 2018, Vol. 39
2018, Vol. 39

2. 陕西省循环农业工程技术研究中心, 杨凌 712100
 , DAI Yin-yue1,2 , ZHANG Wei1,2 , FU Shu-yue1,2 , YANG Gai-he1,2
, DAI Yin-yue1,2 , ZHANG Wei1,2 , FU Shu-yue1,2 , YANG Gai-he1,2 , HAN Xin-hui1,2 , CHEN Zheng-xing1,2 , GAO De-xin1,2 , FENG Yong-zhong1,2
, HAN Xin-hui1,2 , CHEN Zheng-xing1,2 , GAO De-xin1,2 , FENG Yong-zhong1,2
2. Shaanxi Engineering Research Center of Circular Agriculture, Yangling 712100, China
退耕还林还草工程实施以来, 黄土丘陵地区植被覆盖显著增加[1, 2], 土壤侵蚀得到有效控制[3]、土壤质量明显改善[4], 生态效益逐渐提高[5].植被恢复过程实质上是植物与土壤相互反馈的协同作用过程[6].其中, 土壤为植物生长提供必要的水分和养分, 而植物在生长过程中以凋落物和根系等形式实现土壤养分循环和质量改良[7].因此, 明确植被恢复过程中植物群落特征与土壤养分变化的关系, 对揭示黄土丘陵区植被恢复过程及土壤性质变化趋势具有重要的科学意义.
植物作为生态系统的生产者, 在长期的生长过程中逐渐形成了独特的形态、生理和物候特征.研究表明, 不同科植物的叶片养分含量存在差异, 如豆科植物中N含量较多, 禾本科植物的C含量较高, 菊科植物叶片中P和K含量大于禾本科和豆科[8].在植物以凋落物的形式为土壤提供养分输入的过程中[9], 不同物种进行养分归还速度和数量不同[10], 如阔叶树的养分归还比例大于针叶树[11], 次生林的养分归还量大于原生林[12], 从而导致土壤养分状况存在差异.此外, 不同造林物种还可以通过影响土壤微生物群落组成及多样性[13]、土壤酶活性[14]和植物的根系分泌物[15], 导致土壤养分含量的差异.综上, 不同植物种群通过其自身独特的生物学特性影响土壤养分的来源及代谢过程, 从而导致不同的植物-土壤反馈机制[16].然而, 植被恢复过程中物种组成变化引起的土壤理化性质及生物学活性变化过程极为复杂, 相关研究仍不是很明确, 如研究发现植被恢复中, 植物的多样性、生物量[17]以及植物的群落组成[18, 19]对土壤造成影响, 但是恢复过程中不同功能组植物(植物科属组成)对于土壤特性的影响机理亟待深入研究.
撂荒演替是一种改善土壤条件和恢复退化环境的有效途径, 已成为生态学研究的热门话题之一[20].有研究表明, 撂荒演替过程中物种组成变化表现为由一年生草本群落逐渐向多年生草本和小灌木群丛转变[21], 菊科植物逐渐取代禾本科植物成为群落优势物种, 其中豆科植物伴生[22], 物种多样性也随演替年限的增加而增加[23].撂荒恢复过程中群落结构组成及多样性的变化会促进土壤理化性质的改善以及微生物丰富度和酶活性的提升.有学者将土壤理化性质的变化与植物群落进行了相关性分析, 多集中于数学关系的研究层面, 有研究表明土壤养分和物种多样性呈显著的正相关性[18], 土壤有机质和全氮与物种多样性[19]及地上生物量[17]呈显著正相关, 但缺乏土壤养分对物种组成变化响应过程的深入分析, 尤其是对不同功能组植物是如何影响土壤养分及生物学特征的研究.因此, 本文选择黄土丘陵区耕地撂荒后不同恢复年限的草地序列, 通过对植被群落与多样性特征、土壤养分含量与酶活性变化特征及相关性关系分析, 以期为深入认知撂荒地恢复过程中植物-土壤的互馈与协同恢复机制提供科学依据.
1 材料与方法 1.1 研究区域概况研究区位于陕西省延安市吴起县杨青川流域(36°33′33″~37°24′27″N, 107°38′57″~108°32′49″E), 海拔1 233~1 809 m, 土壤为黄绵土, 地形为黄土高原粱峁丘陵沟壑区, 属暖温带半干旱气候, 年平均降水量为478.3 mm, 雨季在7~9月, 年均温度7.8℃, 年日照时间约为2 400 h, 年最高气温在7~8月(37℃), 年最低气温在1~2月(-20℃).草本植被多以禾本科(Poaceae)、豆科(Leguminosae)和菊科(Compositae)为主, 优势种主要为猪毛蒿(Artemisia scoparia)、铁杆蒿(Artemisia sacrorum)、茭蒿(Artemisia giraldii)、达乌里胡枝子(Lespedeza dahurica)及白羊草(Bothriochloa flaccidum)等.
1.2 样地选择与植被调查通过查阅资料及实地调查, 2015年7~8月于流域内选择撂荒10、15、20、30和45 a的草地群落作为研究对象, 详细信息见表 1.每个草地中设置3个20 m×20 m的大样方, 3个样方之间的距离超过了大多数土壤变量的空间依赖性(<13.5 m)[13], 在每个大样方中按“S”形随机地选取10个1 m×1 m的小样方, 使用样方法调查记录样方内所有出现的草本植物的植物名, 使用标尺测量样方内每株植物的高度, 同种植物的盖度及密度[18].并使用GPS进行样地经纬度、海拔及坡向等样地信息的采集.
|
|
表 1 采样点基本信息及土壤基本性质1) Table 1 Characteristics of the soil sampling sites and basic soil properties |
1.3 土壤样品采集
在每个20 m×20 m的大样方中按照“S”形选取10个采样点, 除去表面凋落物及植物残体后, 用直径为5 cm的土钻采集表层(0~10 cm)土壤, 将同一样地内10个土样混合均匀后分成两份, 一份过2 mm筛网筛分后4℃冷藏保存, 用于测定土壤酶活性, 另一份室温下风干后用于测定土壤理化性质.
