 2018, Vol. 39
2018, Vol. 39



厌氧氨氧化(anaerobic ammonium oxidation, ANAMMOX)工艺因其成本低、效率高引起人们的广泛关注.与传统硝化反硝化工艺相比, ANAMMOX工艺无需氧气和外加碳源, 并且污泥产量低[1, 2].然而许多废水如水产加工废水、畜禽养殖废水等都含有高浓度的氮和化学需氧量(COD), 这意味着单一ANAMMOX过程很难有效地处理废水.目前, 人们对厌氧氨氧化耦合反硝化工艺(SAD)的研究兴趣越来越浓厚[3].在有机碳存在的环境中, 厌氧氨氧化与反硝化反应能同步发生且相互促进. ANAMMOX菌(AnAOB)与反硝化菌(HDB)的协同作用可以同时去除有机物和氨氮.另外, SAD工艺还可以解决传统生物脱氮工艺外碳源不足的问题[4].
当前由于淡水资源短缺且水质下降, 海水的应用越来越受到人们的广泛关注, 尤其是在沿海地区[5].制革厂、食品、纺织工业以及渔业也都排放高盐度的废水[6, 7], 这不可避免地产生了大量的含盐废水.盐度对微生物代谢、污染物去除以及污泥沉降等方面均会产生负面影响[8, 9].微生物在高盐度条件下可以通过“相容性溶质”策略来调节渗透压以适应高盐环境[10].同时, 细胞内处于高离子强度时, 相容性溶质还可以作为蛋白质的稳定剂[11, 12].
海藻糖作为一种相容性溶质可以调节微生物渗透压, 添加海藻糖强化SAD工艺处理高盐废水鲜见报道.本研究通过按浓度梯度逐步提高海藻糖浓度, 探讨了海藻糖对SAD工艺处理高盐水脱氮除碳效能的影响, 并利用动力学分析了反应稳定运行的相关策略, 以期为强化SAD工艺处理高盐废水提供理论依据.
1 材料与方法 1.1 实验装置本实验所采用的SBR反应器如图 1所示.反应器是双层结构, 由有机玻璃制成, 有效容积为7 L, 其温度通过外室水循环控制在(35±0.5)℃.反应器用黑布包裹, 以防止光营养微生物的生长.该反应器运行一个周期包括进水0.5 h, 厌氧搅拌反应6 h, 污泥沉淀0.5 h和出水排水0.5 h.进水pH通过添加1 mol ·L-1的NaOH或HCl控制在7.4±0.2.海藻糖按浓度梯度0~0.6 mmol ·L-1逐步添加, 实验采用人工配制的模拟废水, 经高纯N2吹脱10 min以保持厌氧状态.

|
1.取样口及进出水口; 2.搅拌; 3.储水箱; 4.水浴进出水口; 5.水浴恒温控制器; 6.进水蠕动泵 图 1 厌氧氨氧化耦合反硝化装置示意 Fig. 1 Schematic diagram of the SBR configuration |
在本研究之前, 反应器已经运行了两年多, 具有长期稳定的脱氮性能.混合液悬浮固体(MLSS)的浓度和污泥沉降比(SV)分别为11.48 g ·L-1、11%.本实验采用的合成废水是由实际海水制备而来的.废水的盐度为32 g ·L-1, NH4+-N、NO2--N和COD分别由NH4Cl、NaNO2以及葡萄糖按进水80、160和214 mg ·L-1提供.实验所采用的合成废水成分为(g ·L-1): KH2PO4 0.029、CaCl2 ·2H2O 0.136、KHCO3 1.4、MgSO4 ·7H2O 0.06, 微量元素Ⅰ(1 mL ·L-1): EDTA 5 g ·L-1, FeSO4 ·7H2O 5 g ·L-1, 微量元素Ⅱ(1 mL ·L-1): EDTA 15 g ·L-1, H3BO3 0.014 g ·L-1, MnCl2 ·4H2O 0.99 g ·L-1, CuSO4 ·5H2O 0.25 g ·L-1, ZnSO4 ·7H2O 0.43 g ·L-1, NiCl2 ·6H2O 0.19 g ·L-1, Na2MoO4 ·2H2O 0.22 g ·L-1, CoCl2 ·6H2O 0.24 g ·L-1, NaSeO4 ·10H2O 0.21 g ·L-1.
