 2018, Vol. 39
2018, Vol. 39



作为生物膜法处理技术之一, 生物滤池在污水处理领域已有不少应用, 如工厂化水产养殖用到的各种生物包[1], 低污染源水的深度处理[2], 以及与其它水处理工艺的耦合[3]等.在城市污水处理方面, 生物滤池常用于污水深度脱氮, 一般是强化硝化-反硝化, 对应的构筑物有曝气生物滤池、反硝化滤池等.曝气生物滤池是目前研究和应用较多的一种, 有关其净化机制、效能评价、参数优化、技术开发等研究较多[4].然而持续曝气的生物滤池不仅耗能高, 同时滤池内的好氧环境也不利于反硝化.为此, 研究者们又提出了间歇曝气生物滤池, 通过创造滤池内的交替好氧/厌氧(或缺氧)状态, 不仅节能, 还有利于同步脱氮除磷[5].
对于新启用的生物滤池而言, 挂膜所需时间长短会影响到实际生产应用.挂膜通常是指进行微生物适应性驯化, 定向培育或富集功能微生物.由于生物滤池的构筑方式、处理对象、运行环境条件等多样, 目前关于生物滤池的启动时间长短报道不一[2, 6~8].另外, 生物滤池的净化效率不仅受控于滤料种类、填充深度、水力负荷、污染负荷、环境条件等, 还与池形(或流态)密切相关, 如硝化型曝气滤池可设计成下行流, 通过水-气对流增加氧气传质效果; 反硝化滤池可设计成折流式, 减小死水区, 增加污染物与基质的充分接触, 延长停留时间.
生物滤池的净化作用主要依赖于功能微生物, 后者又与上述影响因素密切相关.然而目前从微生物层面深入剖析滤池工况调整与净化效率的量化关系及其机制的报道较少.为了探究曝气、挂膜周期(或生物膜发育成熟度)、池形(或流态)等改变对生物滤池净化效率的影响, 本文设计了4种不同运行工况的生物滤池, 即3组垂直流滤池(MAVF、NAVF、NVF)和一组折流式水平流滤池(BHF). 4组滤池在同一环境条件下同步处理模拟配制的生活污水, 通过各组滤池净化效率的比较及微生物群落结构解析, 明确滤池净化效率与上述影响因素的量化关系, 以期为实际生产应用提供理论指导.
1 材料与方法 1.1 试验系统设计与构建试验构建的4组生物滤池框架均为有机玻璃, 其中3组垂直流滤池规格一致, 即L×W×H为48 cm×48 cm×60 cm, 水平流滤池为100 cm×48 cm×48 cm, 水平流滤池内还设置了9个等间距平行交错的折流板(39 cm×0.9 cm×48 cm). 4组滤池内均填充孔径3~5 mm、孔隙率为0.433的多孔陶粒; 垂直流、水平流滤池填充深度分别为38、34 cm.曝气滤池安装了曝气系统, 具体在曝气滤池底部铺设PVC布气管, 布气管直径20 mm, 表面钻有间距5~8 mm、大小5~6 mm的出气孔; 布气管内穿插纳米微孔曝气管(内径×外径=10 mm×15 mm), 后者通过聚乙烯软管(内径×外径=10 mm×13 mm)与旋涡风机相连(风量:60 m3·h-1; 最大风压:10 kPa; 品牌:亚士霸; 型号:HG-250;产地:浙江台州).运行时, 污水经水泵抽提后通过垂直流滤池表面的布水管流入滤池内, 其中MAVF滤池出水流入BHF滤池, 而NAVF和NVF滤池出水直接外排.各组滤池底部或末端装有出水阀, 用于调控滤池排水(图 1).

|
图 1 垂直流和折流式水平流滤池的结构示意 Fig. 1 Schematic diagrams of the vertical-flow and baffled horizontal-flow biofilters |
本试验开始之前, MAVF、BHF滤池已于前期运行1 a, 主要用于处理模拟配制的高浓度养殖废水[9], 滤池内已形成成熟的生物膜; 而NAVF、NVF为新启用滤池.试验启动时, 于各组滤池内接种采自武汉某污水处理厂生化池末端的活性污泥, 每天接种1次, 连续接种3 d.过后, 再往滤池内引入模拟配制的生活污水进行挂膜/微生物驯化, 并开始采集数据, 期间MAVF、NAVF滤池进行曝气, 其余未曝气.生活污水的配制参考该污水厂沉砂池的出水.生活污水的配制方案如下:每升水含面粉、葡萄糖(含少量钠、钙、铁、锌等元素)、碳酸氢铵、磷酸二氢钾的质量依次为0.30、0.15、0.225 6和0.043 9 g.配制废水对应各项污染物的质量浓度见表 1.