1.4 土壤理化性质及土壤酶的测定采用烘干法测定土壤含水量(soil water content, SWC)[24], 环刀法测量容重(soil bulk density, SBD)[25].土壤有机碳(SOC)使用重铬酸钾容量法测定, 全氮(TN)采用半微量凯氏测定法, 全磷(TP)利用碳酸钠熔融-钼锑抗比色法测定, 速效氮(AN)采用KCl浸提流动注射分析仪测定, 速效磷(AP)采用碳酸氢钠法测定[25].碱性磷酸酶(alkaline phosphatase, ALP)使用磷酸苯二钠比色法测定, 用1 g土壤中2 h后的苯酚量(mg)表示; 过氧化氢酶(catalase, CAT)采用高锰酸钾滴定法测定, 以1 g土壤20 min后消耗0.1 mol·L-1的高锰酸钾量(mL)表示; 脲酶(urease, UE)利用苯酚钠-次氯酸钠比色法测定, 活性以24 h后1 g土壤中的NH4+-N量(mg)表示; 蔗糖酶(saccharase, SC)采用3, 5-二硝基水杨酸比色法测定, 以24 h后1 g土壤中含有的葡萄糖量(mg)表示[26, 27].
1.5 物种多样性计算选取Gleason丰富度指数(G, Gleason index指数)、Simpson多样性指数(D, Simpson index指数)、Shannon-Wiener多样性指数(H, Shannon-Wiener index指数)和Pielou均匀度指数(E, Pielou index指数)这4个指数来进行物种多样性的衡量[28~29], 选取重要值来反映物种在这一群落的优势程度[30].
Gleason丰富度指数(G):

|
Simpson多样性指数(D):
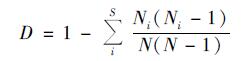
|
Shannon-Wiener多样性指数(H):
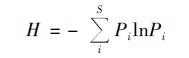
|
Pielou均匀度指数(E):
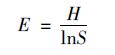
|
式中, S为物种数量, A为样方面积, Ni为i种的重要值, N为样地中全部种的重要值之和; Pi为i种的相对重要值, Pi=Ni/N.

|
使用Excel和SPSS 21.0对数据进行计算及分析, 利用单因素方差分析(one-way DUNCAN)进行差异显著性检验, Pearson相关分析进行植物多样性与土壤养分及酶活性的相关性分析, 利用多项式拟合(Polynomial Fit)对各多样性指数进行线性拟合, 使用Canoco 5对植被及土壤进行PCA、RDA分析, 并用Origin 8.5及Canoco 5绘图.
2 结果与分析 2.1 不同年限草地植物群落物种组成 2.1.1 不同年限草地植物群落物种数量特征不同撂荒年限草地群落中科、属、种的组成如表 2所示.不同撂荒年限草地群落总物种数量表现为:20 a>15 a=30 a>45 a=10 a.总科数、总属数随着撂荒年限的增加均呈先增加后降低的趋势, 且均在撂荒20 a的群落中物种数量最大, 撂荒10 a的群落物种数量最小.菊科、禾本科以及豆科植物在不同撂荒年限草地中均有分布, 且不同撂荒年限草地群落均以菊科、禾本科、豆科为主, 三大科的物种数量随着撂荒年限的增加而增加, 但三大科占本群落的比率呈先降后升的趋势, 占比从大到小的顺序为45 a>30 a>10 a>20 a=15 a.
|
|
表 2 不同撂荒年限草地植被科、属、种组成 Table 2 Number of plant families, genus, and species for different years of abandonment |
2.1.2 不同撂荒年限草地的群落恢复特征
本研究结果显示, 不同撂荒年限草地群落物种组成存在差异, 且PCA分析表明其可分成四类(图 1), 分别是10 a群落、15 a群落、20 a群落、30 a群落和45 a群落.草地撂荒恢复过程中, 共分布15科35属43种植物(表 3).按物种重要值来看, 猪毛蒿、达乌里胡枝子、铁杆蒿、茭蒿、白羊草等菊科、禾本科以及豆科植物在不同撂荒年限草地中均占优势地位, 且部分物种虽在不同撂荒年限草地群落中均有分布, 但它们在群落中的地位及作用不同.撂荒10 a草地优势种为猪毛蒿, 亚优势种为鹅观草(Roegneria kamoji), 伴生种为甘草(Glycyrrhiza uralensis)与草地早熟禾(Poa pratensis)等; 撂荒15 a草地群落优势种仍为猪毛蒿, 亚优势种有达乌里胡枝子和赖草(Leymus secalinus), 伴生种为阿尔泰狗娃花(Heteropappus altaicus)与茭蒿等; 恢复至撂荒20 a时, 铁杆蒿与达乌里胡枝子逐渐成为草地优势种, 假苇拂子茅(Calamagrostis pseudophragmites)成为亚优势种, 大针茅(Stipa grandis)及茭蒿等物种为群落的伴生种; 撂荒30 a时, 茭蒿取代达乌里胡枝子和铁杆蒿成为草地群落优势种, 铁杆蒿变为亚优势种, 达乌里胡枝子与白羊草则成为伴生种; 撂荒45 a时, 白羊草及茭蒿为草地的优势种, 亚优势种为铁杆蒿及达乌里胡枝子.45 a撂荒恢复过程中, 豆科的达乌里胡枝子与菊科的铁杆蒿在5个年限草地中均有分布, 禾本科的赖草及菊科的茭蒿主要分布在撂荒15 a以后的草地中, 唇形科(Labiatae)的活血丹(Glechoma longituba)、香青兰(Dracocephalum moldlavica)及蔷薇科(Rosaceae)的翻白委陵菜(Potentilla discolor)、二裂委陵菜(Potentilla bifurca)在撂荒10~20 a的草地中分布, 而蔷薇科的蛇莓(Duchesnea indica)与堇菜科(Violaceae)的紫花地丁(Viola philippica)主要在撂荒20~45a的草地中分布, 其中还有一些伞形科(Umbelliferae)、旋花科(Convolvulaceae)、藜科(Chenopodiaceae)、败酱科(Valerianaceae)、毛茛科(Ranunculaceae)、茜草科(Rubiaceae)、远志科(Polygalaceae)、牻牛儿苗科(Geraniaceae)及车前科(Plantaginaceae)等物种在撂荒恢复过程中作为偶见种在草地中分布.

