 2019, Vol. 40
2019, Vol. 40



2. 中国烟草总公司重庆市公司, 重庆 400023
2. Chongqing Municipal Tobacco Company, Chongqing 400023, China
氮(N)元素生物地球化学循环是陆地生态系统中最基本的物质循环过程, 在农业生态系统的氮循环中, 氮的投入和损失是氮循环的关键过程.氮素以N2O的形式排放到大气中是氮损失的重要途径, 其中农业生态系统是N2O重要的排放源[1].菜地生态系统是重要的旱地生态系统, 与粮食作物种植系统不同, 菜地种植系统具有化肥施用量大、灌水频繁、复种指数高等特点[2], 易造成土壤微生物(特别是N转化功能微生物)群落结构和数量发生变化[3]、导致土壤N2O大量排放[4]、引发土壤酸化等一系列环境问题[5]. Wang等[4]的研究表明, 我国菜地土壤因施肥产生的N2O直接排放量(以N计)每年高达6.7×107 kg.我国氮肥投入量大, 但利用率却只有30%, 剩余的氮肥将以氨挥发、N2O、硝酸盐等形式进入到环境中, 成为污染源.因此, 在菜地土壤中开展有机物料替代部分化肥研究, 同时掌握菜地土壤N循环相关的微生物的群落结构和丰度, 不仅能提高土壤质量, 而且能改善因氮肥施用带来的生态环境问题(缓解N2O排放).
硝化作用是土壤N转化的重要过程, 含氨单加氧酶基因(amoA)的氨氧化古菌(ammonia-oxidizing archaea, AOA)和氨氧化细菌(ammonia-oxidizing bacteria, AOB)驱动的氨氧化过程是硝化作用的关键步骤, 该过程的中间产物NH2OH会发生化学分解而释放N2O, 同时硝化作物的产物NO3-可以为反硝化微生物所用间接影响N2O的释放[6, 7].硝化作用是旱地土壤N2O排放的主要途径, 土壤pH、水分和温度等性质以及底物和产物通过影响AOA和AOB群落结构和丰度, 进而影响土壤N2O排放[7].与N2O释放相关的微生物种群特征是决定土壤N2O释放的基础[8], AOA是土壤重要的N2O释放来源, 在红壤蔬菜田土壤氨氧化过程中占绝对优势[9].而在极高的氮素投入的设施菜地中, 土壤氨氧化微生物主要以AOB为主[10].由于产生N2O的主要微生物可能是反硝化真菌、AOA或异养氨氧化微生物等, 因此部分研究中未发现相关功能微生物种群特征与N2O释放之间的关系[11, 12].
秸秆是我国重要的农业资源, 将其还田不仅可以提高土壤肥力和改善土壤结构, 还能促进微生物活性, 并能够减缓因焚烧而带来的大气污染[13, 14].秸秆还田对菜地土壤的N2O排放存在重要影响[2], 同时也会影响土壤氨氧化微生物群落.有研究表明, 水稻秸秆还田对氨氧化细菌群落丰度和结构都没有明显的影响[14], 甚至降低了amoA基因多样性指数和均匀度[15].但在酸性水稻土(pH约5.3)中, 水稻秸秆还田可以显著增加AOA种群丰度[16].萨如拉等[17]基于秸秆还田定位试验, 表明玉米秸秆深翻还田能提高土壤AOB amoA基因多样性指数, 且随着还田时间的增加呈增加趋势. AOA在红壤微生物组成和生态系统中具有重要地位[9], AOB是石灰性紫色土硝化作用的主要推动者[18], 可见不同来源的秸秆、土壤类型对土壤氨氧化微生物群落结构和丰度的影响存在不一致性.国内对水稻秸秆还田研究较多, 且以往的研究主要集中探讨了秸秆还田对土壤N2O排放或土壤氨氧化微生物的影响, 关于秸秆还田, 尤其是玉米秸秆与化肥减量配施后, 菜地土壤AOA、AOB微生物群落结构和丰度变化与土壤N2O排放的综合分析研究较少.紫色土为初育岩性土, 在我国农业生产中占有重要地位, 因此本研究以紫色土为研究对象, 采用静态箱-气相色谱法, 原位观测了玉米秸秆与化肥配施下的菜地土壤N2O排放, 并利用克隆技术和实时荧光定量PCR技术, 结合土壤理化性质和氨氧化微生物功能基因丰度, 分析了秸秆与化肥配施对土壤N2O排放及相关微生物的影响, 以期为调控和减缓紫色土N2O排放提供理论依据.
1 材料与方法 1.1 研究概况及试验材料位于重庆市北碚区的“国家紫色土土壤肥力与肥料效益长期监测基地”, 海拔266.3 m, 年平均气温18.3℃.最高和最低气温平均为28. 7℃和7.7℃, ≥10℃, 积温6006℃, 年降雨量1086.6 mm, 年日照时数1276.7 h, 属于亚热带季风气候.试验土壤为侏罗纪沙溪庙组紫色泥页岩发育形成的紫色土, 中性紫色土亚类, 灰棕紫泥土属, 是重庆四川紫色土区分布最广的一种土壤.其基本理化性质为:有机质含量11.12 g·kg-1, 碱解氮83.02 mg·kg-1, 速效磷44.10 mg·kg-1, 速效钾208.75 mg·kg-1, pH为5.8.试验中所用秸秆为玉米秸秆, 取自试验基地农田, 其有机碳含量为338.8 g·kg-1, C/N为33.9, 经晒干后切成5~10 cm左右, 备用.