1.3 分析项目和方法根据标准方法分析了NH4+-N、NO2--N、NO3--N和COD的浓度[13].污泥体积指数(SVI)和污泥沉降比(SV)也是基于标准方法进行测量的.对每个水样测3次取平均值. pH值: WTW pH/Oxi 340i;温度:水银温度计; 总氮去除速率(TNRR)和总氮去除率根据三氮进出水浓度和水力停留时间(HRT)得出, 如式(1)和(2)所示.

|
(1) |

|
(2) |
式中, HRT为水力停留时间(HRT=6 h, 采样间隔t=0.5 h), cinf为进水总氮浓度(mg ·L-1); ceff为出水总氮浓度(mg ·L-1).
厌氧氨氧化脱氮贡献率和反硝化脱氮贡献率如式(3)和(4)所示:
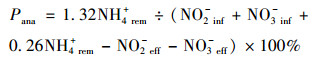
|
(3) |

|
(4) |
式中, 下标中ana、den、rem、inf、eff分别代表厌氧氨氧化、反硝化、去除率、进水和出水. NH4+、NO2-和NO3-的浓度以N(mg ·L-1)表示[14].
1.4 动力学模型Haldane模型被用来描述硝化过程中基质抑制[15]. Ewards模型[16]用来描述高浓度基质浓度抑制微生物的生长. Aiba模型[17]用来描述乙醇发酵过程中产物的抑制作用. Luong模型[18]用来描述丁醇抑制酵母的生长过程. Haldane模型和Aiba模型目前被广泛应用于拟合ANAMMOX反应器的基质抑制动力学特性[19, 20].在本研究中, 这4个模型用于描述海藻糖添加对SAD处理高盐废水脱氮过程影响的动力学特性.
(1) Haldane模型
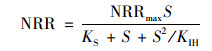
|
(5) |
式中, NRRmax为最大基质转化速率, kg ·(m3 ·d)-1; S为出水基质浓度, mg ·L-1; KS为半饱和常数, mg ·L-1; KIH为Haldane动力学常数, mg ·L-1.
(2) Ewards模型
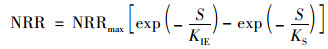
|
(6) |
式中, KIE为动力学Ewards常数, mg ·L-1.
(3) Aiba模型
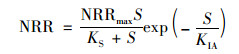
|
(7) |
式中, KIA为Luong动力学常数, mg ·L-1.
(4) Luong模型
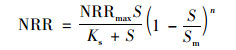
|
(8) |
式中, Sm为微生物净生长停止时的最大基质浓度, mg ·L-1; n为Luong动力学常数, mg ·L-1.
对于单周期内基质降解过程动力学模型, 不同的数学动力学模型运用于不同的生物反应器.修正的Logistic模型、修正的Boltzman模型和修正的Gompertz模型适合描述SBR反应器中ANAMMOX单周期内基质的降解过程.
(5) 修正的Logistic模型
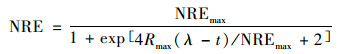
|
(9) |
把式(9)与式(2)合并:
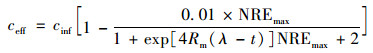
|
(10) |
式中, NRE为总氮去除率(%); NREmax为最大总氮去除率(%); Rmax为最大总氮去除速率(% ·h-1); λ为延迟时间(h); t为反应时间(h).
(6) 修正的Boltzman模型
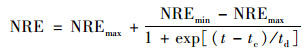
|
(11) |
把式(11)与式(2)合并:
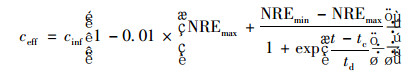
|
(12) |
式中, NREmin为最小总氮去除率(%); tc为1/2 NREmax时所对应的时间(h); td为时间常数(h).