|
图 1 草地植被群落物种分布主成分分析 Fig. 1 Principal component analysis of the plant species distribution in the grassland community |
|
|
表 3 不同撂荒年限草地植物群落物种重要值/% Table 3 Importance value of the plant species in plant communities in different years of abandonment/% |
在不同撂荒年限草地群落的优势种不断发生变化, 但达乌里胡枝子、铁杆蒿、茭蒿及赖草等菊科、豆科和禾本科植物的优势度始终较大.随着撂荒年限的增加, 豆科植物重要值呈增加趋势, 10~45 a豆科植物重要值分别为21.66%、23.29%、27.88%、29.82%、33.38%;禾本科植物重要值呈先增加, 后趋于平缓的趋势, 10~45 a禾本科植物重要值分别为18.73%、19.83%、22.05%、27.57%、26.08%;菊科植物重要值呈降低趋势, 10~45 a菊科植物重要值分别为48.80%、44.37%、36.98%、33.07%、26.97%.三大科重要值之和占草地群落植物总重要值85%以上, 为植物群落分布的主要物种.
2.2 不同撂荒年限草地群落的多样性利用抛物线方程对各多样性指数进行拟合可得到二次拟合图像, 发现草地群落的多样性指数随着撂荒年限的增加变化明显.从图 2可以得出, 在撂荒恢复10~45 a, G、H和E随着撂荒年限的增加均呈现出先增后减趋势, 且都在撂荒20 a达到最大值.G最小值在撂荒45 a, H与E的最小值在撂荒10 a.D随着撂荒年限的增加与其他多样性指数的变化趋势不同, 呈先减后增的趋势, 最大值出现在撂荒10 a, 最小值出现在撂荒15 a.

|
图 2 不同撂荒年限草地群落的多样性指数 Fig. 2 Diversity indexes of grassland communities in different years of abandonment |
不同撂荒年限草地土壤养分含量如图 3所示.SOC含量随着撂荒年限的增加呈逐渐增加的趋势, 从撂荒10 a的4.69g·kg-1显著增加到撂荒45 a的11.15 g·kg-1, 相比增加了2.38倍; 土壤AP在土壤中含量的变化趋势与SOC相同, 从撂荒10 a的0.5 mg·kg-1显著增加到撂荒45 a的2.05 mg·kg-1, 相比增加了4.1倍; 土壤TN、TP含量随着撂荒年限的增加呈先增后减的趋势, 且均在撂荒30 a达最大值, 分别为0.73 g·kg-1与0.56 g·kg-1; 土壤AN含量随着撂荒年限的增加呈先减后增的趋势, 在撂荒15 a时达最小值到23.32 mg·kg-1.总的来说, 草地群落土壤养分含量随着撂荒年限的增加呈波动型增加.

|
图 3 不同撂荒年限草地土壤养分含量 Fig. 3 Soil nutrient content in plant communities in different years of abandonment |
不同撂荒年限草地群落土壤养分化学计量比如图 4所示.草地群落土壤中C:N随着撂荒年限的增加呈先减后增的趋势, 最小值出现在撂荒20 a, C:N从撂荒10~30 a增减都不显著, 30~45 a增加显著; 草地群落土壤C:P随着撂荒年限的增加呈增加的趋势, 撂荒45a相比撂荒10a, C:P增加了2.24倍; 草地群落土壤N:P随着撂荒年限的增加呈先增后减的趋势, N:P比值的增减范围在0.3~1.3之间, 最大比值出现在撂荒30 a.

|
图 4 不同撂荒年限草地群落土壤化学计量比 Fig. 4 C, N, and P stoichiometry in plant communities in different years of abandonment |
4种土壤酶随着撂荒年限的增加所表现出的趋势如图 5所示.随着撂荒年限的增加, 土壤中ALP与UE活性均呈现出持续增加的趋势, 其中ALP活性在撂荒45 a时相比撂荒10 a增加了1.66倍, UE活性在撂荒45 a时相比撂荒10 a增加了2.22倍, 且增加显著; 土壤中CAT和SC活性随着撂荒年限的增加呈现先增后减的趋势, 其中CAT活性在撂荒15~20 a增加显著, 在撂荒20 a达到最大值后活性降低但不显著, SC活性在撂荒30 a达最大值后显著降低.总体上看土壤酶活性是随着撂荒年限的增加而升高的.