1.2 试验设计试验设置:① CK, 无物料还田; ② F, 常规化肥处理; ③ 100FS, 秸秆还田配施100%化肥处理; ④ 70FS, 秸秆还田配施70%化肥处理; ⑤ 60FS, 秸秆还田配施60%化肥处理; ⑥50FS, 秸秆还田配施50%化肥处理.每个处理3个重复, 采用随机区组排列在18个2 m×1 m能独立排灌的微区内, 各个微区的水肥管理均相同.
试验于2016年11月开始, 共种植3季蔬菜, 施用两次秸秆, 分别在种植蔬菜(莴笋, 辣椒)前将秸秆均匀撒覆在土壤表面进行翻耕(每次施用量为12000 kg·hm-2), 翻耕深度为30~40 cm, 为保持相对一致性, 对照CK和F处理也进行同样翻耕.各作物的氮、磷、钾肥的常规施用量按文献[19]的化肥施用量, 化肥减量处理在常规使用量的基础上减少相应的百分比, 各季作物栽培方式和灌溉、除草等田间管理按照当地习惯进行, 松根除草的深度为10 cm左右, 灌溉时间通常在施肥和追肥后, 以及土壤含水量较低的时候, 每个处理均进行灌溉.氮、磷、钾肥品种分别为尿素(N46%)、过磷酸钙(P2O512%)、氯化钾(K2O 60%).具体的蔬菜种植及施肥情况详见表 1.在辣椒最后一次收获后(2017年9月14日), 采集0~20 cm表层土壤, 取部分鲜样用于铵态氮、硝态氮、可溶性氮含量和氨氧化微生物(AOA和AOB)群落分析, 剩余土壤自然风干后用于测定土壤理化性质.
|
|
表 1 种植及施肥情况 Table 1 Planting and fertilization |
1.3 试验方法 1.3.1 N2O排放
采用静态箱-气相色谱法测定菜地土壤N2O排放通量, 采样箱材质为不锈钢材料, 由底座和顶箱2部分组成, 底座(横截面为50 cm×50 cm)长期固定于田间, 底壁插入土中约10 cm, 底座顶端有深、宽均为3 cm的密封水槽, 采样前往槽内注水以防底座和顶箱结合处漏气.顶箱(50 cm×50 cm×50 cm)外覆绝热材料以减少采样箱内温度随外界气温升高对试验结果的影响, 同时, 顶箱内部安装2个小风扇, 用于混合箱内气体.顶箱侧面安装电源插头、温度探头(JM624型便携式测温计)和采样管.试验期间, 采样频率为每周1次, 每次控制在当天09:00~11:00间进行, 采样时用60 mL注射器采集第1次样品, 之后每间隔10 min采样1次, 罩箱时间为30 min, 共采集4个气体样品.在气体样品采集的同时, 记下开始和结束时的气温和5 cm深土壤温度, 并采集表层土样, 测定含水量.遇到下雨及施肥情况, 采样频率增加, 为每2 d一次, 持续一周.
1.3.2 土壤氨氧化微生物土壤DNA利用FastDNA®SPIN Kit for soil(MP Biomedicals, LLC)试剂盒, 分别称取0.5 g新鲜土样, 按照试剂盒上的操作步骤进行土壤微生物总DNA的提取.提取后的DNA -20℃存放, 用于AOA和AOB amoA基因拷贝数和克隆测序.
克隆文库:选用不同的引物对土壤AOA和AOB amoA基因进行扩增(引物见表 2).利用DNA纯化回收试剂盒(Universal DNA Purification Kit, TIANGEN)对扩增PCR产物进行纯化, 纯化产物通过pGEM-T载体(Promega Corp, USA)进行克隆.根据“蓝白斑”(抑制剂:0.2% X-Gal, 0.1% Amp, 0.05% IPTG)对克隆子进行初步筛选, 再通过菌液PCR扩增(体系25 μL)电泳筛选后, 选出目的大小片段的克隆, 每个处理选择30个克隆子送至生工生物有限公司进行序列测定.利用Muthur软件将测序成功的结果进行序列分析, 筛选核糖酸序列相似度>97%序列, 将其视为同一个可操作分类单元(operational taxonomic units, OTU).将代表基因序列提交NCBI (National Center for Biotechnology Information)以获取基因登录号. AOA和AOB amoA基因登录号分别为:MH200776-MH200817, MH200739-MH200775.
|
|
表 2 氨氧化细菌和氨氧化古菌扩增引物 Table 2 Primers of ammonia oxidizing bacteria and ammonia-oxidizing bacteria |
采用实时荧光定量PCR法测定AOA和AOB amoA基因拷贝数.分别选取1份质量高的qPCR产物, 利用DNA凝胶纯化试剂盒(Universal DNA Purification Kit, TIANGEN)对扩增的产物进行纯化, 并将纯化后的产物进行克隆.将测序成功的阳性克隆子利用质粒提取试剂盒(TIAN prep Mini Plasmid Kit, TIANGEN)提取克隆子的质粒, 测定其DNA浓度, 并将其以10倍为间隔系列稀释成实时荧光定量PCR测定标准品.将样品与标准品一起进行Real-Time PCR检测, 包括阴性对照在内每个样品设3个重复. AOA和AOB引物分别为amo_F/amo Ra(序列5′-3′:ATGGTCTGGCTAAGACGMTGTA/GCGGCCATCCATCTGTATGT)和amoA_1F/amoA_1R(序列5′-3′同表 1). AOA反应体系:ABI Prower SybrGreen qPCR Master Mix(ABI, USA)12.5 μL, 引物各1 μL, DNA模板1 μL, 加ddH2O至25 μL; 反应程序:预变性95℃ 1 min, 变性94℃ 45 s, 退火55℃ 45 s, 延伸温度72℃ 45 s, 共进行35个循环. AOB反应体系:ABI Prower SybrGreen qPCR Master Mix(ABI, USA)12.5 μL, 引物各0.5 μL, DNA模板2.5 μL, 加ddH2O至25 μL.反应程序:预变性94℃ 5 min, 变性94℃ 1 min, 退火53℃ 1 min, 延伸温度72℃ 1 min, 共进行35个循环.每个循环中荧光收集在83℃下进行, 以防止由于引物二聚体的存在所引起的误差.溶解曲线程序为65~98℃, 每0.2℃读数, 其间停留6 s. qPCR测定控制软件ABI7500 sequence detection system.