(7) 修正的Gompertz模型
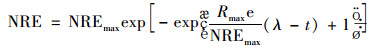
|
(13) |
把式(13)与式(2)合并:
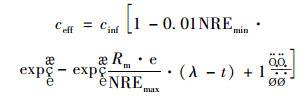
|
(14) |
海藻糖强化作用下的NH4+-N、NO2--N和NO3--N进出水浓度如图 2(a)所示.当海藻糖浓度≤0.25mmol ·L-1, 出水NH4+-N和出水NO2--N逐渐下降到0 mg ·L-1, NH4+-N和NO2--N去除率逐渐上升并最终达到100%.与没有添加海藻糖时相比, NH4+-N、NO2--N和总氮去除率分别提高了50%、43%和46%.在海藻糖浓度分别为0.05、0.1、0.15、0.2和0.25 mmol ·L-1时, 平均COD去除率分别为56.87%、68.03%、75.57%、87.00%和95.95%.以上结果表明0.25 mmol ·L-1海藻糖对于SAD工艺处理高盐废水效能的提高是最有效的, 反应器的脱氮效能最佳. Vyrides等[21]报道添加K+对缓解盐度抑制有积极的作用, 然而其效用相对于添加甜菜碱处于劣势.然而海藻糖浓度为0.5mmol ·L-1时, 平均出水NH4+-N浓度为33.25 mg ·L-1, 此时NH4+-N平均去除率为58.82%, 高于没有添加海藻糖时NH4+-N平均去除率(50.38%), 去除率高出8.44%.这一结果表明0.5 mmol ·L-1海藻糖还未对ANAMMOX产生抑制作用.马春[22]研究了在NaCl为10 g ·L-1条件下添加1mmol ·L-1甜菜碱出水NH4+-N和NO2--N减少.这一结果与本研究一致, 通过投加适量的海藻糖对于强化促进ANAMMOX和反硝化的协同作用是有效的.

|
(a)NH4+-N、NO2--N和NO3--N进出水浓度; (b)厌氧氨氧化与反硝化对氮去除的贡献率; (c)NH4+-N、NO2--N和COD负荷以及去除速率(d)化学计量比 图 2 不同海藻糖浓度下SAD的性能 Fig. 2 Performance of the simultaneous anammox and denitrification (SAD) over a long-term test of 248 cycles |
ANAMMOX和反硝化脱氮贡献率的变化规律如图 2(b)所示.当海藻糖在0~0.15 mmol ·L-1时, ANAMMOX脱氮贡献率在50%左右波动, 平均脱氮贡献率为53.01%, 与此同时, 反硝化脱氮贡献率跟ANAMMOX的变化趋势一致, 其平均脱氮贡献率为46.99%.此阶段平均COD去除率为64%.此阶段ANAMMOX与反硝化表现出良好的脱氮除碳的协同作用.当海藻糖浓度大于0.3 mmol ·L-1时, 反硝化脱氮贡献率大于ANAMMOX脱氮贡献率.当海藻糖浓度为0.5 mmol ·L-1时, 反硝化脱氮贡献率在57.94%~66.58%之间, ANAMMOX脱氮贡献率在33.42%~42.46%之间, 此时反硝化成为耦合反应中的主导反应.有报道指出[23]海藻糖能够被生物降解, 当海藻糖添加浓度超过ANAMMOX作为渗透调节剂的吸收浓度范围时, 多余的海藻糖反而会充当HDB的有机碳源.随着海藻糖的添加, 有机负荷(OLR)增加, AnAOB的活性降低而HDB的活性得到提升[24~26], 因而反硝化成为耦合反应中的主导反应.
NH4+-N去除速率(ARR)、NO2--N去除速率(NRR)、COD去除速率(ORR)的变化趋势如图 2(c)所示. ARR逐渐增加直到海藻糖浓度为0.25 mmol ·L-1, 此时平均ARR为0.29 kg ·(m3 ·d)-1, 在海藻糖浓度大于0.25 mmol ·L-1开始下降, 在浓度为0.5 mmol ·L-1时ARR为0.19 kg ·(m3 ·d)-1, 仍然高于没有添加海藻糖时ARR的值[0.16 kg ·(m3 ·d)-1]在最适浓度(0.25 mmol ·L-1)海藻糖添加下, ARR提高了81.25%, 0.5 mmol ·L-1海藻糖对ANAMMOX没有产生抑制作用. NRR随着海藻糖的添加而升高, 在0.25 mmol ·L-1时达到最大值[0.63 kg ·(m3 ·d)-1], 随后保持稳定. ORR随着海藻糖的添加而增加.与没有添加海藻糖相比, NRR提高了75%. Liu等[28]报道添加1 mmol ·L-1甜菜碱ANAMMOX反应中NRR和总氮去除率高于没有添加相容性溶质的值, 此外甜菜碱添加可以显著减少ANAMMOX在高盐条件下的驯化时间[28].这与Cyplik等[27]的研究结果相似, 该结果发现添加1 mmol ·L-1的四氢嘧啶能够加快反硝化反应进程.以上结果表明海藻糖能够促进ANAMMOX反应速率和反硝化反应速率.
