 2019, Vol. 40
2019, Vol. 40



制药、造纸、采矿等工业所排放的富硫酸盐废水对会对环境和人体健康造成极大危害.传统的物理化学处理方法如化学沉淀、离子交换和液膜分离等技术成本较高[1].由于富硫酸盐废水中一般缺乏有机物, 因而, 基于自养型硫酸盐还原菌(sulfate-reducing bacteria, SRB)的生物修复技术已引起人们的广泛关注[2~5].
微生物电解系统(microbial electrolysis system, MES)是近年来发展的一种生物电化学系统, 在废水处理以及能源回收等领域具有极大的潜能[6]. MES反应装置均由阴、阳极室组成.在阳极室, 产电微生物以有机物作为电子供体释放电子至阳极, 电子经外电路传递至(非)生物阴极.通过构建生物阴极型MES, 电化学活性菌可从阴极间接或直接获取电子实现多种污染物的还原去除, 如SO42-、NO3-、对氯硝基苯、三氯乙烯等[7~10].由于阴极的受体物质普遍具有较低的还原电位, 且阴极电极过电位以及欧姆损失的存在, 在MES的阴极和阳极之间通常需要施加低值外电压来促使反应的进行.目前, 基于双室MES生物阴极还原硫酸盐的研究探究了自养硫酸盐还原[3, 4]、重金属锌与硫酸盐的协同去除[7], 以及单质硫的回收[5]等, 但多采用0.8 V左右的外加电压.关于外加电压对MES的影响, 据报道, 外加电压的升高(0.8~1.4 V)可以促进MES阳极有机物氧化过程中电子的释放[11, 12]; Coma等[11]研究异养型生物阴极硫酸盐还原发现, 硫酸盐还原速率和能耗均随着电压的升高(0.7~1.4 V)而升高; 亦有研究表明提高外加电压(0~2.0 V)或电场(0~3.5 mA)可以促进生物膜胞外聚合物的分泌, 但高于0.8 V或1.5 mA时则会抑制阴极微生物的生长代谢[13, 14].可见, 外加电压的大小可显著影响MES的运行性能, 然而, 阴极生物膜作为去除污染物的关键因子, 其对外电压改变的响应尚属未知.
本研究通过构建双室MES, 探究外电压为0.4、0.5、0.6、0.7和0.8 V情形下自养生物阴极还原硫酸盐的性能, 同时, 探讨不同外加电压下阴极生物膜的胞外聚合物及群落响应, 这对生物阴极型MES在低能耗条件下实现硫酸盐废水的自养还原及机制研究有着重要的意义.
1 材料与方法 1.1 自养SRB混合菌群的富集珠江底泥(23°06′N, 113°17′E)作为接种源, 位于广东省广州市滨江东路中山大学北门.配置0.72 g·L-1硫酸盐基质(约500 mg·L-1 SO42-), 使用1 mol·L-1 H3PO4调节pH至6.0用于驯化自养型SRB[15].将珠江底泥与硫酸盐基质接种至100 mL厌氧瓶中, 并在顶空通入2 min混合气[H2-CO2 (80:20)]用作SRB生长代谢所需的碳源和氢源, 同时驱赶瓶中残留的氧气[3].将厌氧瓶置于恒温振荡器(30℃, 160 r·min-1), 4 d一个周期, 进行长期培养富集.定时监测厌氧瓶中SO42-的浓度, 待基质中SO42-的去除率趋于稳定时, 即可作为MES阴极室的接种液.
1.2 双室MES的构建双室MES反应器均由有机玻璃构件通过螺丝组装而成, 分为阴、阳极两个室, 中间由阳离子交换膜(CEM, Ultrex CMI-7000, 美国)隔开.碳纤维编制的碳刷作为阳极电极, 使用前需在马弗炉中450℃条件下进行30 min预热处理, 以去除碳刷表面的氧化物[16].石墨碳板作为阴极电极, 使用前需进行打磨、切割成长、宽、高各为30、20、2 mm, 用酸碱浸泡24 h后, 冲洗, 浸泡于去离子水中[17].阴、阳两极使用环氧树脂固定, 钛丝连接进行电流传输, 在外部串联一个10 Ω电阻用于采集电流信息, 插入电极后每个反应室的有效容积为30 mL, 具体装置如图 1所示.