|
|
表 1 试验模拟配制的废水组成1)/mg·L-1 Table 1 Composition of the simulated wastewater used for the experiment/mg·L-1 |
目前关于生物滤池启动时间长短尚无定论, 如张菊萍等[6]采用接种挂膜方式, 在平均水温为16.7℃时, 仅需17 d就可以实现曝气生物滤池成功挂膜; 王建华等[7]考察了进水是否含有机物对硝化型曝气生物滤池挂膜的影响, 发现进水不含有机物的滤池成功挂膜仅需18 d, 而进水含有机物的滤池成功挂膜需24 d; 端艳等[8]称悬浮陶粒曝气生物滤池处理城镇污水的自然挂膜启动时间为23 d; 王东等[2]比较了沸石和陶粒填料曝气生物滤池处理微污染水源水的低温启动特性, 在水温为10~14℃的启动条件下, 均可在30 d内完成接种挂膜.本研究将新滤池挂膜时间设定为一个月, 期间将各组垂直流滤池水力负荷设为0.05、0.10、0.18 m3·(m2·d)-1这3个水平, 每个水平持续运行10 d.
3组垂直流滤池每天间歇进水1次, 其中MAVF出水即为BHF进水.曝气滤池每天间歇曝气6 h, 即上午09:00~12:00、下午14:30~17:30;对应3种水力负荷的气水比约为1 200:1、600:1和300:1.水样采集点为滤池进、出水, 采样频率为每天1次.采用美国YSI多参数水质分析仪(型号:Pro Plus)现场测定温度(T)、溶解氧(DO)、氧化还原电位(ORP)、碱度(pH)、电导率(Cond)、比电导率(SC)、总溶解固体(TDS)、盐度(Sal)等在线参数.水样采集后, 按国家标准方法[10]测定COD、TN、NO3--N、NO2--N、TAN、TP、IP等指标, 其中COD为高锰酸盐指数. COD又分为总有机物(TCOD)和溶解性有机物(DCOD). DCOD为水样经0.45μm滤纸过滤后测定值, TCOD为直接测定值.
挂膜结束后, 采集各组滤池陶粒样品送杭州联川生物技术股份有限公司进行微生物检测, 具体是通过16S rDNA高通量测序方法完成微生物群落结构分析.测序平台为MiSeq, 采用细菌16S rDNA V3+V4区域通用引物:338F ACTCCTACGGGAGGC AGCAG和806R GGACTACHVGGGTWTCTAAT.下机原始数据利用overlap将双端数据进行拼接, 并进行质控、嵌合体过滤以获得高质量的有效数据, 随后对其进行97%的相似度聚类.为了降低假阳性率, 过滤singleton序列以获得最终的OTU丰度及代表序列.
1.3 数据分析生物滤池的净化效率除受污染负荷影响外, 还与滤料类型、填充深度密切相关, 后者会影响到构建成本.为了综合多方面因素, 本文采用去除率和一阶去除率常数k来评价滤池的净化效能, 即:
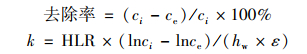
|
式中, k为一价去除率常数, d-1; HLR为水力负荷, m·d-1; ci、ce分别为进、出水污染物质量浓度, mg·L-1; hw为基质填充深度; ε为孔隙率.
采用独立t-test检验各组滤池进出水间理化特征的差异.因为BHF滤池进水即为MAVF出水, 为了屏蔽进水浓度对滤池净化效率的影响, 以进水污染负荷为协变量, 采用协方差分析比较各组滤池间净化效率的差异.此外, 为了比较各组滤池微生物群落结构及净化效率的相似性, 采用层次聚类法对优势菌种的相对丰度、所有OTU序列数及一阶去除率常数k进行聚类; 同时为了探究优势菌种与滤池出水理化环境特征及净化效率的关系, 还对这些变量进行了冗余度(redundancy analysis, RDA)排序分析.选择RDA是因为某些污染物指标的k值为负.排序、聚类分析分别在CANOCO 4.5、ORIGIN8.6软件中完成, 其它分析在SPSS 19.0软件中完成.
2 结果与讨论 2.1 不同运行工况滤池进出水理化特征比较由表 2可知, 4种不同工况生物滤池进出水理化参数间的差异主要体现在温度、溶解氧、氧化还原电位、碱度等指标上, 而电导率、比电导率、总溶解固体、盐度等指标的变化较小.另外, 与3组垂直流滤池相比, 水平流滤池进出水间的差异更小(仅氧化还原电位存在显著差异).进一步比较发现, 经垂直流滤池处理后, 出水温度显著升高, 溶解氧、pH显著降低.温度升高可能是因为曝气所致(鼓风机持续运行发热, 吹出的空气高于室温); 溶解氧、pH降低是因为滤池内部存在硝化过程, 需要消耗溶解氧和碱度.曝气显著提高了滤池出水溶解氧和pH(NVF出水的DO、pH分别与NAVF、MAVF出水的DO、pH相比, 所有P<0.05), 原因可能是因为硝化过程需要消耗CO2, 曝气能不断地向滤池内部输送O2和CO2, 而未曝气滤池硝化过程需要消耗原水中的CO32-或HCO3-, 后者致使pH降低[11].此外, 依据高立杰等[12]提出的DO等级划分方法, 即<0.3mg·L-1代表厌氧、0.4~0.7 mg·L-1代表兼氧及>1.0mg·L-1代表好氧, 本研究4组滤池内溶解氧含量处于好氧和兼氧水平.