1.3.3 土壤理化性质土壤全氮:采用H2SO4-H2O2消煮-蒸馏滴定测定, 所用仪器为上海纤化仪器公司生产的KDN-02C定氮仪; 土壤全磷:采用H2SO4-H2O2消煮-钒钼黄比色法测定; 土壤全钾:采用H2SO4-H2O2消煮-火焰光度计法测定(FP640型仪器); 土壤碱解氮:碱解扩散法测定; 土壤速效钾:NH4Ac浸提-火焰光度计法测定; 土壤有效磷:0.5 mol·L-1 NaHCO3浸提-钼蓝比色法测定; 土壤有机质:重铬酸钾容量法测定.土壤铵态氮和硝态氮采用2 mol·L-1 KCl提取, 取滤液分别用靛酚蓝比色法和紫外分光光度计法测定.具体测定详细步骤参见文献[20, 21].
1.4 数据处理采用气相色谱(Agilent 7890A气相色谱仪)测定气体N2O浓度. N2O检测器为ECD(电子俘获检测器), 检测温度300℃, 柱温50℃, 气体排放速率由4个气样浓度值经线性回归分析得出具体工作条件详见文献[22].温室气体排放通量(f)的计算公式[2]为:

|
式中, f表示N2O排放通量[μg·(m2·h)-1]; ρ表示标准状态下N2O-N的密度; V为采样箱体积(m3); A为采样底座内土壤表面积(m2); Δc/Δt表示气体的排放速率, 即通过每个小区每次采集的4个样品的气体浓度与时间进行一次线性回归, 回归方程的系数就是Δc/Δt; T为采样箱内温度(℃).
OTU代表着克隆文库中不同基因类型, 可计算各项多样性指数[23, 24].
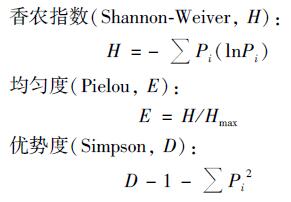
|
式中, Pi为同一种OTU的克隆数占总克隆数的比例, S为不同OTU种类数, Hmax=lnS.
采用SPSS 20.0、Origin 8.5和Excel 2010软件进行数据处理、绘图制表.不同处理间差异性比较采用LSD最小显著差数法和非参数Kruskal-Wallis检验(P﹤0.05).利用Mothur软件对测序的序列进行OTU分析, 然后利用HemI 1.0软件对功能基因进行热度分析.利用CANOCO软件对土壤环境参数和多样性指数相关性进行冗余分析(redundancy analysis, RDA).
2 结果与分析 2.1 秸秆与化肥减量配施对N2O排放的影响在整个试验观测期内, 各处理的土壤N2O排放量存在明显的季节变化规律, 表现为冬季较低, 夏季较高.如图 1所示, 秸秆与化肥配施较对照CK均提高了土壤N2O排放量, 各处理的土壤N2O排放峰值分别出现在2017年的5月12日和6月23日.其中100FS处理的N2O排放量最高, 为57.59~6238.02 μg·(m2·h)-1, 其次为70FS处理, 而F处理的N2O排放量较低, 可见秸秆与化肥配施处理较常规施肥处理提高了土壤N2O排放量.通过计算试验期内的N2O累积排放通量, 秸秆与化肥配施处理的N2O累积排放通量明显高于CK和F处理(P < 0.05), 其中100FS处理高达60.76 kg·hm-2, 其次为70FS处理.

|
图 1 不同处理下N2O排放量和累积排放量 Fig. 1 Emission fluxes and cumulative emission of N2O under different treatments |
如图 2所示, F处理的土壤铵态氮含量最高, 为82.19 mg·kg-1, 其次为100FS(55.72 mg·kg-1), 分别较CK明显增加了79.17 mg·kg-1和52.71 mg·kg-1(P < 0.05).秸秆与化肥减量配施处理较F和100FS处理显著降低土壤铵态氮含量, 但处理间差异不显著.对土壤硝态氮而言, F和100FS处理含量最高, 分别为13.11 mg·kg-1和12.32 mg·kg-1, 而60FS处理的硝态氮含量最低, 为5.00 mg·kg-1.