|
图 1 MES生物阴极还原硫酸盐原理示意 Fig. 1 Schematic diagram of the sulfate-reducing biocathode MES |
大坦沙污水处理厂位于广东省广州市荔湾区(23°07′N, 113°12′E), 将其二沉池出水和1 g·L-1乙酸钠基质(COD浓度约为780 mg·L-1)按50%的体积比接种至MES阳极室, 用于富集阳极产电微生物[3].将厌氧瓶中预先富集的SRB菌液与硫酸盐基质按50%的体积比接种至阴极室, 以48 h为一个周期放置于生化培养箱(LRH-150B, 中国)30℃条件下培养. MES反应器先在恒定电压0.8 V(IT6720, 美国)条件下驯化10 d, 阴极室通入1 min混合气[H2-CO2(80:20)]用于加速阴极生物膜的形成.随后, 撤去混合气并将外电压降低, 在0.4、0.5、0.6、0.7和0.8 V电压下进行批次培养. MES反应器运行稳定时, 选择数据和趋势较平行的3个批次进行平均值和方差分析.使用多通路电压数据采集器(Integra 2700, 美国)采集反应器定值电阻两端实时电压数据(每隔15 min自动采集).饱和甘汞电极(0.242 V vs. SHE, 雷磁232型, 中国)被插入阳极室与阴极室用于观测不同外电压下电势的变化.
1.4 检测与计算分析阴极室进出水SO42-与S2-分别使用硫酸钡比浊法与亚甲蓝分光光度法进行监测; 阳极室进出水COD变化使用重铬酸钾分光光度法进行监测.收集阴极室的气体, 使用气相色谱仪(岛津GC-2014, 日本)分析H2的含量.根据电压数据采集器采集的MES两端电压值U计算电流I=U/(R), 将电流对运行时间(t, s)积分按公式(1)计算得到MES体系的周期电荷量Q(C), 并根据公式(2)计算得到电流密度曲线IV(A·m-3).
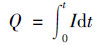
|
(1) |
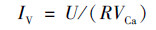
|
(2) |
式中, R为定值电阻(Ω), VCa为MES阴极室容积(m3)
根据出水SO42-的浓度获得MES生物阴极还原硫酸盐的量(mol), 按公式(3)计算得出该体系用于还原SO42-的电子回收率CE(%).

|
(3) |
式中, 8为将硫酸根还原为硫化物所需的电子数目, Δn为引进去除的硫酸根物质的量(mol), F为法拉第常数(96 485 C·mol-1), Q为周期电荷量(C).
阴极聚多糖与蛋白量分别使用苯酚-硫酸分光光度法和考马斯亮蓝法进行测定, 用无菌刀片切割5 mm×5 mm的阴极碳板, 测试前将样品置于3 mL 0.2 mol·L-1 NaOH剧烈振荡促使附着在阴极上的生物膜分散至溶液中, 然后用3 mL去离子水冲洗, 将两者混合所得的样品用于后续聚多糖及蛋白量的分析测定[18, 19].
采用SPPSS 19.0软件进行方差齐同质性检验与单因素方差分析(one-way ANOVA), 通过LSD和Dunnet T3检验对不同外电压下MES自养型生物阴极还原硫酸盐性能及阴极生物膜聚多糖和蛋白量的数据进行多重比较.首先进行方差齐同质性检验, 查看方差齐性检验结果, 选择LSD(方差齐性检验方法, P < 0.05表示差异显著)检验不同外电压对阳极COD去除率、硫酸盐去除率、电子回收率、蛋白量和生物量影响的显著性; 选择Dunnet T3(方差不齐检验方法, P < 0.05表示差异显著)检验不同外电压对周期电荷量、硫酸盐还原速率、SO42--S2-转化效率和聚多糖影响的显著性.
1.5 16S rRNA基因高通量测序经45 d运行后, 将MES反应器拆开, 使用无菌刀片将阴极生物膜刮落, 进行送样测序用于分析阴极微生物群落的变化.上机测序前, 根据E.Z.N.A.® soil试剂盒(Omega Bio-tek, Norcross, GA, U.S.)操作手册进行样品总DNA抽提, DNA浓度和纯度利用NanoDrop2000进行检测, 利用1%琼脂糖凝胶电泳检测DNA提取质量; 使用338F (ACTCCTACGGGAGGCAGCAG)和806R (GGACTA CHVGGGTWTCTAAT)引物对V3-V4可变区进行扩增, 随后, 扩增产物在Illumina MiSeq测序平台(Majorbio, 上海, 中国)上建库进行上机测序.高质量的序列在97%的相似性下被归类为一个运算分类单元(OTU), 每个OTU里丰度最高的序列被选取作为代表序列, 然后将这些代表序列使用RDP classifier(http://rdp.cme.msu.edu/classifier/classifier.jsp)按70%置信度分类到门和属水平, 用于分析阴极微生物群落的变化, 同时使用Chao1和ACE指标评估物种的丰富度, 以及Shannon和Coverage指标评估物种的多样性和样品有效序列的覆盖度[20].高通量测序获得的原始序列数被保存在NCBI SRA, 登录号为SRP149289.