|
|
表 2 4组滤池进出水理化参数比较1) Table 2 Comparison of the physicochemical parameters between inflow and outflow of the four biofilters |
2.2 不同运行工况滤池净化效能比较
由表 3可知, 4组滤池的出水硝氮、亚硝氮含量都很低(出水硝氮、亚硝氮的均值分别在0.40 mg·L-1和0.02 mg·L-1以下), 表明4组滤池内均无明显的硝氮、亚硝氮积累, 这也说明滤池内反硝化进行得很充分.相应地, 各组滤池出水中溶解性氮素以未被硝化的氨氮为主.此外, 通过比较滤池出水溶解性氮素和有机氮含量, 发现MAVF及BHF滤池出水总氮以有机氮为主, 而NAVF及NVF滤池出水总氮以溶解性氮素为主, 这可能是因为前两组滤池内生物膜相对成熟, 微生物数量丰富, 并由生物膜更新脱落所致.因此, 成熟滤池出水经沉淀处理后可进一步提高总氮去除效果.比较各组滤池的去除率, 发现3组垂直流滤池对有机物的去除率均较高, 其中TCOD的去除率都在80%以上, 但水平流滤池除对有机物/有机氮有一定的去除作用外, 对其它指标的去除率基本为负. BHF滤池对有机物、有机氮及总氮的去除率为正, 对氨氮、硝氮、亚硝氮及溶解性氮素的去除率为负, 表明滤池内发生了有机物的氨化、氨氮的硝化/亚硝化及反硝化等.同时, 由BHF滤池进出水各氮素指标的数值大小可知, BHF滤池内发生的上述氮素转化强度非常弱, 可能归结于BHF滤池内部较低的溶解氧水平.
|
|
表 3 4种不同工况滤池的进出水浓度、去除率及一阶去除率常数比较1) Table 3 Comparison of the inflow/outflow pollutant concentrations and their percent removal rate or k of the four biofilters operated under different working conditions |
3组垂直流滤池对硝氮都有较高的k值(表 3), 表明滤池内部反硝化作用较为明显, 可能是因为反硝化异养菌易于增殖, 滤池的间歇曝气运行没有对反硝化过程造成抑制.相反, 在水平流滤池中, 各项氮素指标的k值基本为负(总氮和有机氮除外), 表明水平流滤池内部的硝化-反硝化过程都受到了一定程度的抑制, 可能是因为水平流滤池没有曝气, 滤池内部的低溶解氧水平和相对不高的环境温度不利于硝化过程的进行, 进而不能为反硝化提供足够的反应底物.
方差分析结果显示, 不同工况滤池之间各项指标的k值都存在显著差异(表 3).多重比较发现, 4组滤池对碳、氮、磷的净化效率基本是MAVF>NAVF>NVF>BHF.在有机物去除方面, MAVF显著高于NAVF和NVF, 后者显著高于BHF. MAVF较NAVF的TCOD、DCOD的k值分别提高了35.2%、40.5%, 而NAVF较NVF的TCOD、DCOD的k值分别只提高了10.9%、13.3%.类似地, MAVF较NAVF的TP、IP、TAN、NO3--N、TN、DIN等k值分别提高了40.0%、33.3%、437.5%、22.8%、300.0%、385.2%, 而MAVF较NAVF的TP、IP、TAN、NO3--N、TN、DIN等k值分别只提高了25.0%、26.3%、100.0%、35.8%、120.0%、80.0%.这些增加值表明, 试验工况下滤池内碳、氮、磷的去除主要依赖于微生物的吸收和降解, 且生物膜成熟度对污染物去除的影响大于曝气.有例外曝气较生物膜成熟度对硝氮去除的影响更大, 可能是因为试验装置反硝化易于发生, 且主要受控于硝氮底物浓度, 而后者又主要来源于氨氮的氧化.此外, 各组滤池对磷的去除率均不高, 可能是因为滤池除磷仍然以生物除磷为主, 这与笔者之前报道的结果相似[9].
2.3 不同运行工况滤池微生物群落结构比较4种不同工况滤池在试验末期取陶粒样品进行微生物群落结构检测, 通过16S rDNA高通量测序分析共获得96 708条有效序列, 它们在4组滤池内的分布是:MAVF(16676)、NAVF(57367)、NVF(11161)和BHF(11504).这些有效序列经聚类后共获得2 260个OTU, 它们在4组滤池内的分布是:MAVF(680)、NAVF(691)、NVF(413)和BHF(828).再依据4组滤池所含OTU序列数绘制等级丰度图和相应的α多样性指数.由样品测序稀释曲线图 2(a)可知, Chao1曲线趋于平缓, 表明测序数据量已饱和; 由图 2(b)可知, 4组滤池的辛普森指数(Simpson)一致(均为0.98), 而观察种(Observed species)、香农(Shannon)、Chao1等指数存在明显差别, 顺序都是BHF>MAVF>NAVF>NVF, 表明滤池的成熟度与微生物多样性密切相关, 即滤池愈成熟, 多样性指数愈高.