|
图 2 不同处理下铵态氮和硝态氮含量 Fig. 2 Soil ammonium-and nitrate-nitrogen contents under different treatments |
从表 3可以看出, 施肥处理的土壤总氮(TN)含量较CK提高了0.1~0.2 g·kg-1, 但处理间不存在显著差异. 100FS处理能显著提高土壤碱解氮(AN)含量, 较F处理提高了15.2 mg·kg-1, 而秸秆与化肥减量配施处理较F处理有所下降, 其中70FS和60FS处理显著降低了31.3 mg·kg-1和39.7 mg·kg-1.对土壤可溶性有机氮(DON)而言, 各处理的含量表现为50FS>100FS>F>60FS>70FS>CK处理.与F处理相比, 秸秆与化肥配施处理能增加土壤有机质(SOM)含量, 其中化肥减量配施处理显著提高了4.2~6.6 g·kg-1, 同时F处理的SOM较CK下降了3.8 g·kg-1.秸秆与化肥减量配施处理的pH值较F和100FS处理提高了0.7~0.9个单位, 但处理间差异不显著.从表 3还可以看出, 施肥可以显著提高蔬菜的产量, 秸秆与化肥减量处理的产量较F处理略有下降, 但是差异不显著.
|
|
表 3 不同处理下土壤理化性质的变化1) Table 3 Changes of the physico-chemical soil parameters under different treatments |
2.4 秸秆与化肥减量配施对土壤氨氧化微生物amoA基因丰度及多样性的影响 2.4.1 土壤氨氧化微生物amoA基因丰度
通过荧光定量PCR技术获得土壤AOA和AOB amoA基因拷贝数(图 3).供试土壤中的AOB的基因拷贝数远高于AOA, 可见本研究的土壤AOB占有一定优势.对AOA而言, F处理的土壤中AOA基因拷贝数高达50.9×103 copies·g-1, 明显高于其他处理, 而100FS处理的AOA基因拷贝数明显低于其他处理(P < 0.05), 秸秆与化肥减量配施处理的AOA基因拷贝数不存在显著差异.各处理对AOB基因拷贝数的影响不同于AOA, 如图 3所示, 60FS处理的AOB基因拷贝数最大(1.03×106 copies·g-1), 明显高于其他处理, 其次为50FS和70FS(P < 0.05), 而F处理的AOB基因拷贝数最低(1.36×105 copies·g-1), 但与CK和100FS处理间不存在显著差异.

|
图 3 不同处理下土壤AOA和AOB amoA基因拷贝数 Fig. 3 AOA amoA and AOB amoA gene copies in the soil under different treatments |
不同处理对土壤AOA和AOB amoA基因多样性的影响存在差异.从表 4可以看出, 70FS处理的土壤AOA的OTUs、香农指数(H)、均匀度(E)和优势度(D)均最高, 其次为CK和F处理, 而100FS处理的多样性各指标较低.对土壤AOB而言, 60FS处理的OTUs、H、E和D指数均最高, 而100FS处理的H和D指数最低, 70FS处理的E指数最低.
|
|
表 4 不同处理下土壤AOA和AOB amoA基因多样性指标 Table 4 Biodiversity indices of AOA amoA and AOB amoA in soil under different treatments |
2.4.3 土壤氨氧化微生物热度分析
通过Heatmap分析发现(图 4), 50FS处理的AOA群落结构不同于其他处理, 此外100FS和60FS处理、F和70FS处理的AOA群落结构较为类似; 其中100FS处理的OTU1菌群占有优势地位, F处理OTU3菌群占优势.对AOB而言, CK的群落结构明显不同于其他处理, 50FS和60FS处理的AOB群落结构较为相似, 70FS处理主要以OTU5和OTU1菌群占优势.

|
图 4 不同处理下土壤氨氧化微生物(AOA和AOB)热图 Fig. 4 Heatmap analysis of ammonia-oxidizing microorganism communities (AOA and AOB) in soil under different treatments |
基于微生物群落OTU的组成, 进行土壤氨氧化微生物AOA、AOB与环境因子冗余分析(RDA).如图 5所示, 第一轴和第二轴分别解释了42.9%和27.0%AOA群落结构变化, 40.7%和21.4%的AOB群落结构变化.土壤铵态氮、有机质和有效磷含量对AOA群落结构存在显著影响(P < 0.05, 蒙特卡罗算法); 土壤可溶性有机氮、总氮、速效钾以及有效磷含量与AOB群落结构间具有显著的相关关系(P < 0.05, 蒙特卡罗算法).

|
图 5 不同处理下土壤氨氧化微生物(AOA和AOB)与环境因子变化的RDA排序 Fig. 5 RDA ordination analysis of ammonia-oxidizing microorganism communities (AOA and AOB) in soil under different treatments |
土壤N2O是硝化和反硝化作用的产物, 其中硝化作用是旱地土壤N2O排放的主要途径[7].反应底物(铵态氮)浓度和微生物活性是最直接和核心的控制因素, 控制硝化作用的微生物(AOA和AOB)成为控制N2O排放的重要因素[25].秸秆作为有机物料, 施入土壤后会改变土壤养分组成、温度、水分等, 从而影响硝化作用相关的土壤微生物活性[2].但是, 关于秸秆还田对土壤N2O排放的研究结果仍存在争议, 且不同来源的秸秆C/N比不同对土壤氮素固持转化影响各异.有研究表明, 小麦秸秆的施用在不影响作物产量的条件下, 可以降低土壤N2O排放[26];而Shan等[27]的研究表明, 由于秸秆还田可以为硝化和反硝化微生物提供良好的养分环境, 从而引起水稻秸秆还田处理的N2O排放显著增加了42%. Liu等[28]的研究也表明, 在夏玉米季中进行小麦秸秆还田可以增加58%N2O排放.本研究中, 与对照(CK)和常规施肥(F)相比, 玉米秸秆与化肥配施均不同程度地增加了紫色土N2O排放量和累积排放量, 其中秸秆与全量化肥配施(100FS)处理的土壤N2O排放量和累积排放量最大, 且随着化肥施用量的减少, 土壤N2O排放量呈递减趋势.这是因为玉米秸秆施入土壤后, 一方面可以为微生物提供硝化作用的反应底物参与N2O产生; 另一方面玉米秸秆还田后, 会增加土壤碳素含量(表 3), 为微生物参与下的N2O排放的硝化和反硝化过程提供充足的营养元素[29, 30], 因此与常规施肥处理相比, 秸秆与化肥配施处理提高了土壤N2O排放量.但是玉米秸秆还田会增加土壤C/N比, 从而促进微生物对氮素的固持, 减少N2O产生所需要的氮基质, 最终抑制土壤N2O排放[31], 所以相对于秸秆与全量化肥配施处理而言, 玉米秸秆与化肥减量配施处理的施氮水平低, 在不显著影响作物产量的情况下(表 3), 秸秆与化肥减量配施处理在一定程度上可以降低土壤N2O排放[2].