2 结果与讨论 2.1 不同外电压下生物阴极型MES的产电性能分析如图 2(a)所示, 更换基质后, 0.4~0.8 V电压下MES均在更换基质的6 h以内获得最高电流密度.随着外加电压的升高, 电流密度也随之升高, 最大值分别可达(6.1±0.3)、(12.8±0.7)、(19.4±0.5)、(35.4±1.9)和(48.0±2.3)A·m-3.以48 h为一个运行周期, 0.8 V电压下MES的周期电荷量显著高于0.4 V(P < 0.05), 分别为152.9 C±7.1 C和19.1 C±0.2 C.最大电流密度、周期电荷量与外电压之间分别呈正相关, 其相关系数分别为0.92和0.99[图 2(b)], 此结果与文献[12]报道一致.为进一步探讨外加电压对生物阳极和阴极的影响, 分别监测了两电极电势随时间的变化.阳极电势表现出周期平台期, 随着外电压的升高, 平台期电势分别为-0.24、-0.24、-0.19、-0.16和-0.12 V(vs. SHE)[图 2(c)].阳极液COD的监测结果表明, 阳极微生物COD去除率随着外加电压的增加而显著增加(P < 0.05), 在0.4、0.7和0.8 V电压下, COD平均去除率分别为17.2%、34.1%和44.5%, 其消耗量分别为130.7、259.0和337.8 mg·L-1, 这意味着0.8 V电压下阳极微生物具有更高的代谢活性(表 1). Logan等[21]发现阳极电势的升高有利于产电微生物氧化有机物释放电子至电极将化学能转化为电能, 从而导致更高的电流输出.阴极电势在外电压为0.4~0.6 V的范围内变化较平稳, 随着外电压升高, 电势值分别为-0.63、-0.66和-0.74 V; 0.7 V和0.8 V电压下, 阴极电势峰值出现在-0.84 V左右[图 2(d)].阴极电势的降低有利于阴极微生物获取能量进行生长代谢, 但当阴极电势过低时, 微生物所获得的有效能量却会减少[22, 23]. 0.8 V和0.4 V电压下阳极和阴极电势差值分别为0.12 V和0.20 V, 表明外电压的施加值对阴极电势影响更大.据报道, 在生物电化学系统中阴极过电位明显高于阳极过电位[24].电活性微生物可有效降低电极的过电位, 在生物阴极型MES中, 阳极表面产电微生物的生物量远大于阴极表面, 如Xiang等[12]对产乙酸MES的研究表明阳极生物量高出阴极生物量约8倍.因此, 阳极电势比阴极电势更稳定, 具体的微生物作用机制还需要进一步的研究.

|
(a)电流密度曲线;(b)最大电流密度、周期电荷量与外加电压值之间的相关性分析;(c)阳极电势;(d)阴极电势 图 2 不同外加电压下MES的电化学特性 Fig. 2 Electrochemical properties of the MES under different applied voltage |
MES阴极液中SO42-与S2-浓度随时间的变化如图 3所示.各外加电压下, SO42-浓度均随着MES的运行逐渐降低, 伴随着S2-浓度的逐渐升高, 而在非生物阴极条件下阴极室并无硫酸盐去除与硫化物产生[4, 15], 表明生物还原是SO42-去除的主要途径.经计算, 0.4~0.8 V电压范围内, SO42-的平均还原速率分别为7.9、31.5、48.8、78.9和68.0 g·(m3·d)-1(表 1).经单因素方差分析中Dunnet T3检验多重比较发现, 在0.05的显著性水平下, 随着外电压的升高(0.4~0.7 V), 硫酸盐还原速率显著升高, 而当外电压进一步升高时, 硫酸盐还原速率并无显著变化, 且0.7 V电压下的硫酸盐还原速率略高于0.8 V.与此同时, 出水中S2-浓度分别增加到(4.1±0.4)、(13.2±0.7)、(17.9±0.9)、(31.9±2.2)和(25.1±1.4)mg·L-1.这些结果表明, 外电压为0.7 V时MES可以获得最大的硫酸盐还原去除, 该还原速率与Hu等[15]报道的0.8 V电压下的硫酸盐还原速率相似[68.7 g·(m3·d)-1].根据SO42-的还原量计算得到, 0.4、0.5、0.6、0.7和0. 8 V电压下的电子回收率分别为27.6%±3.2%、34.2%±1.1%、35.7%±1.5%、41.8%±2.6%和31.9%±3.5%.经单因素方差分析中LSD检验多重比较发现, 0.5 V和0.6 V电压下的电子回收率显著高于0.4 V, 与0.8 V电压下的电子回收率并无显著差异, 且0.7 V电压下的电子回收率显著高于0.8 V(P < 0.05), 表明0.5~0.7 V电压范围内的阴极电子能更有效地被SRB获取用于还原硫酸盐.收集阴极室的气体进行分析, 仅在0.8 V电压下监测到少量H2, 这与Luo等[3]在不同阴极电势条件下MES阴极室观测到的H2变化趋势一致, 由此推测0.8 V电压下H2的快速产生和逸出使得阴极电子未能被SRB高效利用, 从而造成相对低的硫酸盐还原率和电子回收率[10].根据化学方程式(4)计算SO42--S2-转换效率(%), 即阴极室出水实际含有的S2-与理论上产生的S2-之比, 获得MES阴极的SO42--S2-转化率分别为78.6%(0.4 V)、62.8%(0.5 V)、55.2%(0.6 V)、60.6%(0.7 V)和55.6%(0.8 V), 各电压下SO42--S2-转化率并无显著差异(P < 0.05).从化学方程式(5)可以看出, 可能是因为阴极还原产物在溶液pH中性条件下会有部分以H2S和HS-形式存.