|
图 5 不同撂荒年限草地群落土壤酶活性 Fig. 5 Soil enzymes in plant communities in different years of abandonment |
对不同撂荒年限草地群落及其多样性与土壤养分和土壤酶的关系采用冗余度(RDA)分析, 结果如图 6.将土壤养分及土壤酶作为响应变量时, 菊科、禾本科及豆科对其有极显著影响(菊科:贡献率=72.8%, F=29.5, P=0.002;豆科:贡献率=69.1%, F=25, P=0.002;禾本科:贡献率=66.0%, F=22, P=0.002), 盖度对其有显著影响(盖度:贡献率=25.4%, F=4.1, P=0.032), 总属数、总种数、总科数及植物多样性对土壤养分及土壤酶没有显著性影响.将SBD及SWC作为响应变量时, 盖度、总科数、总种数、G、H及总属数对其有极显著影响(盖度:贡献率=73.5%, F=35, P=0.002;总科数:贡献率=54.1%, F=15.1, P=0.004;总种数:贡献率=54.0%, F=15, P=0.004;G:贡献率=54.0%, F=15, P=0.004;H:贡献率=45.9%, F=10.9, P=0.01;总属数:贡献率=42.7%, F=9.6, P=0.01), 菊科及禾本科对其有显著性影响(菊科:贡献率=32.3%, F=6.1, P=0.036;禾本科:贡献率=29.9%, F=5.5, P=0.046), 其他草地群落指标对SWC及SBD没有显著性影响.

|
Leg:豆科; Poa:禾本科; Cov:盖度; Com:菊科; D:Simpson 多样性指数; E:Pielou 均匀度指数; G:Gleason 丰富度指数; H:Shannon-Wiener 多样性指数; ALP:碱性磷酸酶; UE:脲酶; SC:蔗糖酶; CAT:过氧化氢酶; SWC:土壤含水量; SBD:土壤容重; 图中Gleason丰富度指数和物种总种数箭头重合 图 6 草地群落组成及多样性与土壤养分和土壤酶关系的RDA分析 Fig. 6 RDA analysis among plant families, genus, species, and diversity indexes and C, N, and P and enzymes of the soil |
以上结果得出草地群落的不同科植物对土壤养分及酶活性影响较大.对禾本科, 菊科及豆科的重要值与土壤养分及土壤酶进行相关性分析.由表 4可看出, 禾本科对SOC、TN、AN、AP、ALP、UE和SC呈极显著正相关(P<0.01), 对TP和CAT呈显著正相关(P<0.05);豆科与SOC、TN、AN、AP、ALP、CAT、UE和SC呈极显著正相关(P<0.01), 豆科对TP没有显著相关性; 与其他科不同, 菊科与SOC、TP、AN、AP、ALP、CAT、UE和SC呈极显著负相关(P<0.01), 菊科对TP无显著相关性.
|
|
表 4 三大科重要值与土壤养分及土壤酶的相关性分析1) Table 4 Correlations between the importance of the three families, soil nutrients, and enzymes |
3 讨论
随着撂荒年限的增加, 撂荒恢复各研究阶段主要群落分别为猪毛蒿(10 a、15 a)达乌里胡枝子+铁杆蒿(20 a)茭蒿+铁杆蒿(30 a)白羊草+茭蒿(45 a), 且恢复过程中优势物种均为豆科, 菊科及禾本科植物.草地群落的科、属、种数量呈先增后减的趋势, 菊科、禾本科和豆科物种数量占群落总种数比例呈先减后增的趋势, 三大科植物占比由撂荒10 a的66.67%上升至撂荒45 a的75%(表 2).这是由三大科植物独特的生物学特性决定的, 它们能适应该地区干旱少雨、土壤贫瘠的生态环境[31].菊科和禾本科植物通过增加根系生物量及根系深度来获取更多的水分[32], 豆科与根瘤菌的共生可使其在相对干旱条件下快速生长[33], 并且与土壤产生协同作用增加土壤养分含量, 为其他物种提供养分资源[34].同时, 植物总生产力的增加提高了凋落物生物量, 凋落物覆盖的增加减少了土壤水分蒸发及地表径流, 土壤含水量升高[35], 从而促进植物的生长, 其他科物种数量增加.在撂荒20~30 a间, 草地群落的生物量增加、总蒸腾量增多, 致使土壤含水量逐渐降低, 进而导致植物群落多样性减少, 而三大科植物相比其他科植物更加耐旱, 在干旱胁迫下, 豆科、菊科、禾本科植物通过增加根冠比[36], 产生抗旱性酶类[37]等来适应干旱环境, 且撂荒30 a后铁杆蒿、猪毛蒿等菊科植物化感作用的增强, 抑制了其他草本植物种子的发芽和幼苗的生长[38], 所以三大科植物占群落总数的比例及优势度逐渐增高.三大科植物优势度的改变也影响了物种的多样性, 本研究中多样性指数G、H和E随着撂荒年限的增加均呈现出先增后减趋势(图 2), 郭曼等[39]在黄土高原的研究也有相同的结果, 但草地的G、H和E指数明显高于林下草本多样性指数[40], 可能是因为有乔木林竞争及郁闭作用.此外, 物种的生物学特性、菊科植物的化感作用、种内种间竞争、SWC和SBD等因素也会影响物种的多样性[41].