土壤氨氧化古菌(AOA)和氨氧化细菌(AOB)进行的氨氧化反应是硝化作用的第一步, 控制着硝化作用的速度[9], 同时也是影响N2O排放的重要因素.有研究表明, 施肥(有机肥和无机肥)会增加土壤NH3的含量, 促进了AOB的生长, 但对AOA的影响较小, 从而使得AOA和AOB对施肥表现出不同的响应模式[32].在本研究中, 土壤AOB amoA基因拷贝数普遍比AOA amoA基因拷贝数高1~2个数量级, 可见本试验环境更有利于AOB生长.其中, 常规施肥处理中AOA amoA基因拷贝数高于其他处理, 而AOB amoA基因拷贝数最低. Di等[33]的研究表明AOA和AOB生长的土壤含氮量不同, AOA倾向于营养贫瘠的低N环境, 而AOB对高N环境较为适应; 本试验中常规施肥处理的铵态氮和硝态氮含量高于其他处理, 但高N的环境不利于AOA发挥作用[34].此外Nicol等[35]通过分析AOA和AOB种群延pH 4.5~7.5演替的规律, 发现AOA amoA基因拷贝数与pH呈负相关关系, AOB则呈正相关关系; 曹彦强等[36]的研究表明, 酸性紫色土不利于AOB在氨氧化过程中发挥作用.因此, 在本试验常规施肥处理的酸性紫色土中(pH=4.7), 即使高N的环境也不利于AOB在硝化过程中发挥作用, 这可能是常规施肥处理的AOB amoA基因拷贝数低于其他处理的重要原因.虽然酸性环境中起主导作用的氨氧化微生物是AOA[36], 但高N的环境不利于AOA发挥作用, 因此即使F处理AOA amoA基因拷贝数较高, 但也有可能导致常规施肥处理因硝化作用产生的N2O排放量低于秸秆与化肥配施处理.对于秸秆与全量化肥配施处理, 土壤AOA多样性指数和amoA基因拷贝数均较低.这可能是因为100FS处理的AOA优势菌群OTU1占比最大, 占优势的OTU1菌群大量生长繁殖, 会抑制其他非优势菌群的生长, 从而出现了AOA多样性指数下降和均匀度减少的现象[37, 38], 同时降低了AOA amoA基因拷贝数. 100FS处理的土壤酸性环境(pH=4.7)同样也导致了AOB amoA基因拷贝数明显低于秸秆与化肥减量配施处理(pH 5.4~5.6), 但由于秸秆的施用, 会增加微生物的营养元素, 因此100FS处理的AOB amoA基因拷贝数略高于F处理, 但未达到显著水平, 可见土壤pH对本试验土壤AOB amoA基因拷贝数的影响起到关键性作用.已有研究表明, amoA基因拷贝数并不与氨氧化微生物的活性有直接的联系, 因此其无法真实反映氨氧化微生物的硝化活动[39, 40].虽然100FS处理的AOA和AOB amoA基因拷贝数均显著低于秸秆与化肥减量配施处理, 但N2O排放量显著高于其他处理.一方面是因为在秸秆等量的还田下, 秸秆与化肥减量配施处理的C/N比较高, 促进微生物对氮素的固持, 但在有限氮供应的条件下, 较高的C/N比并不利于微生物对秸秆的降解释放氮素, 因此秸秆与化肥配施处理减少了N2O排放的氮基质(图 2), 从而在一定程度上抑制了N2O排放; 另一方面, AOA可以通过将铵态氮转化为硝态氮, 为反硝化微生物提供代谢底物来间接影响土壤N2O排放, 同时通过Heatmap分析发现, 100FS处理的AOA的优势菌群OTU1占比最高, 该优势菌群可能是硝化作用的主要驱动者, 间接促进了硝化作用中N2O的产生.常规施肥处理的AOA优势菌群OTU1低于秸秆与化肥配施处理, 这也进一步说明了在AOA amoA基因拷贝数高的情况下, F处理N2O排放较少的原因, 这表明氨氧化微生物中的优势菌群才是硝化作用的主要驱动者.