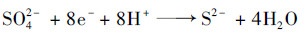
|
(4) |
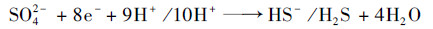
|
(5) |

|
图 3 不同外加电压下MES阴极室SO42-与S2-浓度变化 Fig. 3 Concentration of SO42- and S2- in the cathode chamber changing with time under different applied voltage |
|
|
表 1 生物阴极型MES在不同电压下的硫酸盐还原性能比较1) Table 1 Comparison of the performance and energy recovery efficiency of the SRB-biocathode MES under different applied voltages |
2.3 MES阴极生物量在不同外加电压下的变化
为了更好地解释生物阴极型MES在不同外电压下的性能, 进一步测定了阴极生物膜的胞外聚多糖和蛋白量并对该数据进行单因素方差分析(表 2).阴极生物膜的生物量在电压的升高下呈显著增加的趋势(P < 0.05), 0.8 V和0.7 V电压下为2 748.6 μg·cm-2和2 325.9 μg·cm-2, 相比于0.4 V(1 611.5 μg·cm-2)提高了70%和44%.阴极生物膜的聚多糖从0.4 V的605.0 μg·cm-2提高至0.7 V的928.6 μg·cm-2和0.8 V的1 185.1 μg·cm-2, 相对应地, 蛋白量从1 006.5 μg·cm-2提高至1 397.3 μg·cm-2和1 563.5 μg·cm-2.方差分析表明阴极生物膜的聚多糖在不同外电压间并无显著差异, 0.7 V和0.8 V电压下的蛋白量显著高于0.4 V(P < 0.05), 阴极生物膜的蛋白量受外电压的影响较大.关于生物膜胞外聚合物的研究表明, 聚多糖在细胞固定以及形成生物膜过程中起到重要作用, 而蛋白量可用于评估生物膜的活性[14, 19].根据0.4 V和0.8 V的比较可见, 外电压的升高有利于高活性生物膜的快速形成. Gong等[25]发现外电压的升高有利于阴极与阳极间的电子传输从而进一步地促进阳极产电生物膜的生长. Sun等[26]通过提高MES外加电压获得较高的电流输出从而探究阳极电活性生物膜对电流的响应, 发现输出电流的升高有利于高活性生物膜在阳极表面的生长. 0.8 V下生物膜的聚多糖(附着)和蛋白量(活性)比0.7 V均有所提高, 这与0.8 V下获得最高的输出电流密度相对应; 然而其硫酸盐去除性能却低于0.7 V, 可能是因为0.8 V电压下更利于析氢反应, 一方面H+离子与SO42-竞争电子, 另一方面H2被非硫酸盐还原菌用于自身增殖.
|
|
表 2 不同外加电压下阴极生物膜多糖和蛋白量 Table 2 Polysaccharose and protein contents of the biocathode under different applied voltages |
2.4 MES阴极生物膜群落在不同外加电压下的响应
采用16S rRNA基因高通量测序技术对阴极微生物群落进行多样性分析, 结果如表 3所示.不同外电压下样品的有效序列覆盖度均为99.9%, 表明测序结果的可靠性[14].将获得的有效序列按97%相似水平进行聚类, 所得的OTUs分别为132(0.4 V)、132(0.5 V)、155(0.6 V)、137(0.7 V)和135(0.8 V). Chao1指数与ACE指数代表着物种的丰富度, Shannon指数反映了物种的多样性, 而在0.8 V电压下阴极生物膜群落物种丰富度、多样性与0.4 V、0.5 V电压下无明显差异, 且略低于0.6 V和0.7 V的情形.结果表明, 0.4~0.8 V电压范围内阴极微生物群落多样性并未随着电压的升高而发生明显的变化.
