辉, 等. 硝酸水浴消解-冷原子荧光光谱法测定植物中的总汞[J]. 矿物岩石地球化学通报, 2006, 25(3): 285-287. DOI:10.3969/j.issn.1007-2802.2006.03.012 |
| [10] |
阎海鱼, 冯新斌, 李仲根, 等. 半封闭溶样冷原子荧光测定鱼体中总汞分析方法的建立[J]. 地球与环境, 2005, 33(1): 89-92. Yan H Y, Feng X B, Li Z G, et al. A methodological development in measuring total mercury in fish using semi-closed digestion and CVAFS[J]. Earth and Environment, 2005, 33(1): 89-92. |
| [11] | 李仲根, 冯新斌, 何天容, 等. 王水水浴消解-冷原子荧光法测定土壤和沉积物中的总汞[J]. 矿物岩石地球化学通报, 2005, 24(2): 140-143. DOI:10.3969/j.issn.1007-2802.2005.02.009 |
| [12] |
仇广乐, 冯新斌, 梁琏, 等. 溶剂萃取-水相乙基化衍生GC-CVAFS联用测定苔藓样品中的甲基汞[J]. 分析测试学报, 2005, 24(1): 29-32. Qiu G L, Feng X B, Liang L, et al. Determination of methylmercury in moss by ethylation-gas chromatography-cold vapor atomic fluorescence spectrometry with solvent extraction[J]. Journal of Instrumental Analysis, 2005, 24(1): 29-32. DOI:10.3969/j.issn.1004-4957.2005.01.007 |
| [13] |
阎海鱼, 冯新斌, Liang L, 等. GC-CVAFS法测定鱼体内甲基汞的分析方法研究[J]. 分析测试学报, 2005, 24(6): 78-80. Yan H Y, Feng X B, Liang L, et al. Determination of methyl mercury in fish using GC-CVAFS[J]. Journal of Instrumental Analysis, 2005, 24(6): 78-80. DOI:10.3969/j.issn.1004-4957.2005.06.019 |
| [14] | 何天容, 冯新斌, 戴前进, 等. 萃取-乙基化结合GC-CVAFS法测定沉积物及土壤中的甲基汞[J]. 地球与环境, 2004, 32(2): 83-86. |
| [15] | Yin R S, Feng X B, Foucher D, et al. High precision determination of mercury isotope ratios using online mercury vapor generation system coupled with multicollector Inductively coupled plasma-mass spectrometer[J]. Chinese Journal of Analytical Chemistry, 2010, 38(7): 929-934. DOI:10.1016/S1872-2040(09)60055-4 |
| [16] |
冯新斌, 尹润生, 俞奔, 等. 汞同位素地球化学概述[J]. 地学前缘, 2015, 22(5): 124-135. Feng X B, Yin R S, Yu B, et al. A review of Hg isotope geochemistry[J]. Earth Science Frontiers, 2015, 22(5): 124-135. |
| [17] | Estrade N, Carignan J, Sonke J E, et al. Measuring Hg isotopes in bio-geo-environmental reference materials[J]. Geostandards and Geoanalytical Research, 2010, 34(1): 79-93. DOI:10.1111/ggr.2010.34.issue-1 |
| [18] | Huang Q, Liu Y L, Chen J B, et al. An improved dual-stage protocol to pre-concentrate mercury from airborne particles for precise isotopic measurement[J]. Journal of Analytical Atomic Spectrometry, 2015, 30(4): 957-966. DOI:10.1039/C4JA00438H |
| [19] |
李云凯, 贡艺. 基于碳、氮稳定同位素技术的东太湖水生食物网结构[J]. 生态学杂志, 2014, 33(6): 1534-1538. Li Y K, Gong Y. Food web structure of the East Lake Taihu by analysis of stable carbon and nitrogen isotopes[J]. Chinese Journal of Ecology, 2014, 33(6): 1534-1538. |
| [20] |
易现峰, 张晓爱. 稳定性同位素技术在生态学上的应用[J]. 生态学杂志, 2005, 24(3): 306-314. Yi X F, Zhang X A. Application of stable isotopic approach in ecology:a review[J]. Chinese Journal of Ecology, 2005, 24(3): 306-314. DOI:10.3321/j.issn:1000-4890.2005.03.016 |
| [21] | 万袆, 胡建英, 安立会, 等. 利用稳定氮和碳同位素分析渤海湾食物网主要生物种的营养层次[J]. 科学通报, 2005, 50(7): 708-712. DOI:10.3321/j.issn:0023-074X.2005.07.016 |