|
图 2 4种不同工况滤池的稀释曲线、等级丰度图和α多样性指数 Fig. 2 Rarefaction curves, rank abundance curves, and α diversity index of the four biofilters under different working conditions |
依据测序获得的特征序列进行物种注释, 4组滤池共注释到341属种微生物, 其中MAVF、NAVF、NVF、BHF的物种数分别为212、200、161、257, 这与前面描述的多样性指数变化趋势一致.此外, 将所有物种的相对丰度均列出并不便于分析, 而且优势物种往往就能反映群落结构的主要特征.因此, 本文选取丰度最高的前20个物种或功能分类, 将其余的设置为others, 进行相对丰度计算, 并据此绘制相对丰度堆叠柱状图[图 3(a)和3(b)]和物种分类热图[图 3(c)和(d)], 便于直观进行样品间物种丰度的比较.

|
(a)和(c):门水平; (b)和(d):属水平 图 3 4种不同工况滤池的微生物相对丰度和物种分类热图 Fig. 3 Relative microbial abundance and taxa heatmap of the four biofilters under different working conditions |
由相对丰度堆叠柱状图可知, 在门水平上, 各组滤池的变形菌(Proteobacteria)丰度最高, 其次是拟杆菌(Bacteroidetes)和放线菌(Actinobacteria), 而厚壁菌(Firmicutes)仅在BHF中相对丰度较高.在属水平上, 相对丰度最高的是红环菌(Rhodocyclaceae_unclassified), 其余依次是黄杆菌(Flavobacterium)、纤维单胞菌(Cellulomonas)、β变形菌(β-proteobacteria_unclassified)、脱氯单胞菌(Dechloromonas)等.郭晓娅等[13]以玉米淀粉废水为反硝化碳源进行试验, 发现反应器内优势菌群中红环菌目相对丰度较大, 据此推测红环菌目可能较易于降解淀粉, 这与本研究以面粉(含淀粉)和葡萄糖为碳源进行脱氮除磷微生物富集相似.
从门水平的热图组成来看, NAVF和NVF的优势菌种组成最相似, 其中疣微菌(Verrucomicrobia)、拟杆菌、放线菌等丰度较高, 而Ignavibacteriae、绿弯菌(Chloroflexi)、硝化螺旋菌(Nitrospirae)、蓝细菌(Cyanobacteria)等丰度较低.相反, Ignavibacteriae、绿弯菌、硝化螺旋菌、蓝细菌等在BHF和MAVF中丰度较高.聚类结果显示, NAVF和NVF最相似, 而BHF与3组垂直流滤池差异最大.在属水平上, 聚类结果与门水平一致.同样, 4组滤池的优势菌种组成存在明显差别, BHF滤池中地杆菌(Geobacter)丰度最高, 而β变形菌、红环菌、伯克氏菌(Burkholderiales_unclassified)、沉积物杆状菌(Sediminibacterium)、芽殖杆菌(Gemmobacter)、脱氯单胞菌、热单胞菌(Thermomonas)等在MAVF中丰度较高.各组滤池的优势菌种相对丰度组成存在明显差别, 它们的代谢特征和主要功能注释见表 4.
传统观点认为, 硝化过程属于化能自养过程, 且严格好氧[26, 29].本研究通过物种注释, 于4四组滤池内仅检测到了3种专性硝化微生物, 即亚硝化单胞菌属(Nitrosomonas)、亚硝化螺菌属(Nitrosospira)及硝化螺菌属(Nitrospira), 且三者的相对丰度都很低(表 5).这与戴琦[30]、黄菲[31]等报道的硝化/亚硝化菌富集低丰度结果相似.戴琦等[30]采用膜生物反应器处理含环丙沙星(CIP)的模拟废水, 发现在0、5、10mg·L-1的CIP投加浓度下, 亚硝化单胞菌属的总相对丰度依次为0.02、0.01、0.006, 硝化螺菌属的总相对丰度依次为0.73、1.57、0.82;黄菲等[31]研究了在冬季低温条件下, 膜生物反应器与传统活性污泥法工艺运行效果及微生物群落特征的差异, 发现R1(高污泥浓度膜生物反应器)、R2(低污泥浓度膜生物反应器)、R3(传统活性污泥工艺)中主要的硝化菌为Nitrospira, 其总相对丰度依次为1.22%、1.64%、0.15%.将上述3种自养硝化菌与表 4中前20种优势菌种按代谢功能进行合并统计, 结果见表 5.从中可知, 4组滤池内丰度最高的微生物为异养反硝化脱氮菌, 其次为异养反硝化聚磷菌, 再次为异养硝化菌.因为很多异养反硝化脱氮菌兼为异养反硝化聚磷菌和(或)异养硝化菌(表 4), 因此, 三者丰度相对较高.