秸秆还田会影响土壤硝化作用相关微生物的种类、数量和活性[41].本研究中AOA的菌群OTU1在6个处理中均存在, 说明此类菌群适应环境范围较广, 生态位相对较宽, 对不同环境因子的耐受性强, 其中100FS处理的OTU1最多, 从而抑制其他AOA菌群的生长, 最终使得100FS处理的多样性最低, 分布最不均匀, 但在秸秆还田的基础上, 减施30%~40%的化肥, 可以有效提高AOA菌群的多样性, 这是因为AOA在低N的环境中也能生长[16], 但是过低的氮量也不利于AOA菌群的生长.这也进一步说明了, 虽然秸秆与化肥减量提高了AOA菌群的多样性, 但是优势菌群OTU1较少, 因此在AOA起氨氧化作用的酸性紫色土中, 相对于秸秆与全量化肥配施处理, 化肥减量处理降低了N2O排放.对AOB群落结构而言, CK处理的AOB群落结构明显不同于其他处理(图 4), 可见无机肥和秸秆的施用会明显改变AOB群落结构.本试验中, 并没有AOB菌群OTU在6个处理中均存在, 说明AOB相较于AOA的生态位较小, 对不同环境因子的耐受性较弱.在秸秆还田的基础上减少30%~40%化肥施用, 也可以提高AOB菌群的多样性, 这可能与该处理下的紫色土有机质含量较高有关, 因为土壤有机质含量对保持土壤微生物多样性具有重要作用[17, 42], 有机质含量越高, 土壤颗粒中微生物群落结构越复杂, 多样性越高[43].通过冗余分析发现, 土壤有机质、铵态氮和有效磷显著影响AOA群落结构, 而显著影响AOB群落结构的因子较多, 这也进一步说明了AOA相较于AOB对不同环境因子的耐受性强.由于本试验紫色土偏酸性, 促使磷生成难溶性物质, 不利于微生物利用[44], 同时加上该研究区属亚热带季风气候, 多雨的环境易造成土壤中盐基离子(如K+)被淋洗, 因此磷和钾元素成为抑制微生物群落结构发展的原因, 方宇等[37]的研究也表明AOA对土壤速效养分有很强的依赖性.
4 结论秸秆与化肥配施较常规处理降低了土壤铵态氮和硝态氮含量, 提高了土壤有机质含量、N2O排放量和累积排放量.秸秆与全量化肥配施处理的蔬菜产量较大, N2O排放量最大, AOA和AOB amoA基因丰度和多样性较低, 但土壤AOA优势菌群才是硝化作用的主要驱动者, 直接和间接地影响N2O排放.土壤AOB amoA基因拷贝数远高于AOA, 但其对不同环境因子的耐受性和生态位低于AOA.在秸秆还田的基础上, 化肥减量30%~40%对蔬菜产量没有显著影响, 且有效提高了紫色土AOA和AOB amoA基因多样性, 在一定程度上减缓了土壤N2O排放.
| [1] |
陈晨, 许欣, 毕智超, 等. 生物炭和有机肥对菜地土壤N2O排放及硝化、反硝化微生物功能基因丰度的影响[J]. 环境科学学报, 2017, 37(5): 1912-1920. Chen C, Xu X, Bi Z C, et al. Effects of biochar and organic manure on N2O emissions and the functional gene abundance of nitrification and denitrification microbes under intensive vegetable production[J]. Acta Scientiae Circumstantiae, 2017, 37(5): 1912-1920. |
| [2] |
黄容, 高明, 黎嘉成, 等. 秸秆与化肥减量配施对菜地土壤温室气体排放的影响[J]. 环境科学, 2018, 39(10): 4694-4704. Huang R, Gao M, Li J C, et al. Effect of straw residues in combination with reducing fertilization rate on greenhouse gas emission in vegetable field[J]. Environmental Science, 2018, 39(10): 4694-4704. |
| [3] | Jetten M S M. The microbial nitrogen cycle[J]. Environmental Microbiology, 2008, 10(11): 2903-2909. DOI:10.1111/emi.2008.10.issue-11 |
| [4] | Wang J Y, Xiong Z Q, Yan X Y. Fertilizer-induced emission factors and background emissions of N2O from vegetable fields in China[J]. Atmospheric Environment, 2011, 45(38): 6923-6929. DOI:10.1016/j.atmosenv.2011.09.045 |
| [5] | Qu Z, Wang J G, Almøy T, et al. Excessive use of nitrogen in Chinese agriculture results in high N2O/(N2O+N2) product ratio of denitrification, primarily due to acidification of the soils[J]. Global Change Biology, 2014, 20(5): 1685-1698. DOI:10.1111/gcb.2014.20.issue-5 |
| [6] | Frame C H, Casciotti K L. Biogeochemical controls and isotopic signatures of nitrous oxide production by a marine ammonia-oxidizing bacterium[J]. Biogeosciences, 2010, 7(9): 2695-2709. DOI:10.5194/bg-7-2695-2010 |
| [7] |
毛新伟, 程敏, 徐秋芳, 等. 硝化抑制剂对毛竹林土壤N2O排放和氨氧化微生物的影响[J]. 土壤学报, 2016, 53(6): 1528-1540. Mao X W, Cheng M, Xu Q F, et al. Effects of nitrification inhibitors on soil N2O emission and community structure and abundance of ammonia oxidation microorganism in soil under extensively managed Phyllostachys edulis stands[J]. Acta Pedologica Sinica, 2016, 53(6): 1528-1540. |
| [8] | Philippot L, Andert J, Jones C M, et al. Importance of denitrifiers lacking the genes encoding the nitrous oxide reductase for N2O emissions from soil[J]. Global Change Biology, 2011, 17(3): 1497-1504. |
| [9] |
周志成, 罗葵, 唐前君, 等. 不同施肥方式对红壤蔬菜田氨氧化细菌和氨氧化古菌群落的影响[J]. 中国蔬菜, 2015(7): 33-39. Zhou Z C, Luo K, Tang Q J, et al. Effect of different fertilization on ammonia-oxidizing bacteria and ammonia-oxidizing archaea in red soil vegetable field[J]. China Vegetables, 2015(7): 33-39. DOI:10.3969/j.issn.1000-6346.2015.07.008 |
| [10] | Zhong W H, Bian B Y, Gao N, et al. Nitrogen fertilization induced changes in ammonia oxidation are attributable mostly to bacteria rather than archaea in greenhouse-based high N input vegetable soil[J]. Soil Biology and Biochemistry, 2016, 93: 150-159. DOI:10.1016/j.soilbio.2015.11.003 |