| [22] | Jardine T D, Kidd K A, Fisk A T. Applications, considerations, and sources of uncertainty when using stable isotope analysis in ecotoxicology[J]. Environmental Science & Technology, 2006, 40(24): 7501-7511. |
| [23] | 蔡德陵, 李红燕, 唐启升, 等. 黄东海生态系统食物网连续营养谱的建立:来自碳氮稳定同位素方法的结果[J]. 中国科学C辑生命科学, 2005, 35(2): 123-130. |
| [24] | Hobson K A, Welch H E. Determination of trophic relationships within a high Arctic marine food web using δ13C and δ15N analysis[J]. Marine Ecology Progress Series, 1992, 84: 9-18. DOI:10.3354/meps084009 |
| [25] | Mizutani H, Kabaya Y, Wada E. Nitrogen and carbon isotope compositions relate linearly in Cormorant tissues and its diet[J]. Isotopenpraxis Isotopes in Environmental and Health Studies, 1991, 27(4): 166-168. DOI:10.1080/10256019108622500 |
| [26] | 孟梅.区域环境中汞的污染特征及迁移转化过程研究[D].北京: 中国科学院大学, 2015. http://www.wanfangdata.com.cn/details/detail.do?_type=degree&id=Y2852075 |
| [27] | Perrot V, Pastukhov M V, Epov V N, et al. Higher mass-independent isotope fractionation of methylmercury in the pelagic food web of Lake Baikal (Russia)[J]. Environmental Science & Technology, 2012, 46(11): 5902-5911. |
| [28] | 花秀兵.汞稳定同位素分析方法的优化和应用[D].焦作: 河南理工大学, 2016. http://cdmd.cnki.com.cn/Article/CDMD-10460-1017811698.htm |
| [29] | Laffont L, Sonke J E, Maurice L, et al. Anomalous mercury isotopic compositions of fish and human hair in the Bolivian Amazon[J]. Environmental Science & Technology, 2009, 43(23): 8985-8990. |
| [30] | Gantner N, Hintelmann H, Zheng W, et al. Variations in stable isotope fractionation of Hg in food webs of Arctic lakes[J]. Environmental Science & Technology, 2009, 43(24): 9148-9154. |
| [31] | Jackson T A, Whittle D M, Evans M S, et al. Evidence for mass-independent and mass-dependent fractionation of the stable isotopes of mercury by natural processes in aquatic ecosystems[J]. Applied Geochemistry, 2008, 23(3): 547-571. DOI:10.1016/j.apgeochem.2007.12.013 |
| [32] |
孙荣国, 毛雯, 马明, 等. 水体中甲基汞光化学降解特征研究[J]. 环境科学, 2012, 33(12): 4329-4334. Sun R G, Mao W, Ma M, et al. Characteristics of monomethylmercury photodegradation in water body[J]. Environmental Science, 2012, 33(12): 4329-4334. |
| [33] | Kwon S Y, Blum J D, Carvan M J, et al. Absence of fractionation of mercury isotopes during trophic transfer of methylmercury to freshwater fish in captivity[J]. Environmental Science & Technology, 2012, 46(14): 7527-7534. |
| [34] | Kwon S Y, Blum J D, Chirby M A, et al. Application of mercury isotopes for tracing trophic transfer and internal distribution of mercury in marine fish feeding experiments[J]. Environmental Toxicology and Chemistry, 2013, 32(10): 2322-2330. DOI:10.1002/etc.v32.10 |
| [35] |
花秀兵, 毛宇翔, 刘洪伟, 等. 小浪底水库鱼体和沉积物中汞稳定同位素组成特征[J]. 环境化学, 2016, 35(11): 2245-2252. Hua X B, Mao Y X, Liu H W, et al. Characteristics of stable mercury isotopic compositions in fish and sediments from Xiaolangdi Reservoir[J]. Environmental Chemistry, 2016, 35(11): 2245-2252. DOI:10.7524/j.issn.0254-6108.2016.11.2016032201 |