生态恢复是植被与土壤协同变化的过程, 地上植物群落的结构组成和生物量变化必然会引起地下土壤理化性质的改变.本研究结果显示, 在植被恢复后10~45 a, 土壤养分及酶活性随着撂荒年限的增加呈增加趋势.因撂荒过程中植被生产力增加使群落生物量和凋落物的覆盖增加, 为土壤提供了更多的养分归还, 同时地表覆盖增加使得土壤含水量增加, 且凋落物会响应水分含量增加而加速分解[42], 进而导致养分含量增加.禾本科与豆科植物含有大量的C与N, 植物自身营养物质归还给土壤后使得SOC及土壤TN的含量增加显著(图 3).菊科种内竞争较为激烈, 数量及重要值在撂荒年限增加过程中有减少的趋势, 并且其自身的碳、氮含量不如豆科及禾本科植物[43], 所以菊科与养分含量呈负相关关系.相对于SOC和土壤TN, 土壤TP含量在撂荒过程中增加缓慢(图 3), 这主要是由于土壤P主要来源于微生物介导的含磷矿化物分解与转化[44].C、N和P的不平衡增加导致C/N、C/P和N/P比值发生变化, 尤其是C/P和N/P显著增加(图 4), 表明P元素逐渐成为植物生长的限制性元素[45].此外, 植物群落通过影响土壤酶活性控制的土壤养分代谢过程来影响土壤养分含量[46].本研究中土壤酶活性在撂荒10~20 a显著增加, 而撂荒20 a后部分土壤酶如SC、CAT活性有降低现象(图 5), 且三大科植物对土壤酶活性有显著影响(表 4).撂荒10~20 a植物快速生长, 外源基质(凋落物和根)输入量增加, 土壤微生物及植物活动分泌的代谢产物(土壤酶)增加[47], 且不同植被下土壤理化性质及酶活性不同, 导致土壤养分状况提高[48, 49].随着植被的恢复, 植物耗水增加, 土壤含水量在20 a后下滑, 导致土壤微生物生物量及活性也逐渐降低[50], 引起部分酶活性降低.本研究结果显示, 禾本科与豆科植物与参与C、N和P代谢酶的活性均呈显著正相关, 而菊科则表现为负相关(表 4).一方面是由于禾本科与豆科植物富含丰富的C和N, 可为酶的合成提供丰富的原料[46]; 另一方面, 禾本科和豆科植物富含蛋白质, 需大量ALP、UE、CAT和SC参与其凋落物蛋白化合物水解、有机磷化合物分解及含碳化合物分解过程[51].菊科植物由于其成丛分布数量较少, 在群落中重要值逐渐降低, 导致其与4种酶活性呈负相关, 这也表明菊科植物对土壤的碳氮磷代谢过程成负反馈效应.此外, 与三大科物种相比, 整体物种多样性与土壤养分和酶活性的关系相对较小, 表明撂荒恢复过程中土壤养分主要受某些特定功能群影响[52, 53], 本研究同时发现植物功能群落相对于物种多样性, 更能反映土壤的养分状况.综上, 在撂荒恢复过程中三大科植物占比及重要值逐渐增高, 其通过自身养分归还为土壤提供营养物质, 同时影响土壤微生物及酶活性, 这影响了代谢及养分分解过程, 从而对土壤性质整体状况产生直接与间接的影响.
4 结论撂荒恢复10~45 a过程中, 各阶段主要群落分别为猪毛蒿达乌里胡枝子+铁杆蒿茭蒿+铁杆蒿白羊草+茭蒿, 草地群落的科、属、种数量呈现先增后减的趋势, 菊科、禾本科和豆科物种数量及重要值占群落总种数比例呈先减后增的趋势, 三大科植物占比由撂荒10 a的66.67%上升至撂荒45 a的86.67%(表 2), 其中菊科植物占比随着年限的增加呈减少趋势.相对于土壤养分, 酶活性对三大科变化的响应更为敏感.草地群落中三大科植被的增减也可作为判断在撂荒后植被恢复过程中土壤养分变化的一个重要指标, 通过地上植被的调查间接了解或预测地下土壤变化状况.
| [1] | Sun W Y, Song X Y, Mu X M, et al. Spatiotemporal vegetation cover variations associated with climate change and ecological restoration in the Loess Plateau[J]. Agricultural and Forest Meteorology, 2015, 209-210: 87-99. DOI:10.1016/j.agrformet.2015.05.002 |
| [2] | Bai J J, Di L P, Bai J T. NDVI and regional climate variation since the implementation of revegetation program in northern Shaanxi Province, China[J]. IEEE Journal of Selected Topics in Applied Earth Observations and Remote Sensing, 2014, 7(11): 4581-4588. DOI:10.1109/JSTARS.2014.2365590 |
| [3] |
崔晓临, 雷刚, 王涛, 等. 退耕还林还草工程实施对洛河流域土壤侵蚀的影响[J]. 水土保持研究, 2016, 23(5): 68-73. Cui X L, Lei G, Wang T, et al. Impacts of grain for green project on soil erosion in Luohe River Basin of northern Shaanxi Province, China[J]. Research of Soil and Water Conservation, 2016, 23(5): 68-73. |
| [4] |
张晓霞, 杨宗儒, 查同刚, 等. 晋西黄土区退耕还林22年后林地土壤物理性质的变化[J]. 生态学报, 2017, 37(2): 416-424. Zhang X X, Yang Z R, Zha T G, et al. Changes in the physical properties of soil in forestlands after 22 years under the influence of the Conversion of Cropland into Farmland Project in Loess region, western Shanxi Province[J]. Acta Ecologica Sinica, 2017, 37(2): 416-424. |
| [5] |
邓景成, 高鹏, 穆兴民, 等. 黄土高原退耕还林工程对生态环境的影响及对策建议[J]. 水土保持研究, 2017, 24(5): 63-68. Deng J C, Gao P, Mu X M, et al. Impacts and advice of the grain for green project to ecological environment on the Loess Plateau[J]. Research of Soil and Water Conservation, 2017, 24(5): 63-68. |
| [6] |
雒明伟, 毛亮, 李倩倩, 等. 青藏高原筑路取土迹地恢复植物群落与土壤的碳氮磷化学计量特征[J]. 生态学报, 2015, 35(23): 7832-7841. Luo M W, Mao L, Li Q Q, et al. C, N, and P stoichiometry of plant and soil in the restorable plant communities distributed on the Land Used for Qinghai-Tibet Highway Construction in the Qinghai-Tibetan Plateau, China[J]. Acta Ecologica Sinica, 2015, 35(23): 7832-7841. |
| [7] | Korb J E, Johnson N C, Covington W W. Slash pile burning effects on soil biotic and chemical properties and plant establishment:recommendations for amelioration[J]. Restoration Ecology, 2004, 12(1): 52-62. DOI:10.1111/rec.2004.12.issue-1 |
| [8] |
刘洋, 曾全超, 安韶山, 等. 黄土丘陵区草本植物叶片与枯落物生态化学计量学特征[J]. 应用生态学报, 2017, 28(6): 1793-1800. Liu Y, Zeng Q C, An S S, et al. Ecological stoichiometry of leaf and litter of herbaceous plants in loess hilly and gully regions, China[J]. Chinese Journal of Applied Ecology, 2017, 28(6): 1793-1800. |
| [9] |
