 2018, Vol. 39
2018, Vol. 39



采矿废水是由矿井排水、露天采场排水和废石场雨排水构成的一种酸性废水, 含有高浓度的硫酸盐和重金属等物质.人工湿地作为常见的废水处理方法之一, 在采矿废水的处理领域中已有一定的应用.湿地沉积物中氮含量与其迁移转化过程对湿地系统的结构、功能和生产力有重要影响, 氮含量被视为湿地营养水平指示剂, 是天然或者人工湿地沉积物中的主要限制性养分[1~4].采矿废水、酸性矿山排水、矿井排水等中缺乏氮磷等营养元素, 研究处理此类废水湿地中氮循环过程具有重要的意义.
湿地生态系统的氮素输入过程包括大气氮沉降、生物固氮、人为氮和径流氮输入等途径.对于流入湿地的无机氮, 首先将还原价态氨氮氧化成高价态氮, 例如氨氮被氧化成亚硝酸盐或硝酸盐, 再被还原成氮气.常见的厌氧氨氧化(ANAMMOX)广泛存在于河口、中国华南水稻土和天然淡水湿地等环境中[5~7].近年来, 厌氧氨氧化与铁还原耦合作用(Feammox)过程得到了国内外研究者的广泛关注, 研究对象包括美国新泽西州的河岸湿地、波多黎各的热带雨林土壤、中国稻田土壤和美国卡罗莱纳州萨凡纳河流流域等[8~13].然而, 目前对酸性矿山排水污染土壤及相关湿地中发生的Feammox过程尚缺乏公开报道.
凹山湿地处理的采矿废水含高浓度的SO42-, 沉积物中含有较高浓度的离子交换态铁, 以及湿地中有机质、总氮等营养物质流失较为严重[14].本实验以人工湿地为研究对象, 采集沉积物, 通过添加氨氮和水铁矿, 以及利用乙炔抑制剂的技术手段, 探究Feammox是否存在及其对氮转化过程的影响, 同时分析了水铁矿加入对湿地氨氮厌氧转化过程的影响, 达到了减少湿地系统总氮流失的目的.
1 材料与方法 1.1 沉积物取样地点和预处理考察用于处理采矿废水的安徽省马鞍山市凹山湿地, 并采集沉积物样品.样品迅速装入冰盒保鲜箱, 保鲜箱中预存保湿剂和脱氧剂, 并在6 h内送到合肥工业大学.在厌氧手套箱中沉积物样品经人工除杂后, 过220 mm筛子待用.将沉积物搅拌均匀后装入血清瓶, 加入体积比为1:3的去离子水, 高纯度氩气(99.99%)鼓气密封, 在黑暗32℃下预培养25 d, 以消耗本底氨氮、亚硝酸盐、硝酸盐和氧气[9, 11].预处理完后取样分析未发现氨氮、亚硝酸盐、硝酸盐.
1.2 培养实验在厌氧手套箱中将预处理的沉积物泥浆除去上清液, 分别称取10.0 g剩余泥浆(pH≈6.5)置于25个50.0 mL血清瓶中, 每瓶各加入30.0 mL营养液, 按照实验分组相应加入NH4Cl溶液(1 719.49 mg·L-1)和水铁矿, 添加氨氮反应瓶中初始氮含量为90.0 mg·kg-1.实验加入的营养液组成[13]为16.0 mg·L-1 KH2PO4; 300.0 mg·L-1 MgCl2·6H2O; 74.0 mg·L-1 KCl; 840.0 mg·L-1 NaHCO3; 200.0 mg·L-1 NaCl; 180.0 mg·L-1CaCl2·2H2O; 微量元素溶液1.0 mL·L-1.微量元素溶液组成为: 204.0 mg·L-1 ZnCl2; 240.0 mg·L-1 CoCl2·6H2O; 990.0 mg·L-1 MnCl2·4H2O; 149.0 mg·L-1 CuCl2·2H2O; 220.0 mg·L-1 Na2MoO4·2H2O; 190.0 mg·L-1 NiCl2·6H2O; 140.0 mg·L-1H3BO4·2H2O; 210.0 mg·L-1 NaSeO4·10H2O.
将25个血清瓶随机分为5组, 第一组为空白对照组, 加入2.0 mL去离子水; 第二组加入2.0 mL NH4Cl溶液; 第三组加入2.0 mL NH4Cl溶液, 上层气相含30.0%(~3 kPa)乙炔气体; 第四组加入2.0 mL NH4Cl溶液和41.24 mg水铁矿; 第五组加入2.0 mL NH4Cl溶液、41.24 mg水铁矿, 上层气相含30.0%(~3 kPa)乙炔气体.添加上述NH4Cl溶液和水铁矿后, 将所有血清瓶鼓氩气15 min(DO<0.10 mg·L-1)脱氧之后密封, 其中第三和第五组抽取血清瓶上层氩气6.0 mL, 添加等体积的高纯度纯乙炔(99.99%)气体, 体积比为30.0%(~3 kPa)[8].然后将所有血清瓶置于32℃培养箱避光下培养15 d, 反应瓶每1~2 d以100 r·min-1摇晃1.0 min.