| [36] | Bergquist B A, Blum J D. Mass-dependent and-independent fractionation of Hg isotopes by photoreduction in aquatic systems[J]. Science, 2007, 318(5849): 417-420. DOI:10.1126/science.1148050 |
| [37] | Das R, Salters V J M, Odom A L. A case for in vivo mass-independent fractionation of mercury isotopes in fish[J]. Geochemistry, Geophysics, Geosystems, 2009, 10(11): Q11012. |
| [38] | Senn D B, Chesney E J, Blum J D, et al. Stable isotope (N, C, Hg) study of methylmercury sources and trophic transfer in the northern gulf of Mexico[J]. Environmental Science & Technology, 2010, 44(5): 1630-1637. |
| [39] | Blum J D, Popp B N, Drazen J C, et al. Methylmercury production below the mixed layer in the North Pacific Ocean[J]. Nature Geoscience, 2013, 6(10): 879-884. DOI:10.1038/ngeo1918 |
| [40] | Kritee K, Blum J D, Barkay T. Mercury stable isotope fractionation during reduction of Hg(Ⅱ) by different microbial pathways[J]. Environmental Science & Technology, 2008, 42(24): 9171-9177. |
| [41] | Kritee K, Barkay T, Blum J D. Mass dependent stable isotope fractionation of mercury during mer mediated microbial degradation of monomethylmercury[J]. Geochimica et Cosmochimica Acta, 2009, 73(5): 1285-1296. |
| [42] | Rodríguez-González P, Epov V N, Bridou R, et al. Species-specific stable isotope fractionation of mercury during Hg(Ⅱ) methylation by an anaerobic bacteria (Desulfobulbus propionicus) under dark conditions[J]. Environmental Science & Technology, 2009, 43(24): 9183-9188. |
| [43] | Sun R Y, Sonke J E, Heimbürger L E, et al. Mercury stable isotope signatures of world coal deposits and historical coal combustion emissions[J]. Environmental Science & Technology, 2014, 48(13): 7660-7668. |
| [44] | Sonke J E. A global model of mass independent mercury stable isotope fractionation[J]. Geochimica et Cosmochimica Acta, 2011, 75(16): 4577-4590. DOI:10.1016/j.gca.2011.05.027 |
| [45] | Chen J B, Hintelmann H, Feng X B, et al. Unusual fractionation of both odd and even mercury isotopes in precipitation from Peterborough, ON, Canada[J]. Geochimica et Cosmochimica Acta, 2012, 90: 33-46. DOI:10.1016/j.gca.2012.05.005 |
| [46] | Estrade N, Carignan J, Donard O F X. Isotope tracing of atmospheric mercury sources in an urban area of northeastern France[J]. Environmental Science & Technology, 2010, 44(16): 6062-6067. |
| [47] | Zheng W, Hintelmann H. Isotope fractionation of mercury during its photochemical reduction by low-molecular-weight organic compounds[J]. The Journal of Physical Chemistry A, 2010, 114(12): 4246-4253. DOI:10.1021/jp9111348 |
| [48] | Zheng W, Hintelmann H. Mercury isotope fractionation during photoreduction in natural water is controlled by its Hg/DOC ratio[J]. Geochimica et Cosmochimica Acta, 2009, 73(22): 6704-6715. |
| [49] | Li P, Du B Y, Maurice L, et al. Mercury isotope signatures of methylmercury in rice samples from the Wanshan mercury mining area, China:environmental implications[J]. Environmental Science & Technology, 2017, 51(21): 12321-12328. |