李鑫, 曾全超, 安韶山, 等. 黄土高原纸坊沟流域不同植物叶片及枯落物的生态化学计量学特征研究[J]. 环境科学, 2015, 36(3): 1084-1091. Li X, Zeng Q C, An S S, et al. Ecological stoichiometric characteristics in leaf and litter under different vegetation types of Zhifanggou Watershed on the Loess Plateau, China[J]. Environmental Science, 2015, 36(3): 1084-1091. |
| [10] | Horodecki P, Jagodziński A M. Tree species effects on litter decomposition in pure stands on afforested post-mining sites[J]. Forest Ecology and Management, 2017, 406: 1-11. DOI:10.1016/j.foreco.2017.09.059 |
| [11] |
赵勇, 吴明作, 樊巍, 等. 太行山针、阔叶森林凋落物分解及养分归还比较[J]. 自然资源学报, 2009, 24(9): 1616-1624. Zhao Y, Wu M Z, Fan W, et al. Comparison of nutrient return and litter decomposition between coniferous and broad-leaved forests in Hilly Region of Taihang Mountains[J]. Journal of Natural Resources, 2009, 24(9): 1616-1624. DOI:10.3321/j.issn:1000-3037.2009.09.011 |
| [12] |
曾昭霞, 刘孝利, 王克林, 等. 桂西北喀斯特区原生林与次生林凋落物及养分归还特征比较[J]. 生态环境学报, 2010, 19(1): 146-151. Zeng Z X, Liu X L, Wang K L, et al. Comparison of litterfall and nutrients return properties of primary and secon-dary forest ecosystems, the karst region of Northwest Guangxi[J]. Ecology and Environmental Sciences, 2010, 19(1): 146-151. DOI:10.3969/j.issn.1674-5906.2010.01.027 |
| [13] | Ren C J, Chen J, Deng J, et al. Response of microbial diversity to C:N:P stoichiometry in fine root and microbial biomass following afforestation[J]. Biology and Fertility of Soils, 2017, 53(4): 457-468. DOI:10.1007/s00374-017-1197-x |
| [14] |
何兵, 李睿玉, 张红, 等. 5种外来菊科植物对根际土壤微生物群落和酶活性的影响[J]. 安全与环境学报, 2013, 13(3): 176-180. He B, Li R Y, Zhang H, et al. Effects of five alien Compositae plants on the growth of the microbial community of the rhizosphere and the soil's enzyme activity[J]. Journal of Safety and Environment, 2013, 13(3): 176-180. DOI:10.3969/j.issn.1009-6094.2013.03.040 |
| [15] | An S S, Cheng Y, Huang Y M, et al. Effects of revegetation on soil microbial biomass, enzyme activities, and nutrient cycling on the Loess Plateau in China[J]. Restoration Ecology, 2013, 21(5): 600-607. DOI:10.1111/rec.2013.21.issue-5 |
| [16] | Perkins L B, Nowak R S. Native and non-native grasses generate common types of plant-soil feedbacks by altering soil nutrients and microbial communities[J]. Oikos, 2013, 122(2): 199-208. DOI:10.1111/more.2013.122.issue-2 |
| [17] |
薛超玉, 焦峰, 张海东, 等. 黄土丘陵区弃耕地恢复过程中土壤与植物恢复特征[J]. 草业科学, 2016, 33(3): 368-376. Xue C Y, Jiao F, Zhang H D, et al. The characteristics of plant species and soil quality in the restoration process of the Loess Hilly Region[J]. Pratacultural Science, 2016, 33(3): 368-376. |
| [18] |
刘海威, 张少康, 焦峰. 黄土丘陵区不同退耕年限草地群落特征及其土壤水分养分效应[J]. 草业学报, 2016, 25(10): 31-39. Liu H W, Zhang S K, Jiao F. Relationships between community characteristics and soil nutrients and moisture in abandoned hill country grassland[J]. Acta Prataculturae Sinica, 2016, 25(10): 31-39. DOI:10.11686/cyxb2016175 |
| [19] |
杨丽霞, 陈少锋, 安娟娟, 等. 陕北黄土丘陵区不同植被类型群落多样性与土壤有机质、全氮关系研究[J]. 草地学报, 2014, 22(2): 291-298. Yang L X, Chen S F, An J J, et al. Relationships among community diversity and soil organic matter, total nitrogen under different vegetation types in the Gully Region of Loess Region[J]. Acta Agrectir Sinica, 2014, 22(2): 291-298. |
| [20] | Zhang C, Liu G B, Xue S, et al. Soil bacterial community dynamics reflect changes in plant community and soil properties during the secondary succession of abandoned farmland in the Loess Plateau[J]. Soil Biology and Biochemistry, 2016, 97: 40-49. DOI:10.1016/j.soilbio.2016.02.013 |
| [21] |
杜峰, 徐学选, 张兴昌, 等. 陕北黄土丘陵区撂荒群落排序及演替[J]. 生态学报, 2008, 28(11): 5418-5427. Du F, Xu X X, Zhang X C, et al. The ordination of abandoned old-field communities and secondary succession rate, successional divergence or convergence in the Loess Hilly Region of northern Shaanxi Province[J]. Acta Ecologica Sinica, 2008, 28(11): 5418-5427. DOI:10.3321/j.issn:1000-0933.2008.11.025 |
| [22] |