|
|
表 3 总体测序结果与α多样性指数 Table 3 Overall pyrosequencing information and α diversity indices for the bacterial samples |
不同外电压下MES阴极微生物群落门和属水平的分布如图 4所示. Proteobacteria在MES阴极生物膜群落结构的门水平分布中占主导, 其在0.4、0.5、0.6、0.7和0.8 V电压下的相对丰度分别为95.6%、95.7%、76.5%、67.4%和78.8%[图 4(a1)]. Proteobacteria包含许多已知的电化学活性菌和与硫代谢相关的菌, 因此在硫酸盐还原型生物阴极占据主导[27].生物膜中丰度较高的还包括Firmicutes和Bacteroidetes, 这与已有研究中关于生物阴极微生物群落的分布情况一致[3, 20, 28]. Firmicutes与Bacteroidetes的相对丰度随着电压的上升逐渐升高, 在0.7 V电压下获得最大值分别为5.1%和15.3%, 在0.8 V电压下降低至3.5%和11.1%. Firmicutes包含的一些革兰氏阳性菌, 能够利用电子介体进行胞外电子转移[29]; Bacteroidetes在阳极生物膜的胞外电子传递过程中拥有独特的优势[30].除此之外, 随外电压升高呈现先上升后下降趋势的菌群还有Actinobacteria、Synergistetes和Spirochaetae[图 4(a2)], 表明随着电压的升高选择性的富集了一些菌群, 而这些菌群可能在种间电子传递的过程中起着作用.

|
(a1)和(a2)门水平分布; (b)为属水平分布 图 4 阴极微生物群落相对丰度 Fig. 4 Relative abundance of the cathodic bacterial community |
Desulfovibrio在阴极微生物群落属水平分布中占主导, 其在0.4、0.5、0.6、0.7和0.8 V电压下的相对丰度分别为60.3%、86.5%、61.0%、64.1%和70.3%[图 4(b)].这与Teng等[7]报道的结果相似, 其研究中硫酸盐还原生物阴极表面Desulfovibrio的相对丰度为66%. Desulfovibrio是一类典型的电化学活性SRB, 在许多硫酸盐还原型生物电化学系统中均检测到该菌的存在.在本研究中, Desulfovibrio的相对丰度并未随着外加电压的升高发生明显的波动, 表明Desulfovibrio在利用电极呼吸代谢方面具有独特的优势且能够应对电势波动的冲击.已有研究证实SRB能直接从电极获取电子还原硫酸盐, 无需电子介体或H2的存在[4].生物电化学系统中, 阴极电化学活性菌主要通过细胞膜表面的氧化还原组分如细胞色素c从电极直接获取电子[31]. Lentimicrobium在0.6~0.8 V电压范围内占据8.5%左右的相对丰度, 其可利用糖类物质在厌氧环境下发酵产氢[32], 而阴极微生物细胞凋亡的过程中可能存在一些糖类物质, 为其提供能源的同时被转化为H2. Halothiobacillus在0.4~0.8 V电压范围内占据2~7%左右的相对丰度, 该菌是一类硫氧化菌, 在微氧环境下能够以硫化物作为电子供体进行化能自养, 该类菌的存在可能会使还原产物负二价硫氧化为单质硫或是硫酸盐[33, 34], 这为废水中硫的回收提供了可能性.此外, 0.4 V电压下选择性地富集了g_norank_f_Hydrogenophilaceae与g_norank_f_Hyphomicrobiaceae, 其相对丰度分别为7%和12%, 据文献报道, 这两种菌可利用阴极作为电子供体进行自养生长[35], 具体的代谢途径仍需进一步研究. 0.7 V电压下还发现了Acetobacterium, 已有文献报道Acetobacterium可利用阴极作为唯一电子供体将CO2转化为乙酸[12], Acetobacterium和Desulfovibrio在生物电化学系统中存在共代谢关系[36], Acetobacterium有可能通过电合成乙酸的方式促进生物阴极的电子利用和还原硫酸盐的性能.