ΔNO2--N/ΔNH4+-N(Ra)和ΔNO3--N/ΔNH4+-N(Rb)的变化趋势如图 2(d)所示.随着海藻糖的添加, Rb在整个反应过程基本稳定, 而Ra在整个反应过程中波动变化.文献[1]报道ANAMMOX反应ΔNO2--N/ΔNH4+-N和ΔNO3--N/ΔNH4+-N理论值分别为1.32和0.26. Ra和Rb分别高于和低于理论值.其原因可能是由于抑制条件下, 细胞的裂解会导致有机物和过量的氨释放, 随后反硝化会利用这一过程中产生的有机物质作为电子供体, 使得硝酸盐和亚硝酸盐减少[29].
2.2 海藻糖对SAD典型周期内脱氮除碳效能的影响典型单周期内氮和COD去除如图 3所示.当海藻糖浓度为0.05 mmol ·L-1时, NH4+-N、NO2--N和COD浓度随反应时间逐渐递减, NO3--N呈现微小的变化最后接近于0 mg ·L-1. NO2--N的转化和COD的降解变化几乎表现为同步.这一结果表明AnAOB和HDB菌具有良好的活性. pH值从7.44升高到8.03.海藻糖浓度为0.25 mmol ·L-1时, NH4+-N, NO2--N和COD在反应时间为5 h时几乎被完全去除.这一结果表明0.25 mmol ·L-1海藻糖为SAD处理高盐废水最适添加浓度. pH值从7.32升高到7.86.海藻糖浓度为0.4mmol ·L-1时, NO2--N和COD在反应时间为4 h完全去除, NH4+-N浓度随着反应时间逐渐减少, 在反应时间6 h末, 出水NH4+-N浓度为22.95 mg ·L-1. pH值从7.38升高到8.06而后又降低到7.7.海藻糖浓度为0.5 mmol ·L-1时, NO2--N和COD在反应时间为3.5 h时完全去除, 而NH4+-N在反应时间为3.5 h时减少到32.12 mg ·L-1, 在反应周期末却又升高到37.77 mg ·L-1. pH值从7.32升高到8.02, 随后又降到7.55.

|
图 3 不同海藻糖浓度下典型周期内氮去除过程 Fig. 3 Nitrogen removal in a typical SBR cycle with various influent trehalose concentrations |
当海藻糖浓度为0.4 mmol ·L-1和0.5 mmol ·L-1时, 单周期内pH值均出现了先升高然后在反应时间3.5 h后下降的现象. NO2--N消耗尽的点刚好为pH下降的点, NO2--N消耗尽时, ANAMMOX和反硝化反应停止, pH下降可能是由于NH4+和游离氨(FA)之间的化学平衡导致, OH-消耗使得反应器内pH值下降.另外, FA的形成会在一定程度上抑制ANAMMOX过程[25, 30], 从而使得pH下降. ANAMMOX由于没有NO2--N可利用, 使得反应器内的剩余NH4+-N不能被完全转化.出现上升的原因可能是因为SBR系统中存在其他形式的氮转化反应.文献[31]报道称AnAOB能够表现出异化作用即挥发性脂肪酸(VFA)氧化以硝酸盐作为中间产物将NO2--N转化为NH4+-N(DNRA).在批次实验中, 富集的ANAMMOX和自养菌可以表现出高效率的DNRA耦合ANAMMOX反应, 从而使得NH4+-N浓度上升.
本研究发现单周期内ΔpH和COD之间存在良好的线性关系(ΔpH=0.001 8COD-0.0486, R2=0.9921).这一结果与Li等[14]的研究相似.有报道指出, ANAMMOX更适应中性的pH条件[25].高浓度海藻糖抑制了ANAMMOX的活性, 一方面是由于海藻糖的降解使得反应器内有机碳源增加, 从而使得HDB活性提高, 在基质NO2--N的竞争中处于优势地位.另一方面, 高浓度海藻糖使得反应器内pH值升高, 这在一定程度也抑制了ANAMMOX的活性.