张海东, 汝海丽, 焦峰, 等. 黄土丘陵区退耕时间序列梯度上草本植被群落与土壤C、N、P、K化学计量学特征[J]. 环境科学, 2016, 37(3): 1128-1138. Zhang H D, Ru H L, Jiao F, et al. C, N, P, K stoichiometric characteristic of leaves, root and soil in different abandoned years in Loess Plateau[J]. Environmental Science, 2016, 37(3): 1128-1138. |
| [23] |
秦伟, 朱清科, 张宇清, 等. 陕北黄土区生态修复过程中植物群落物种多样性变化[J]. 应用生态学报, 2009, 20(2): 403-409. Qin W, Zhu Q K, Zhang Y Q, et al. Dynamics of plant community species diversity in the process of ecological rehabilitation in North Shaanxi Loess Area[J]. Chinese Journal of Applied Ecology, 2009, 20(2): 403-409. |
| [24] |
张学礼, 胡振琪, 初士立. 土壤含水量测定方法研究进展[J]. 土壤通报, 2005, 36(1): 118-123. Zhang X L, Hu Z Q, Chu S L. Methods for measuring soil water content:a review[J]. Chinese Journal of Soil Science, 2005, 36(1): 118-123. DOI:10.3321/j.issn:0564-3945.2005.01.034 |
| [25] | 鲍士旦. 土壤农化分析[M]. 第三版. 北京: 中国农业出版社, 2000: 25-78. |
| [26] |
张超, 刘国彬, 薛萐, 等. 黄土丘陵区不同林龄人工刺槐林土壤酶演变特征[J]. 林业科学, 2010, 46(12): 23-29. Zhang C, Liu G B, Xue S, et al. Evolution of soil enzyme activities of Robinia pseudoacacia plantation at different ages in Loess Hilly Region[J]. Scientia Silvae Sinicae, 2010, 46(12): 23-29. DOI:10.11707/j.1001-7488.20101204 |
| [27] | Ren C J, Kang D, Wu J P, et al. Temporal variation in soil enzyme activities after afforestation in the Loess Plateau, China[J]. Geoderma, 2016, 282: 103-111. DOI:10.1016/j.geoderma.2016.07.018 |
| [28] | 马克平. 生物群落多样性的测度方法Ⅰα多样性的测度方法(上)[J]. 生物多样性, 1994, 2(3): 162-168. DOI:10.3321/j.issn:1005-0094.1994.03.007 |
| [29] | 马克平, 刘玉明. 生物群落多样性的测度方法Ⅰα多样性的测度方法(下)[J]. 生物多样性, 1994, 2(4): 231-239. DOI:10.3321/j.issn:1005-0094.1994.04.009 |
| [30] |
林勇, 艾训儒, 姚兰, 等. 木林子自然保护区不同群落类型主要优势种群的生态位研究[J]. 自然资源学报, 2017, 32(2): 223-234. Lin Y, Ai X R, Yao L, et al. Niches of main dominant species of different community types in Mulinzi Nature Reserve[J]. Journal of Natural Resources, 2017, 32(2): 223-234. |
| [31] |
郝文芳, 杜峰, 陈小燕, 等. 黄土丘陵区天然群落的植物组成、植物多样性及其与环境因子的关系[J]. 草地学报, 2012, 20(4): 609-615. Hao W F, Du F, Chen X Y, et al. Composition and diversity analysis of natural-community plants in the Loess Hilly Region[J]. Acta Agrestia Sinica, 2012, 20(4): 609-615. |
| [32] |
张超, 董淑琦, 刘国彬, 等. 黄土丘陵区植被演替中不同草本植物间的化感效应[J]. 草地学报, 2012, 20(5): 848-854. Zhang C, Dong S Q, Liu G B, et al. Allelopathic effect of different herbaceous plants during vegetation succession in the Hilly-Gully Region of Loess Plateau[J]. Acta Agrestia Sinica, 2012, 20(5): 848-854. |
| [33] |
张仁懿, 徐当会, 陈凌云, 等. 基于N:P化学计量特征的高寒草甸植物养分状况研究[J]. 环境科学, 2014, 35(3): 1131-1137. Zhang R Y, Xu D H, Chen L Y, et al. Plant N status in the alpine grassland of the Qinghai-Tibet Plateau:base on the N:P stoichiometry[J]. Environmental Science, 2014, 35(3): 1131-1137. |
| [34] | Van Ruijven J, Berendse F. Positive effects of plant species diversity on productivity in the absence of legumes[J]. Ecology Letters, 2003, 6(3): 170-175. DOI:10.1046/j.1461-0248.2003.00427.x |
| [35] |
王冬, 杨政, 郝红敏, 等. 黄土区退耕草地凋落物-土壤界面水分过程特征研究[J]. 水土保持研究, 2015, 22(1): 80-84. Wang D, Yang Z, Hao H M, et al. Hydrological processes of litter-soil interfacial systems in restoration grasslands of the Loess Plateau, China[J]. Research of Soil and Water Conservation, 2015, 22(1): 80-84. |
| [36] |
苏日古嘎, 张玉霞, 朱爱民.干旱胁迫对3种野生豆科牧草生长发育和抗旱生理机制的影响[J].内蒙古民族大学学报(自然科学版), 2017, 32(5): 424-428, 441. Suriguga, Zhang Y X, Zhu A M. Effects of drought stress on growth development and physiological mechanisms of drought resistance of 3 wild Leguminous forages[J]. Journal of Inner Mongolia University for Nationalities (Natural Sciences), 2017, 32(5): 424-428, 441. |
| [37] |
李磊, 贾志清, 朱雅娟, 等. 我国干旱区植物抗旱机理研究进展[J]. 中国沙漠, 2010, 30(5): 1053-1059. Li L, Jia Z Q, Zhu Y J, et al. Research advances on drought resistance mechanism of plant species in arid area of China[J]. Journal of Desert Research, 2010, 30(5): 1053-1059. |
| [38] |
周凯, 郭维明, 徐迎春. 菊科植物化感作用研究进展[J]. 生态学报, 2004, 24(8): 1776-1784. Zhou K, Guo W M, Xu Y C. Advances of research on allelopathic potential in compositae[J]. Acta Ecologica Sinica, 2004, 24(8): 1776-1784. DOI:10.3321/j.issn:1000-0933.2004.08.027 |