|
|
表 5 4组生物滤池中自养硝化菌及优势功能菌占细菌总丰度的百分比/% Table 5 Percentage of autotrophic nitrifiers and dominant functional bacteria of the total bacterial abundance among the four biofilters/% |
进一步分析自养硝化菌相对丰度数值, 发现NVF滤池缺乏自养硝化及亚硝化菌, BHF滤池内缺乏自养亚硝化菌, 而NAVF中缺乏自养硝化菌, 只有MAVF滤池同时含有硝化及亚硝化菌, 且MAVF滤池自养硝化菌(硝化+亚硝化菌)丰度明显高于其它3组滤池.这与2.2节观测到的MAVF滤池氨氧化作用最强, BHF滤池氨氧化作用最弱相一致. NVF及BHF滤池内的氨氧化过程均由异养菌完成, 且NVF滤池的氨氧化能力强于BHF滤池, 这是因为前者的异养硝化菌丰度明显高于后者(表 5).此外, 与两组曝气滤池(MAVF和NAVF)相比, 其余两组未曝气滤池(NVF和BHF)内均未检测到亚硝化菌, 说明曝气促进了滤池内氨氧化菌的富集.将表 5中各功能菌相对丰度与碳、氮、磷去除的k值进行相关性分析, 发现仅自养硝化菌(硝化+亚硝化菌)与氨氮、总氮及溶解性氮素之间存在显著正相关(P<0.05), Pearson相关系数分别为0.97、0.97和0.96, 这可能也暗示影响各种滤池脱氮的主要过程仍然为硝化过程, 且主要依赖于自养硝化菌.
2.4 滤池净化效能与微生物结构特征耦联分析如2.3节所述, 将获得的2260个OTU按各自所含的序列数进行样品聚类, 结果如图 4(a)所示; 同样地, 按测定的各项污染物指标k值进行样品聚类, 结果如图 4(b)所示.由图 4可知, 两种不同途径聚类结果与丰度最高的前20个优势物种热图聚类[图 3(c)和3(d)]具有一致性, 即丰度最高的前20个优势物种基本能涵盖所有物种的聚类信息.另外, 各组滤池净化效率聚类结果与物种聚类结果一致, 这也表明滤池之间净化效率的差异主要由滤池内微生物结构差异所致.为了进一步探讨滤池净化效率与微生物群落组成的关系, 将滤池的净化效率与图 3中优势物种组成以及后者与滤池出水理化特征进行多元排序分析, 结果如图 5所示.

|
图 4 基于4种滤池所有OTU和各项指标k值的层次聚类图 Fig. 4 Hierarchical cluster dendrogram based on all OTU and k values of the four biofilters |

|
图中数字1~20代表的物种见表 4 图 5 微生物优势菌种与滤池净化效率k、出水理化特征间的冗余度排序 Fig. 5 Ordination plots of redundancy analysis (RDA) between the dominant microorganisms and purification efficiency k/physicochemical characteristics of the outflow |
由各项污染物指标的k值与优势菌种的排序图 5(a)可知, 总氮、氨氮及溶解性氮素的去除具有一致性, 且与沉积物杆状菌、动胶菌(Zoogloea)、伯克氏菌、β变形菌、热单胞菌等正相关, 这是因为这些细菌兼有异养硝化和(或)反硝化功能(表 4), 而上述3种形态氮素的去除主要依赖于硝化-反硝化.沉积物杆状菌为兼性好氧异养菌, 在中性条件下的最佳生长温度为10~37℃[32]. Papirio等[26]以芬兰坦佩雷市某污水处理除厂活性污泥为接种物进行硝化菌的富集培养, 当流化床反应器的起始氨氮浓度为66.5~77.8mg·L-1、平均溶解氧为7.0mg·L-1时, 经过35d富集培养后于接种污泥及富集物中都检测到了优势菌属沉积物杆状菌的存在, 表明沉积物杆状菌可能具有硝化功能, 这与本研究结果相似.本研究中, 虽然沉积物杆状菌相对丰度不高(在MAVF、NAVF、NVF、BHF中的相对丰度分别为2.92、0.82、0.49、0.09), 但与自养硝化菌一样, 仍然发挥着一定的硝化功能.本研究中NVF滤池缺乏自养硝化菌, 但却发生了硝化作用, 证明了异养硝化菌的存在.此外, 排序图中有机物、磷及硝氮的去除也具有一致性, 表明磷主要通过反硝化聚磷方式去除.本研究通过16S rDNA高通量测序和物种注释并未检测到好氧聚磷菌.