2.3 动力学分析出水基质浓度可作为评价反应器脱氮效能的一个重要的因素.在本研究中Haldane模型、Ewards模型、Aiba模型和Luong模型被用来拟合SAD脱氮过程动力学特性.模型拟合得到的结果如图 4所示. 4个模型均具有较高的R2, 这一结果显示4个模型均适合描述海藻糖添加条件下SAD处理高盐废水脱氮过程.从4个模型拟合的结果中可以得到最大基质转化速率的值, 但其相对误差各不相同. Haldane模型、Ewards模型、Aiba模型所得到的NRRmax的相对误差分别为27.94%、14.97%和9.09%, 而Luong模型得到的相对误差仅为4.15%, 这一结果表明Luong模型跟其他3个模型相比最适合描述海藻糖添加条件下SAD处理高盐废水的脱氮过程. Luong模型拟合所得到的各参数NRRmax、KS、Sm和n分别为0.954 kg ·(m3 ·d)-1、0 mg ·L-1、184.785 mg ·L-1和0.718.以上参数的获取为反应器的管理和控制, 以及反应器脱氮性能的分析等提供了较好的参考价值.

|
图 4 海藻糖添加下SAD全过程脱氮过程动力学模型曲线 Fig. 4 Curve of the whole process of SAD kinetic model with trehalose addition |
采用修正的Logistic模型、修正的Boltzman模型和修正的Gompertz模型对典型周期内的氮去除表现进行拟合, 拟合结果如图 5和表 1所示. 3个模型所得到的预测值与实验值之间的对比如表 2所示.对比3个模型所得到的参数, 均具有较高的R2.修正的Gompertz模型拟合得到的R2值的平均值比其他2个模型都高. 3个模型拟合得到的NREmax均接近于实验值.修正的Gompertz模型得到的预测值与实验值最为贴近.以上结果表明修正的Gompertz模型更适合描述海藻糖添加条件下单周期内基质的降解过程. Zhu等[32]用再次修正的Logistic模型拟合有机物存在时ANAMMOX脱氮过程并预测ANAMMOX脱氮效能, 结果表明不同有机物浓度下λ在0.69~1.93范围内变化, 表明NH4+-N去除的延迟.而在本研究中3个模型拟合得到的λ都几乎接近于0, 这一结果表明海藻糖添加条件下SAD对氮的去除没有延迟.当海藻糖浓度为0.15 mmol ·L-1时, 修正的Logistic模型和修正的Gompertz模型拟合所得到的λ分别为0.12 h和0.064 h.当海藻糖浓度不超过0.25 mmol ·L-1时, 修正的Logistic模型和修正的Gompertz模型拟合得到的Rmax随着海藻糖浓度的增加而增加, 且在海藻糖浓度为0.25 mmol ·L-1时达到最大值, 由此可以看出, ≤0.25 mmol ·L-1海藻糖能够促进SAD的脱氮效能. 3个模型拟合所得到的参数包括最大总氮去除率、单周期内最大氮去除速率, 并且能够预测出单周期内SAD脱氮延迟时间.这些参数的获取将为反应器的优化设计和反应器的运行调控提供参考.

|
图 5 不同单周期内氮去除过程动力学模型拟合曲线 Fig. 5 Curve of nitrogen removal process kinetic model at different trehalose concentrations |
|
|
表 1 不同海藻糖浓度SAD脱氮过程动力学拟合参数 Table 1 Parameters of SAD process kinetics at different trehalose concentrations |
|
|
表 2 单周期内模型拟合得到的预测值和实验数据之间的对比 Table 2 Comparison of predicted and experimental data for nitrogen removal at single circle |
3 结论
(1) 采用海藻糖可以强化SAD工艺处理高盐废水的脱氮除碳效能, 最适投加量为0.25 mmol ·L-1.在该条件下, 与没有添加海藻糖相比, NH4+-N、NO2--N和总氮去除率分别提高了50%、43%和46%, ARR和NRR分别提高了81.25%和75.00%.
(2) 相比于Haldane模型和Aiba模型, Luong模型更适合拟合海藻糖添加条件下SAD的脱氮性能.由其拟合得到的NRRmax、KS、Sm和n分别为0.954 kg ·(m3 ·d)-1、0 mg ·L-1、184.785 mg ·L-1和0.718.