由于Desulfovibrio在MES生物阴极占绝对主导地位, 其极可能在还原硫酸盐的过程中起着主要作用, 因此, 对Desulfovibrio的种水平结构进行了进一步分析(图 5).从图 5中可以看出, 尽管外电压引起阴极电势的较大变化, 但生物阴极表面始终以Desulfovibrio magneticus RS-1和s_unclassified_g_Desulfovibrio为主导. 0.4、0.5、0.6、0.7和0.8 V电压下Desulfovibrio magneticus RS-1的相对丰度分别为20.4%、58.5%、43.7%、40.2%和63.2%; s_unclassified_g_Desulfovibrio的相对丰度分别为77.7%、40.6%、51.8%、50.9%和34.1%, 二者的总丰度均大于90%.据报道, Desulfovibrio magneticus RS-1是一种趋磁细菌, 可利用可溶性Fe2+在生物体内形成磁性矿物, 在磁场的作用下进行富集[37].近期, Zhou等[38]通过对生物电化学系统施加电磁场发现外加的电磁场能够延长放电周期, 明显提高电流输出, 且有利于电化学活性菌如Geobacter的富集. Desulfovibrio magneticus RS-1的富集与电磁场之间有可能存在某种关系, 后续可对电磁场作用下Desulfovibrio magneticus RS-1的响应机制进行深入研究. s_unclassified_g_Desulfovibrio的相对丰度随电压升高呈现降低趋势, 在外加电压为0.4 V下的相对丰度高达77.7%, 因而推测其具有直接利用阴极作为电子供体的能力, 但仍有待进一步研究. Desulfovibrio vulgaris的相对丰度在0.7 V电压下为5%, 而其余电压下都低于0.2%. Liamleam等[2]提出Desulfovibrio vulgaris能够以H2作为电子供体还原硫酸盐, 是一种典型的氢营养型SRB.此外, Desulfovibrio gigas在0.8 V电压下具有一定的丰度为1.2%, 该菌具有镍铁氢化酶能够催化H2的产生[39].结果表明, 0.7~0.8 V外加电压下, 由于H2析出速度的增加, 氢营养型SRB开始占据主导, 从而影响MES生物阴极还原硫酸盐的性能.

|
图 5 不同外加电压下脱硫弧菌属在种水平下的分布 Fig. 5 Relative abundance of Desulfovibrio at the species level under different applied voltages |
(1) 外加电压为0.4~0.8 V时, MES的输出电流、周期电荷量、COD去除量与外加电压(0.4~0.8 V)呈正相关关系; 硫酸盐还原量随着电压的升高先升高后降低, 在0.7 V时获得最大硫酸盐还原速率[78.9 g·(m3·d)-1]和最高S2-出水浓度(31.9 mg·L-1±2.2 mg·L-1); MES的电子回收率均在30%左右, 在0.7 V时获得最高值为41.8%, ,推测产氢可能是电子损失的一个途径.
(2) 阴极表面生物量在外电压的升高下呈显著增加的趋势, 0.8 V电压下相比0.4 V提高了70%;阴极微生物群落结构分析发现, Proteobacteria在门水平分布中占主导, Desulfovibrio在属水平分布中占主导; Desulfovibrio的相对丰度并未随着外加电压的升高发生明显的波动, 表明Desulfovibrio在利用电极呼吸代谢方面具有独特的优势且能够应对电势波动的冲击.
| [1] |
相凤欣, 任立人, 吴丹, 等. 生物处理含硫酸盐废水生成单质硫的研究进展[J]. 环境工程, 2012, 30(S2): 151-155. Xiang F X, Ren L R, Wu D, et al. Advances in bio-treatment of sulfate-laden wastewater and conversion to element sulfur[J]. Environmental Engineering, 2012, 30(S2): 151-155. |
| [2] | Liamleam W, Annachhatre A P. Electron donors for biological sulfate reduction[J]. Biotechnology Advances, 2007, 25(5): 452-463. DOI:10.1016/j.biotechadv.2007.05.002 |
| [3] | Luo H P, Fu S Y, Liu G L, et al. Autotrophic biocathode for high efficient sulfate reduction in microbial electrolysis cells[J]. Bioresource Technology, 2014, 167: 462-468. DOI:10.1016/j.biortech.2014.06.058 |
| [4] | Su W T, Zhang L X, Tao Y, et al. Sulfate reduction with electrons directly derived from electrodes in bioelectrochemical systems[J]. Electrochemistry Communications, 2012, 22: 37-40. DOI:10.1016/j.elecom.2012.04.030 |
| [5] | Blázquez E, Gabriel D, Baeza J A, et al. Treatment of high-strength sulfate wastewater using an autotrophic biocathode in view of elemental sulfur recovery[J]. Water Research, 2016, 105: 395-405. DOI:10.1016/j.watres.2016.09.014 |
| [6] |
符诗雨, 刘广立, 骆海萍, 等. 微生物电解系统生物阴极的硫酸盐还原特性研究[J]. 环境科学, 2014, 35(2): 626-632. Fu S Y, Liu G L, Luo H P, et al. Characterization of biocatalysed sulfate reduction in a cathode of microbial electrolysis system[J]. Environmental Science, 2014, 35(2): 626-632. |
| [7] | Teng W K, Liu G L, Luo H P, et al. Simultaneous sulfate and zinc removal from acid wastewater using an acidophilic and autotrophic biocathode[J]. Journal of Hazardous materials, 2016, 304: 159-165. DOI:10.1016/j.jhazmat.2015.10.050 |
| [8] | Zhan G Q, Zhang L X, Li D P, et al. Autotrophic nitrogen removal from ammonium at low applied voltage in a single-compartment microbial electrolysis cell[J]. Bioresource Technology, 2012, 116: 271-277. DOI:10.1016/j.biortech.2012.02.131 |
| [9] |
周亚, 杨春. 生物电化学系统对4-氯硝基苯的降解[J]. 化工进展, 2018, 37(1): 375-380. Zhou Y, Yang C. Degradation of 4-chloronitrobenzene by bioelectrochemical system[J]. Chemical Industry and Engineering Progress, 2018, 37(1): 375-380. |