由优势菌种与滤池出水的理化特征排序图 5(b)可知, 动胶菌、伯克氏菌、β变形菌、沉积物杆状菌、热单胞菌、Chitinophagaceae_unclassified、芽殖杆菌、丛毛单胞菌科(Comamonadaceae_unclassified)等与溶解氧或氧化还原电位正相关, 表明这些菌倾向于好氧环境, 与这些菌为兼性好氧菌相符(表 4).各组滤池出水总溶解固体、盐度、电导率及比电导率的变化具有一致性, 这是因为上述指标之间的关联性较强, 而且这些指标高低主要取决于出水中溶解性氮素高低(尤其是硝氮).此外, 由图 5(b)还可知, 上述4项指标与脱氯单胞菌、短波单胞菌(Brevundimonas)、简易螺旋菌(Simplicispira)、红环菌、黄杆菌、假单胞菌(Pseudomonas)、成对杆菌(Dyadobacter)、食酸菌(Acidovorax)、纤维单胞菌等正相关.据表 4, 这些优势菌属基本兼有反硝化脱氮功能, 它们与总溶解固体、盐度、电导率及比电导率的正相关, 表明反硝化强度主要取决于反应底物浓度.
3 结论(1) 配制原水经垂直流滤池处理后, 出水溶解氧、pH显著降低.曝气显著提高了滤池出水溶解氧和pH.与3组垂直流滤池相比, BHF滤池对进出水理化参数的影响更弱.本试验期间4组滤池内的溶解氧水平处于好氧和兼氧状态.
(2) 本试验工况下4组滤池内都无明显的硝氮、亚硝氮积累, 说明滤池内反硝化进行得很充分.各组滤池出水中溶解性氮素以未被硝化的氨氮为主, MAVF及BHF滤池出水总氮以有机氮为主, 而NAVF及NVF滤池出水总氮以溶解性氮素为主.垂直流滤池对碳、氮、磷的去除效率显著高于水平流滤池, 且水平流滤池内部发生的氮素转化过程非常微弱.各滤池内碳、氮、磷的去除主要依赖于微生物的吸收和降解, 且生物膜成熟度对污染物去除的影响大于曝气.
(3) 4组滤池的多样性指数高低是BHF>MAVF>NAVF>NVF, 表明滤池愈成熟, 多样性指数愈高.滤池之间净化效率的差异主要由滤池内微生物结构差异所致.各组滤池的优势菌种以兼性异养菌为主, 且以异养反硝化脱氮菌最为丰富. NVF滤池内缺乏自养硝化菌, 但发生了异养硝化过程; MAVF滤池内自养硝化菌相对丰度最高, 与其最强的硝化作用相呼应.曝气促进了滤池内亚硝化菌的富集, 进而增强了氨氧化能力. 4组滤池内均未检测到好氧聚磷菌, 磷的去除以反硝化聚磷为主.滤池对总氮的去除率不高主要归结于富集的自养硝化菌数量不高, 后者导致硝化作用较弱——这不仅引起出水中氨氮残余较高, 还会减少反硝化的底物浓度, 最终致使总氮去除率不高.
| [1] |
王峰, 雷霁霖, 高淳仁, 等. 国内外工厂化循环水养殖模式水质处理研究进展[J]. 中国工程科学, 2013, 15(10): 16-23, 32. Wang F, Lei J L, Gao C R, et al. Research progress of water conditioning in industry recirculating aquaculture mode at home and abroad[J]. Engineering Sciences, 2013, 15(10): 16-23, 32. DOI:10.3969/j.issn.1009-1742.2013.10.003 |
| [2] |
王东, 邱立平, 谢康, 等. 曝气生物滤池处理微污染水源水低温启动特性及效能研究[J]. 水处理技术, 2014, 40(8): 103-106, 110. Wang D, Qiu L P, Xie K, et al. Startup characteristics and efficiency of micro-polluted source water treatment by biologically aerated filter in low temperature[J]. Technology of Water Treatment, 2014, 40(8): 103-106, 110. |
| [3] |
钟丽燕, 郝瑞霞, 王卫东, 等. DNBF-O3-GAC组合工艺深度脱除氮磷及代谢产物[J]. 环境科学, 2018, 39(1): 247-255. Zhong L Y, Hao R X, Wang W D, et al. Combined process of DNBF-O3-GAC for nitrogen and phosphorus and metabolite advanced removal[J]. Environmental Science, 2018, 39(1): 247-255. |
| [4] |
朱加乐, 林燕, 王欣泽, 等. 曝气生物滤池脱氮的研究进展[J]. 化工进展, 2017, 36(3): 1077-1083. Zhu J L, Lin Y, Wang X Z, et al. Research progress on nitrogen removal by biological aerated filter[J]. Chemical Industry and Engineering Progress, 2017, 36(3): 1077-1083. |
| [5] |
孙成江.交替曝气两级生物滤池反硝化除磷工艺效能及其菌群结构研究[D].济南: 济南大学, 2015. Sun C J. Efficiency of denitrifying phosphorus removal and community structure in two stage biological filters operated by alternative aeration[D]. Ji'nan: University of Jinan, 2015. http://cdmd.cnki.com.cn/Article/CDMD-10427-1015443661.htm |
| [6] |
张菊萍, 陆少鸣, 夏莉, 等. 不同挂膜方式启动中置曝气生物滤池的对比研究[J]. 水处理技术, 2015, 41(6): 108-111. Zhang J P, Lu S M, Xia L, et al. Comparison of different forming film methods in biological aeration filter (BAF)[J]. Technology of Water Treatment, 2015, 41(6): 108-111. |
| [7] |
王建华, 陈永志, 彭永臻. 硝化型曝气生物滤池的挂膜与启动[J]. 环境工程学报, 2010, 4(10): 2199-2203. Wang J H, Chen Y Z, Peng Y Z. Biofilm formation and startup of nitrification biological aeration filter[J]. Chinese Journal of Environmental Engineering, 2010, 4(10): 2199-2203. |