(3) 与修正的Logistic模型和修正的Boltzman模型相比, 修正的Gompertz模型得到的预测值与实验值最为贴近, 修正的Gompertz模型更适合描述海藻糖添加条件下单周期内基质的降解过程.
| [1] | Strous M, Heijnen J J, Kuenen J G, et al. The sequencing batch reactor as a powerful tool for the study of slowly growing anaerobic ammonium-oxidizing microorganisms[J]. Applied Microbiology and Biotechnology, 1998, 50(5): 589-596. DOI:10.1007/s002530051340 |
| [2] | van der Star W R L, Abma W R, Blommers D, et al. Startup of reactors for anoxic ammonium oxidation:experiences from the first full-scale anammox reactor in Rotterdam[J]. Water Research, 2007, 41(18): 4149-4163. DOI:10.1016/j.watres.2007.03.044 |
| [3] | Takekawa M, Park G, Soda S, et al. Simultaneous anammox and denitrification (SAD) process in sequencing batch reactors[J]. Bioresource Technology, 2014, 174: 159-166. DOI:10.1016/j.biortech.2014.10.021 |
| [4] |
刘常敬, 李泽兵, 郑照明, 等. 苯酚对厌氧氨氧化工艺耦合反硝化的启动及脱氮性能的影响[J]. 中国环境科学, 2014, 34(5): 1145-1151. Liu C J, Li Z B, Zheng Z M, et al. Effect of phenol on startup and nitrogen removal performance of ANAMMOX coupling denitrifying[J]. China Environmental Science, 2014, 34(5): 1145-1151. |
| [5] | Vörösmarty C J, Mcintyre P B, Gessner M O, et al. Global threats to human water security and river biodiversity[J]. Nature, 2010, 467(7315): 555-561. DOI:10.1038/nature09440 |
| [6] | Belkin S, Brenner A, Abeliovich A. Biological treatment of a high salinity chemical industrial wastewater[J]. Water Science and Technology, 1993, 27(7): 105-112. |
| [7] | Dalmacija B, Karlovic E, Tamas Z, et al. Purification of high-salinity wastewater by activated sludge process[J]. Water Research, 1996, 30(2): 295-298. DOI:10.1016/0043-1354(95)00170-0 |
| [8] | Mannina G, Cosenza A, Trapani D D, et al. Membrane bioreactors for treatment of saline wastewater contaminated by hydrocarbons (diesel fuel):An experimental pilot plant case study[J]. Chemical Engineering Journal, 2016, 291: 269-278. DOI:10.1016/j.cej.2016.01.107 |
| [9] | Song W L, Li Z P, Ding Y, et al. Performance of a novel hybrid membrane bioreactor for treating saline wastewater from mariculture:Assessment of pollutants removal and membrane filtration performance[J]. Chemical Engineering Journal, 2018, 331: 695-703. DOI:10.1016/j.cej.2017.09.032 |
| [10] | Schubert T, Maskow T, Benndorf D H, et al. Continuous synthesis and excretion of the compatible solute ectoine by a transgenic, nonhalophilic bacterium[J]. Applied & Environmental Microbiology, 2007, 73(10): 3343-3347. |
| [11] | Oren A. Thermodynamic limits to microbial life at high salt concentrations[J]. Environmental Microbiology, 2011, 13(8): 1908-1923. DOI:10.1111/j.1462-2920.2010.02365.x |
| [12] | Müller V, Spanheimer R, Santos H. Stress response by solute accumulation in archaea[J]. Current Opinion in Microbiology, 2005, 8(6): 729-736. DOI:10.1016/j.mib.2005.10.011 |
| [13] | AP HA. Standard methods for the examination of water and wastewater (20th ed.)[J]. Washington DC:APHA/AWWA/WEF, 1998. |
| [14] | Li J, Qiang Z M, Yu D S, et al. Performance and microbial community of simultaneous anammox and denitrification (SAD) process in a sequencing batch reactor[J]. Bioresource Technology, 2016, 218: 1064-1072. DOI:10.1016/j.biortech.2016.07.081 |
| [15] | Sheintuch M, Tartakovsky B, Narkis N, et al. Substrate inhibition and multiple states in a continues nitrification process[J]. Water Research, 1995, 29(3): 953-963. DOI:10.1016/0043-1354(94)00216-T |
| [16] | Edwards V H. The influence of high substrate concentrations on microbial kinetics[J]. Biotechnology and Bioengineering, 1970, 12(5): 679-712. DOI:10.1002/(ISSN)1097-0290 |