| [39] |
郭曼, 郑粉莉, 和文祥, 等. 黄土丘陵区不同退耕年限植被多样性变化及其与土壤养分和酶活性的关系[J]. 土壤学报, 2010, 47(5): 979-986. Guo M, Zheng F L, He W X, et al. Variation of vegetation diversity and its relationship with soil nutrient and enzyme activity in lands of different abandoned years in the loess hilly-gully region[J]. Acta Pedologica Sinica, 2010, 47(5): 979-986. |
| [40] |
崔宁洁, 张丹桔, 刘洋, 等. 不同林龄马尾松人工林林下植物多样性与土壤理化性质[J]. 生态学杂志, 2014, 33(10): 2610-2617. Cui N J, Zhang D J, Liu Y, et al. Plant diversity and soil physicochemical properties under different aged Pinus massoniana plantations[J]. Chinese Journal of Ecology, 2014, 33(10): 2610-2617. |
| [41] |
刘旻霞. 亚高寒草甸不同坡向植物光合生理和叶片形态差异[J]. 生态学报, 2017, 37(24): 8526-8536. Liu M X. Studies on physiological and leaf morphological traits for photosynthesis on different slopes in a subalpine meadow[J]. Acta Ecologica Sinica, 2017, 37(24): 8526-8536. |
| [42] | Zhong Y Q W, Yan W M, Wang R W, et al. Differential responses of litter decomposition to nutrient addition and soil water availability with long-term vegetation recovery[J]. Biology and Fertility of Soils, 2017, 53(8): 939-949. DOI:10.1007/s00374-017-1242-9 |
| [43] |
宋彦涛, 周道玮, 李强, 等. 松嫩草地80种草本植物叶片氮磷化学计量特征[J]. 植物生态学报, 2012, 36(3): 222-230. Song Y T, Zhou D W, Li Q, et al. Leaf nitrogen and phosphorus stoichiometry in 80 herbaceous plant species of Songnen grassland in Northeast China[J]. Chinese Journal of Plant Ecology, 2012, 36(3): 222-230. |
| [44] |
杨珏, 阮晓红. 土壤磷素循环及其对土壤磷流失的影响[J]. 土壤与环境, 2001, 10(3): 256-258. Yang Y, Ruan X H. Soil circulation of phosphosrus and its effects on the soil loss of phosphorus[J]. Soil and Environmental Sciences, 2001, 10(3): 256-258. DOI:10.3969/j.issn.1674-5906.2001.03.024 |
| [45] | Fan H B, Wu J P, Liu W F, et al. Linkages of plant and soil C:N:P stoichiometry and their relationships to forest growth in subtropical plantations[J]. Plant and Soil, 2015, 392(1-2): 127-138. DOI:10.1007/s11104-015-2444-2 |
| [46] | Zhang W, Qiao W J, Gao D X, et al. Relationship between soil nutrient properties and biological activities along a restoration chronosequence of Pinus tabulaeformis plantation forests in the Ziwuling Mountains, China[J]. CATENA, 2018, 161: 85-95. DOI:10.1016/j.catena.2017.10.021 |
| [47] |
刘善江, 夏雪, 陈桂梅, 等. 土壤酶的研究进展[J]. 中国农学通报, 2011, 27(21): 1-7. Liu S J, Xia X, Chen G M, et al. Study progress on functions and affecting factors of soil enzymes[J]. Chinese Agricultural Science Bulletin, 2011, 27(21): 1-7. |
| [48] |
刘洋, 黄懿梅, 曾全超. 黄土高原不同植被类型下土壤细菌群落特征研究[J]. 环境科学, 2016, 37(10): 3931-3938. Liu Y, Huang Y M, Zeng Q C. Soil bacterial communities under different vegetation types in the Loess Plateau[J]. Environmental Science, 2016, 37(10): 3931-3938. |
| [49] |
李君剑, 刘峰, 周小梅. 矿区植被恢复方式对土壤微生物和酶活性的影响[J]. 环境科学, 2015, 36(5): 1836-1841. Li J J, Liu F, Zhou X M. Effects of different reclaimed scenarios on soil microbe and enzyme activities in mining areas[J]. Environmental Science, 2015, 36(5): 1836-1841. |
| [50] | Ren C J, Chen J, Lu X J, et al. Responses of soil total microbial biomass and community compositions to rainfall reductions[J]. Soil Biology and Biochemistry, 2018, 116: 4-10. DOI:10.1016/j.soilbio.2017.09.028 |
| [51] |
张笑培, 杨改河, 任广鑫, 等. 黄土高原南部植被恢复对土壤理化性状与土壤酶活性的影响[J]. 干旱地区农业研究, 2010, 28(6): 64-68. Zhang X P, Yang G H, Ren G X, et al. Effects of vegetation restoration on soil physical-chemical properties and activities of soil enzyme in the gully region of the Loess Plateau[J]. Agricultural Research in the Arid Areas, 2010, 28(6): 64-68. |
| [52] | Siebenkäs A, Roscher C. Functional composition rather than species richness determines root characteristics of experimental grasslands grown at different light and nutrient availability[J]. Plant and Soil, 2016, 404(1-2): 399-412. DOI:10.1007/s11104-016-2853-x |
| [53] | Hu G Z, Liu H Y, Yin Y, et al. The role of legumes in plant community succession of degraded grasslands in northern China[J]. Land Degradation & Development, 2016, 27(2): 366-372. |