| [8] | 端艳, 陆少鸣, 江荻. 悬浮陶粒曝气生物滤池在城镇污水处理中的挂膜启动[J]. 水处理技术, 2018(2): 88-92. |
| [9] |
张世羊, 张胜花, 张翔凌, 等. 组合生物滤池对养殖废水的净化效率及影响因素分析[J]. 环境科学, 2017, 38(6): 2419-2428. Zhang S Y, Zhang S H, Zhang X L, et al. Purification efficiency and influencing factors of combined bio-filters for aquaculture wastewater[J]. Environmental Science, 2017, 38(6): 2419-2428. |
| [10] | 国家环境保护总局. 水和废水监测分析方法[M]. 第四版. 北京: 中国环境科学出版社, 2002. |
| [11] | 张自杰, 张忠祥, 龙腾锐, 等. 废水处理理论与设计[M]. 北京: 中国建筑工业出版社, 2003. |
| [12] |
高立杰, 肖羽堂, 高冠道, 等. 微氧水处理技术的特性及应用研究进展[J]. 工业用水与废水, 2007, 38(2): 5-8. Gao L J, Xiao Y T, Gao G D, et al. Characteristics of micro-aerobic water treatment technology and its research progress[J]. Industrial Water & Wastewater, 2007, 38(2): 5-8. DOI:10.3969/j.issn.1009-2455.2007.02.002 |
| [13] |
郭晓娅, 年跃刚, 闫海红, 等. 以玉米淀粉废水为反硝化碳源的污染物降解特征与微生物群落结构研究[J]. 环境工程技术学报, 2016, 6(5): 427-433. Guo X Y, Nian Y G, Yan H H, et al. Pollutants degradation characteristics and microbial community structure using cornstarch wastewater as denitrification carbon source[J]. Journal of Environmental Engineering Technology, 2016, 6(5): 427-433. DOI:10.3969/j.issn.1674-991X.2016.05.063 |
| [14] |
张瑾.基于不同固体碳源生物反硝化性能及典型碳源生物膜特征研究[D].邯郸: 河北工程大学, 2018. Zhang J. Studies on denitrification performance and biofilm characteristics of the different solid carbon source-based denitrication system[D]. Handan: Hebei University of Engineering, 2018. 体碳源生物反硝化性能及典型碳源生物膜特征研 |
| [15] |
余鸿婷, 李敏. 反硝化聚磷菌的脱氮除磷机制及其在废水处理中的应用[J]. 微生物学报, 2015, 55(3): 264-272. Yu H T, Li M. Denitrifying and phosphorus accumulating mechanisms of denitrifying phosphorus accumulating organisms (DPAOs) for wastewater treatment-a review[J]. Acta Microbiologica Sinica, 2015, 55(3): 264-272. |
| [16] |
李鹏, 毕学军, 王军, 等. 常规和倒置A2/O工艺活性污泥微生物群落结构的比较[J]. 中国环境科学, 2017, 37(3): 1137-1145. Li P, Bi X J, Wang J, et al. Microbial diversity in activated sludges of conventional and reversed A2/O processes[J]. China Environmental Science, 2017, 37(3): 1137-1145. |
| [17] |
高惠玲, 袁其朋, 周延, 等. 两种碳源对纤维单胞菌生长及代谢的影响[J]. 北京化工大学学报, 2004, 31(6): 40-43. Gao H L, Yuan Q P, Zhou Y, et al. Effect of two carbon sources on growth and metabolism of Cellulomonas sp[J]. Journal of Beijing University of Chemical Technology, 2004, 31(6): 40-43. DOI:10.3969/j.issn.1671-4628.2004.06.010 |
| [18] | Foley M H, Martens E C, Koropatkin N M. SusE facilitates starch uptake independent of starch binding in B. thetaiotaomicron[J]. Molecular Microbiology, 2018, 108(5): 551-566. DOI:10.1111/mmi.13949 |
| [19] |
丁帮璟, 李正魁, 朱鸿杰, 等. 河岸带表层土壤的铁氨氧化(Feammox)脱氮机制的探究[J]. 环境科学, 2018, 39(4): 1833-1839. Ding B J, Li Z K, Zhu H J, et al. Insight into the mechanism of feammox in the surface soils of a riparian zone[J]. Environmental Science, 2018, 39(4): 1833-1839. |
| [20] | Hirsch P, Schlesner H. Gemmobacter[A]. In: Whitman W. Bergey's Manual of Systematics of Archaea and Bacteria[M]. John Wiley & Sons, Ltd., 2015. |
| [21] |
王思宇, 李军, 王秀杰, 等. 添加芽孢杆菌污泥反硝化特性及菌群结构分析[J]. 中国环境科学, 2017, 37(12): 4649-4656. Wang S Y, Li J, Wang X J, et al. Denitrification characteristics of Bacillus subtilis sludge and analysis of microbial community structure[J]. China Environmental Science, 2017, 37(12): 4649-4656. DOI:10.3969/j.issn.1000-6923.2017.12.030 |