| [11] | Maeda K, Spor A, Edel-Hermann V, et al. N2O production, a widespread trait in fungi[J]. Scientific Reports, 2015, 5: 9697. DOI:10.1038/srep09697 |
| [12] | Attard E, Recous S, Chabbi A, et al. Soil environmental conditions rather than denitrifier abundance and diversity drive potential denitrification after changes in land uses[J]. Global Change Biology, 2011, 17(5): 1975-1989. DOI:10.1111/j.1365-2486.2010.02340.x |
| [13] | Huang R, Lan M L, Liu J, et al. Soil aggregate and organic carbon distribution at dry land soil and paddy soil:the role of different straws returning[J]. Environmental Science and Pollution Research, 2017, 24(36): 27942-27952. DOI:10.1007/s11356-017-0372-9 |
| [14] |
王沛譞, 徐焱, 宋亚娜. 转基因水稻秸秆还田对土壤硝化反硝化微生物群落的影响[J]. 中国生态农业学报, 2018, 26(1): 8-15. Wang P X, Xu Y, Song Y N. Effect of transgenic rice straw return to soil on nitrification and denitrification microbial community[J]. Chinese Journal of Eco-Agriculture, 2018, 26(1): 8-15. |
| [15] |
陈春兰, 陈哲, 朱亦君, 等. 水稻土细菌硝化作用基因(amoA和hao)多样性组成与长期稻草还田的关系研究[J]. 环境科学, 2010, 31(6): 1624-1632. Chen C L, Chen Z, Zhu Y J, et al. Effects of rice straw on the diversity of nitrifying genes (amoA and hao) in paddy soil[J]. Environmental Science, 2010, 31(6): 1624-1632. |
| [16] | Chen X, Zhang L M, Shen J P, et al. Abundance and community structure of ammonia-oxidizing archaea and bacteria in an acid paddy soil[J]. Biology and Fertility of Soils, 2011, 47(3): 323-331. DOI:10.1007/s00374-011-0542-8 |
| [17] |
萨如拉, 高聚林, 于晓芳, 等. 玉米秸秆深翻还田土壤氨氧化细菌amoA基因多样性分析[J]. 玉米科学, 2015, 23(3): 107-111. Sa L L, Gao J L, Yu X F, et al. Analysis of the diversity of ammonia oxidizing bacteria with amoA gene in deep returning maize straw soils[J]. Journal of Maize Sciences, 2015, 23(3): 107-111. |
| [18] |
赵伟烨, 王智慧, 曹彦强, 等. 石灰性紫色土硝化作用及硝化微生物对不同氮源的响应[J]. 土壤学报, 2018, 55(2): 479-489. Zhao W Y, Wang Z H, Cao Y Q, et al. Response of nitrification and nitrifiers to different nitrogen sources in an alkaline purple soil[J]. Acta Pedologica Sinica, 2018, 55(2): 479-489. |
| [19] | 张福锁, 陈新平, 陈青. 中国主要作物施肥指南[M]. 北京: 中国农业大学出版社, 2009. |
| [20] | 鲍士旦. 土壤农业化学分析方法[M]. (第三版). 北京: 中国农业出版社, 2000. |
| [21] | 杨剑虹, 王成林, 代亨林. 土壤农化分析与环境监测[M]. 北京: 中国大地出版社, 2008. |
| [22] |
郭艳亮, 王丹丹, 郑纪勇, 等. 生物炭添加对半干旱地区土壤温室气体排放的影响[J]. 环境科学, 2015, 36(9): 3393-3400. Guo Y L, Wang D D, Zheng J Y, et al. Effect of biochar on soil greenhouse gas emissions in semi-arid region[J]. Environmental Science, 2015, 36(9): 3393-3400. |
| [23] | Kvålseth T O. Note on biological diversity, evenness, and homogeneity measures[J]. Oikos, 1991, 62(1): 123-127. DOI:10.2307/3545460 |
| [24] | Magurran A E. Ecological diversity and its measurement[M]. Cambridge: Princeton University Press, 1988: 7-47. |
| [25] | Cui P Y, Fan F L, Yin C, et al. Urea-and nitrapyrin-affected N2O emission is coupled mainly with ammonia oxidizing bacteria growth in microcosms of three typical Chinese arable soils[J]. Soil Biology and Biochemistry, 2013, 66: 214-221. DOI:10.1016/j.soilbio.2013.08.001 |
| [26] | Htun Y M, Tong Y A, Gao P C, et al. Coupled effects of straw and nitrogen management on N2O and CH4 emissions of rainfed agriculture in Northwest China[J]. Atmospheric Environment, 2017, 157: 156-166. DOI:10.1016/j.atmosenv.2017.03.014 |
| [27] | Shan J, Yan X Y. Effects of crop residue returning on nitrous oxide emissions in agricultural soils[J]. Atmospheric Environment, 2013, 71: 170-175. DOI:10.1016/j.atmosenv.2013.02.009 |
| [28] |
刘琳, 吴彦, 何奕忻, 等. 季节性雪被对高山生态系统土壤氮转化的影响[J]. 应用生态学报, 2011, 22(8): 2193-2200. Liu L, Wu Y, He Y X, et al. Effects of seasonal snow cover on soil nitrogen transformation in alpine ecosystem:a review[J]. Chinese Journal of Applied Ecology, 2011, 22(8): 2193-2200. |