| [10] | Aulenta F, Reale P, Catervi A, et al. Kinetics of trichloroethene dechlorination and methane formation by a mixed anaerobic culture in a bio-electrochemical system[J]. Electrochimica Acta, 2008, 53(16): 5300-5305. DOI:10.1016/j.electacta.2008.02.084 |
| [11] | Coma M, Puig S, Pous N, et al. Biocatalysed sulphate removal in a BES cathode[J]. Bioresource Technology, 2013, 130: 218-223. DOI:10.1016/j.biortech.2012.12.050 |
| [12] | Xiang Y B, Liu G L, Zhang R D, et al. High-efficient acetate production from carbon dioxide using a bioanode microbial electrosynthesis system with bipolar membrane[J]. Bioresource Technology, 2017, 233: 227-235. DOI:10.1016/j.biortech.2017.02.104 |
| [13] | Ding A Q, Yang Y, Sun G D, et al. Impact of applied voltage on methane generation and microbial activities in an anaerobic microbial electrolysis cell (MEC)[J]. Chemical Engineering Journal, 2016, 283: 260-265. DOI:10.1016/j.cej.2015.07.054 |
| [14] | Wang K, Sheng Y X, Cao H B, et al. A novel microbial electrolysis cell (MEC) reactor for biological sulfate-rich wastewater treatment using intermittent supply of electric field[J]. Biochemical Engineering Journal, 2017, 125: 10-17. DOI:10.1016/j.bej.2017.05.009 |
| [15] | Hu J P, Zeng C P, Liu G L, et al. Magnetite nanoparticles accelerate the autotrophic sulfate reduction in biocathode microbial electrolysis cells[J]. Biochemical Engineering Journal, 2018, 133: 96-105. DOI:10.1016/j.bej.2018.01.036 |
| [16] | Hou Y P, Luo H P, Liu G L, et al. Improved Hydrogen Production in the Microbial Electrolysis Cell by Inhibiting Methanogenesis Using Ultraviolet Irradiation[J]. Environmental Science & Technology, 2014, 48(17): 10482-10488. |
| [17] | LaBelle E V, Marshall C W, Gilbert J A, et al. Influence of acidic pH on hydrogen and acetate production by an electrosynthetic microbiome[J]. PLoS One, 2014, 9(10): e109935. DOI:10.1371/journal.pone.0109935 |
| [18] | DuBois M, Gilles K A, Hamilton J K, et al. Colorimetric method for determination of sugars and related substances[J]. Analytical Chemistry, 1956, 28(3): 350-356. DOI:10.1021/ac60111a017 |
| [19] | Zhang X Y, Cheng S A, Wang X, et al. Separator characteristics for increasing performance of microbial fuel cells[J]. Environmental Science & Technology, 2009, 43(21): 8456-8461. |
| [20] | Luo H P, Teng W K, Liu G L, et al. Sulfate reduction and microbial community of autotrophic biocathode in response to acidity[J]. Process Biochemistry, 2017, 54: 120-127. DOI:10.1016/j.procbio.2016.12.025 |
| [21] | Logan B E, Hamelers B, Rozendal R, et al. Microbial fuel cells:methodology and technology[J]. Environmental Science & Technology, 2006, 40(17): 5181-5192. |
| [22] | Finkelstein D A, Tender L M, Zeikus J G. Effect of electrode potential on electrode-reducing microbiota[J]. Environmental Science & Technology, 2006, 40(22): 6990-6995. |
| [23] | Liang P, Fan M Z, Cao X X, et al. Evaluation of applied cathode potential to enhance biocathode in microbial fuel cells[J]. Journal of Chemical Technology and Biotechnology, 2009, 84(5): 794-799. DOI:10.1002/jctb.v84:5 |
| [24] | Rozendal R A, Hamelers H V M, Molenkamp R J, et al. Performance of single chamber biocatalyzed electrolysis with different types of ion exchange membranes[J]. Water Research, 2007, 41(9): 1984-1994. DOI:10.1016/j.watres.2007.01.019 |
| [25] | Gong Y M, Ebrahim A, Feist A M, et al. Sulfide-driven microbial electrosynthesis[J]. Environmental Science & Technology, 2013, 47(1): 568-573. |