| [22] |
戴琦, 刘锐, 梁玉婷, 等. 环丙沙星对膜生物反应器中微生物群落及抗性基因的影响[J]. 环境科学, 2018, 39(3): 1333-1341. Dai Q, Liu R, Liang Y T, et al. Influence of ciprofloxacin on the microbial community and antibiotics resistance genes in a membrane bioreactor[J]. Environmental Science, 2018, 39(3): 1333-1341. |
| [23] | Lee M, Woo S G, Park J, et al. Dyadobacter soli sp. nov., a starch-degrading bacterium isolated from farm soil[J]. International Journal of Systematic and Evolutionary Microbiology, 2010, 60(11): 2577-2582. DOI:10.1099/ijs.0.019653-0 |
| [24] | Mergaert J, Cnockaert M C, Swings J. Thermomonas fusca sp. nov. and Thermomonas brevis sp. nov., two mesophilic species isolated from a denitrification reactor with poly(ε-caprolactone) plastic granules as fixed bed, and emended description of the genus Thermomonas[J]. International Journal of Systematic and Evolutionary Microbiology, 2003, 53(6): 1961-1966. DOI:10.1099/ijs.0.02684-0 |
| [25] | Kavitha S, Selvakumar R, Sathishkumar M, et al. Nitrate removal using Brevundimonas diminuta MTCC 8486 from ground water[J]. Water Science & Technology, 2009, 60(2): 517-524. |
| [26] | Papirio S, Zou G, Ylinen A, et al. Effect of arsenic on nitrification of simulated mining water[J]. Bioresource Technology, 2014, 164: 149-154. DOI:10.1016/j.biortech.2014.04.072 |
| [27] |
王建超, 郝瑞霞, 孟成成, 等. 3DBER-S反硝化脱氮性能及其菌群特征[J]. 环境科学研究, 2015, 28(2): 310-317. Wang J C, Hao R X, Meng C C, et al. Study on performance and bacterial community of coupling 3-dimensional biofilm-electrode with sulfur autotrophic denitrification[J]. Research of Environmental Sciences, 2015, 28(2): 310-317. |
| [28] |
安卫星, 高娜, 夏明, 等. 动胶菌属系统分类、生理特征及其在活性污泥中的作用[J]. 应用与环境生物学报, 2016, 22(6): 1167-1174. An W X, Gao N, Xia M, et al. Physiological characteristics and systematic classification of the Zoogloea species and their role in the activated sludge[J]. Chinese Journal of Applied and Environmental Biology, 2016, 22(6): 1167-1174. |
| [29] | Vymazal J. Removal of nutrients in various types of constructed wetlands[J]. Science of the Total Environment, 2007, 380(1-3): 48-65. DOI:10.1016/j.scitotenv.2006.09.014 |
| [30] |
戴琦, 刘锐, 舒小铭, 等. 环丙沙星对膜生物反应器运行效能的影响及其去除特性[J]. 环境科学, 2018, 39(1): 212-218. Dai Q, Liu R, Shu X M, et al. Removal and influence of ciprofloxacin in a membrane bioreactor[J]. Environmental Science, 2018, 39(1): 212-218. |
| [31] |
黄菲, 梅晓洁, 王志伟, 等. 冬季低温下MBR与CAS工艺运行及微生物群落特征[J]. 环境科学, 2014, 35(3): 1002-1008. Huang F, Mei X J, Wang Z W, et al. Diversity of operation performance and microbial community structures in MBRs and CAS processes at low temperature[J]. Environmental Science, 2014, 35(3): 1002-1008. |
| [32] | Qu J H, Yuan H L. Sediminibacterium salmoneum gen. nov., sp. nov., a member of the phylum Bacteroidetes isolated from sediment of a eutrophic reservoir[J]. International Journal of Systematic and Evolutionary Microbiology, 2008, 58(9): 2191-2194. DOI:10.1099/ijs.0.65514-0 |