| [17] | Aiba S, Shoda M, Nagatani M. Kinetics of product inhibition in alcohol fermentation[J]. Biotechnology and Bioengineering, 1968, 10(6): 845-864. DOI:10.1002/(ISSN)1097-0290 |
| [18] | Luong J H T. Generalization of monod kinetics for analysis of growth data with substrate inhibition[J]. Biotechnology and Bioengineering, 1987, 29(2): 242-248. DOI:10.1002/(ISSN)1097-0290 |
| [19] | Jin R C, Xing B S, Yu J J, et al. The importance of the substrate ratio in the operation of the Anammox process in upflow biofilter[J]. Ecological Engineering, 2013, 53: 130-137. DOI:10.1016/j.ecoleng.2012.12.027 |
| [20] | Chen T, Zheng P, Shen L, et al. Kinetic characteristics and microbial community of Anammox-EGSB reactor[J]. Journal of Hazardous Materials, 2011, 190(1-3): 28-35. DOI:10.1016/j.jhazmat.2010.12.060 |
| [21] | Vyrides I, Santos H, Mingote A, et al. Are compatible solutes compatible with biological treatment of saline wastewater? Batch and continuous studies using submerged anaerobic membrane bioreactors (SAMBRs)[J]. Environmental Science & Technology, 2010, 44(19): 7437-7442. |
| [22] |
马春. 厌氧氨氧化工艺处理低温和高盐度废水的可行性研究[D]. 杭州:杭州师范大学, 2012. Ma C. The feasibility study of anammxo process treating low temperature and high salinity wastewater[D]. Hangzhou:Hangzhou Normal University, 2012. |
| [23] | Vyrides I, Stuckey D C. Compatible solute addition to biological systems treating waste/wastewater to counteract osmotic and other environmental stresses:a review[J]. Critical Reviews in Biotechnology, 2017, 37(7): 865-873. DOI:10.1080/07388551.2016.1266460 |
| [24] | Kang J, Wang J L. Influence of chemical oxygen demand concentrations on anaerobic ammonium oxidation by granular sludge from EGSB reactor[J]. Biomedical and Environmental Sciences, 2006, 19(3): 192-196. |
| [25] | Waki M, Tokutomi T, Yokoyama H, et al. Nitrogen removal from animal waste treatment water by anammox enrichment[J]. Bioresource Technology, 2007, 98(14): 2775-2780. DOI:10.1016/j.biortech.2006.09.031 |
| [26] | Huang X L, Gao D W, Tao Y, et al. C2/C3 fatty acid stress on anammox consortia dominated by Candidatus Jettenia asiatica[J]. Chemical Engineering Journal, 2014, 253(7): 402-407. |
| [27] | Cyplik P, Piotrowska-Cyplik A, Marecik R, et al. Biological denitrification of brine:the effect of compatible solutes on enzyme activities and fatty acid degradation[J]. Biodegradation, 2012, 23(5): 663-672. DOI:10.1007/s10532-012-9542-0 |
| [28] | Liu M, Peng Y Z, Wang S Y, et al. Enhancement of anammox activity by addition of compatible solutes at high salinity conditions[J]. Bioresource Technology, 2014, 167: 560-563. DOI:10.1016/j.biortech.2014.06.015 |
| [29] | Dosta J, Fernández I, Vázquez-Padín J R, et al. Short-and long-term effects of temperature on the Anammox process[J]. Journal of Hazardous Materials, 2008, 154(1-3): 688-693. DOI:10.1016/j.jhazmat.2007.10.082 |
| [30] | Fernã n I, Dosta J, Fajardo C, et al. Short-and long-term effects of ammonium and nitrite on the Anammox process[J]. Journal of Environmental Management, 2012, 95(Suppl): S170-S174. |
| [31] | Castro-Barros C M, Jia M S, van Loosdrecht M C M, et al. Evaluating the potential for dissimilatory nitrate reduction by anammox bacteria for municipal wastewater treatment[J]. Bioresource Technology, 2017, 233: 363-372. DOI:10.1016/j.biortech.2017.02.063 |
| [32] | Zhu W Q, Zhang P Y, Dong H Y, Li J. Effect of carbon source on nitrogen removal in anaerobic ammonium oxidation (anammox) process[J]. Journal of Bioscience and Bioengineering, 2017, 123(4): 497-504. DOI:10.1016/j.jbiosc.2016.11.006 |