| [26] | Sun D, Cheng S A, Zhang F, et al. Current density reversibly alters metabolic spatial structure of exoelectrogenic anode biofilms[J]. Journal of Power Sources, 2017, 356: 566-571. DOI:10.1016/j.jpowsour.2016.11.115 |
| [27] | Bond D R, Holmes D E, Tender L M, et al. Electrode-reducing microorganisms that harvest energy from marine sediments[J]. Science, 2002, 295(5554): 483-485. DOI:10.1126/science.1066771 |
| [28] | Wrighton K C, Virdis B, Clauwaert P, et al. Bacterial community structure corresponds to performance during cathodic nitrate reduction[J]. ISME Journal, 2010, 4(11): 1443-1455. DOI:10.1038/ismej.2010.66 |
| [29] | Xing D F, Cheng S A, Logan B E, et al. Isolation of the exoelectrogenic denitrifying bacterium Comamonas denitrificans based on dilution to extinction[J]. Applied Microbiology and Biotechnology, 2010, 85(5): 1575-1587. DOI:10.1007/s00253-009-2240-0 |
| [30] | Ishii S, Suzuki S, Norden-Krichmar T M, et al. Microbial population and functional dynamics associated with surface potential and carbon metabolism[J]. ISME Journal, 2014, 8(5): 963-978. DOI:10.1038/ismej.2013.217 |
| [31] | Rosenbaum M, Aulenta F, Villano M, et al. Cathodes as electron donors for microbial metabolism:Which extracellular electron transfer mechanisms are involved?[J]. Bioresource Technology, 2011, 102(1): 324-333. DOI:10.1016/j.biortech.2010.07.008 |
| [32] | Sun L W, Toyonaga M, Ohashi A, et al. Lentimicrobium saccharophilum gen. nov., sp. nov., a strictly anaerobic bacterium representing a new family in the phylum Bacteroidetes, and proposal of Lentimicrobiaceae fam. nov.[J]. International Journal of Systematic and Evolutionary Microbiology, 2016, 66(7): 2635-2642. DOI:10.1099/ijsem.0.001103 |
| [33] | LaBarge N, Yilmazel Y D, Hong P Y, et al. Effect of pre-acclimation of granular activated carbon on microbial electrolysis cell startup and performance[J]. Bioelectrochemistry, 2017, 113: 20-25. DOI:10.1016/j.bioelechem.2016.08.003 |
| [34] | Sorokin D Y, Tourova T P, Muyzer G, et al. Thiohalospira halophila gen., sp. nov. and Thiohalospira alkaliphila sp. nov., novel obligately chemolithoautotrophic, halophilic, sulfur-oxidizing gammaproteobacteria from hypersaline habitats[J]. International Journal of Systematic and Evolutionary Microbiology, 2008, 58(7): 1685-1692. DOI:10.1099/ijs.0.65654-0 |
| [35] | Zhang G D, Feng S S, Jiao Y, et al. Cathodic reducing bacteria of dual-chambered microbial fuel cell[J]. International Journal of Hydrogen Energy, 2017, 42(45): 27607-27617. DOI:10.1016/j.ijhydene.2017.06.095 |
| [36] | Xiang Y B, Liu G L, Zhang R D, et al. Acetate production and electron utilization facilitated by sulfate-reducing bacteria in a microbial electrosynthesis system[J]. Bioresource Technology, 2017, 241: 821-829. DOI:10.1016/j.biortech.2017.06.017 |
| [37] | Byrne M E, Ball D A, Guerquin-Kern J L, et al. Desulfovibrio magneticus RS-1 contains an iron-and phosphorus-rich organelle distinct from its bullet-shaped magnetosomes[J]. Proceedings of the National Academy of Sciences of the United States of America, 2010, 107(27): 12263-12268. DOI:10.1073/pnas.1001290107 |
| [38] | Zhou H H, Liu B F, Wang Q S, et al. Pulse electromagnetic fields enhance extracellular electron transfer in magnetic bioelectrochemical systems[J]. Biotechnology for Biofuels, 2017, 10: 238. DOI:10.1186/s13068-017-0929-3 |
| [39] | Volbeda A, Charon M H, Piras C, et al. Crystal structure of the nickel-iron hydrogenase from Desulfovibrio gigas[J]. Nature, 1995, 373(6515): 580-587. DOI:10.1038/373580a0 |