| [29] | Zhu T B, Zhang J B, Yang W Y, et al. Effects of organic material amendment and water content on NO, N2O, and N2 emissions in a nitrate-rich vegetable soil[J]. Biology and Fertility of Soils, 2013, 49(2): 153-163. DOI:10.1007/s00374-012-0711-4 |
| [30] | Yang H S, Xu M M, Koide R T, et al. Effects of ditch-buried straw return on water percolation, nitrogen leaching and crop yields in a rice-wheat rotation system[J]. Journal of the Science of Food and Agriculture, 2016, 96(4): 1141-1149. DOI:10.1002/jsfa.2016.96.issue-4 |
| [31] |
宋贺, 王成雨, 陈清, 等. 长期秸秆还田对设施菜田土壤反硝化特征和N2O排放的影响[J]. 中国农业气象, 2014, 35(6): 628-634. Song H, Wang C Y, Chen Q, et al. Effects of long-term amendment of residue on denitrification characteristics and N2O emissions in greenhouse soil[J]. Chinese Journal of Agrometeorology, 2014, 35(6): 628-634. DOI:10.3969/j.issn.1000-6362.2014.06.004 |
| [32] | Prosser J I, Nicol G W. Archaeal and bacterial ammonia-oxidisers in soil:the quest for niche specialisation and differentiation[J]. Trends in Microbiology, 2012, 20(11): 523-531. DOI:10.1016/j.tim.2012.08.001 |
| [33] | Di H J, Cameron K C, Shen J P, et al. Ammonia-oxidizing bacteria and archaea grow under contrasting soil nitrogen conditions[J]. FEMS Microbiology Ecology, 2010, 72(3): 386-394. |
| [34] | Jia Z J, Conrad R. Bacteria rather than Archaea dominate microbial ammonia oxidation in an agricultural soil[J]. Environmental Microbiology, 2009, 11(7): 1658-1671. DOI:10.1111/emi.2009.11.issue-7 |
| [35] | Nicol G W, Schleper C. Ammonia-oxidising crenarchaeota:important players in the nitrogen cycle?[J]. Trends in Microbiology, 2006, 14(5): 207-212. DOI:10.1016/j.tim.2006.03.004 |
| [36] |
曹彦强, 闫小娟, 罗红燕, 等. 不同酸碱性紫色土的硝化活性及微生物群落组成[J]. 土壤学报, 2018, 55(1): 194-202. Cao Y Q, Yan X J, Luo H Y, et al. Nitrification activity and microbial community structure in purple soils with different pH[J]. Acta Pedologica Sinica, 2018, 55(1): 194-202. |
| [37] |
方宇, 景晓明, 王飞, 等. 长期施肥对氨氧化古菌丰度及群落结构的影响[J]. 植物营养与肥料学报, 2015, 21(6): 1607-1614. Fang Y, Jing X M, Wang F, et al. Effect of long-term fertilization on abundance and community structure of ammonia-oxidizing archaea in paddy soil[J]. Journal of Plant Nutrition and Fertilizer, 2015, 21(6): 1607-1614. |
| [38] | Shen J P, Zhang L M, Zhu Y G, et al. Abundance and composition of ammonia-oxidizing bacteria and ammonia-oxidizing archaea communities of an alkaline sandy loam[J]. Environmental Microbiology, 2008, 10(6): 1601-1611. DOI:10.1111/emi.2008.10.issue-6 |
| [39] | Leininger S, Urich T, Schloter M. Archaea predominate among ammonia-oxidizing prokaryotes in soils[J]. Nature, 2006, 442(7104): 806-809. DOI:10.1038/nature04983 |
| [40] | Hatzenpichler R. Diversity, physiology, and niche differentiation of ammonia-oxidizing archaea[J]. Applied and Environmental Microbiology, 2012, 78(21): 7501-7510. DOI:10.1128/AEM.01960-12 |
| [41] |
白雪, 夏宗伟, 郭彦玲, 等. 硝化抑制剂对不同旱地农田土壤N2O排放的影响[J]. 生态学杂志, 2012, 31(9): 2319-2329. Bai X, Xia Z W, Guo Y L, et al. Effects of nitrification inhibitors on N2O emission from different upland agricultural soils[J]. Chinese Journal of Ecology, 2012, 31(9): 2319-2329. |
| [42] | Yang Y H, Yao J, Hu S, et al. Effects of agricultural chemicals on DNA sequence diversity of soil microbial community:a study with RAPD marker[J]. Microbial Ecology, 2000, 39(1): 72-79. |
| [43] | Sessitsch A, Weilharter A, Gerzabek M H. Microbial population structures in soil particle size fractions of a long-term fertilizer field experiment[J]. Applied and Environmental Microbiology, 2001, 67(9): 4215-4224. DOI:10.1128/AEM.67.9.4215-4224.2001 |
| [44] |
胡红青, 贺纪正, 李学垣, 等. 有机酸对酸性土壤吸附磷的影响[J]. 华中农业大学学报, 1997, 16(1): 37-42. Hu H Q, He J Z, Li X Y, et al. Effect of organic acids on phosphate adsorption by acid soil[J]. Journal of Huazhong Agricultural University, 1997, 16(1): 37-42. |