1.3 测试方法氮气(N2)浓度采用鲁南气相色谱仪(SP 6890, 杭州瑞析科技有限公司)检测; 一氧化二氮浓度(N2O)采用傅里叶变换红外光谱仪(VERTEX 70, 德国)检测; 硫化氢气体(H2S)采用气相分子吸收光谱仪(GMA33xx, 上海北裕分析仪器有限公司, 上海)检测; pH值采用梅特勒-托利多Delt320型酸度计检测.
2 mol·L-1KCl溶液提取泥浆中的NH4+、NO2-和NO3-, 提取2.0 h后, 稀释5~10倍后采用气相分子吸收光谱仪(GMA33xx, 上海北裕分析仪器有限公司, 上海)检测; 0.5 mol·L-1HCl溶液提取泥浆中的Fe(Ⅱ)和Fe(Ⅲ), 用菲啰嗪法测定溶液中的Fe(Ⅱ)和Fe(Ⅲ)浓度[8]; 硫酸盐(SO42-)采用1+1 HCl溶液煮沸提取, 用离子色谱仪(皖仪, S600)测定的硫酸盐浓度; 硫化物浓度(S2-)采用气相色谱法(AJ-2100气相分子吸收光谱仪, 上海安杰环保科技有限公司)检测[15]; 单质硫(S)采用高效液相色谱仪(ULTIMATE 3000, USA)检测[15]; 重金属浓度采用火焰原子吸收法(WAYEE-WYS 2200)检测; TOC浓度采用重铬酸钾氧化法检测.
2 结果与讨论 2.1 不同体系的氮气(N2)产生五组反应体系氮气生成及五组硫酸盐还原过程结果如图 1所示.在空白对照组中氮气产生量为(0.80±0.05)mg·kg-1, 氮气产生速率为(0.05±0.003) mg·(kg·d)-1, 与其他三组对比可以忽略. NH4+组体系中氮气产生量为(63.17±5.34) mg·kg-1, 氮气产生速率为(4.21±0.36) mg·(kg·d)-1, 沉积物氮损失达到了(70.15±1.25)%. NH4++C2H2组体系中氮气产生量为(25.35±3.32) mg·kg-1, 氮气产生速率为(1.69±0.221) mg·(kg·d)-1, 沉积物氮损失达到了(27.05±2.01)%. NH4++Fe(Ⅲ)组体系中氮气产生量为(41.07±5.17) mg·kg-1, 氮气产生速率为(2.74±0.35) mg·(kg·d)-1, 沉积物氮损失达到了(45.44±0.98)%. NH4++Fe(Ⅲ)+C2H2组体系中氮气产生量为(38.18±1.23) mg·kg-1, 氮气产生速率为(2.55±0.08) mg·(kg·d)-1, 沉积物氮损失达到了(42.29±2.11)%.

|
图 1 培养15 d后反应体系中积累氮气产量和硫酸盐/硫离子含量 Fig. 1 Accumulative nitrogen production and sulfate/sulfide content in the reaction system after 15 days |
孙志高等[16]应用C2H2抑制原状土柱培育法研究湿地土壤反硝化速率, 发现反硝化速率变化为0.301~1.595 mg·(kg·d)-1, 分析温度因素因子与反硝化速率存在显著相关, 采用孙等的线性回归方程估算本实验反硝化速率为1.561~1.786 mg·(kg·d)-1, 未加C2H2抑制剂反应器氮气产生速率比此值高出53.42%~169.70%.据报道在沉积物、土壤、纯培养中, C2H2分压≥0.7 kPa时可完全抑制反硝化过程中N2O的还原[17, 18], 而ANAMMOX群落对C2H2的抑制作用更加敏感, 乙炔≥70 Pa时即被抑制95%[19], Feammox菌群对C2H2的敏感性较低[7].如图 1所示, NH4++Fe(Ⅲ)+C2H2组的氮气产量低于NH4++Fe(Ⅲ)组(P<0.05), NH4++C2H2组的氮气产量低于NH4+组(P < 0.01), 这主要是由于乙炔的加入对反硝化及ANAMMOX微生物群落具有抑制作用[11, 13, 19].因此, NH4++C2H2组和NH4++Fe(Ⅲ)+C2H2组体系产生的氮气是由于其他微生物的作用, 可能的途径是Fe(Ⅲ)参与的Feammox或SO42-参与的S-ANAMMOX.
本实验各体系中硫酸盐和硫化物初始含量差异不显著, 分别是(307.59±65.21)mg·kg-1和(64.64±3.83) mg·kg-1.反应体系培养15 d后, 硫酸盐含量为(284.11±25.48)~(295.1±38.73) mg·kg-1, 硫化物含量为(89.04±12.51)~(114.56±25.93) mg·kg-1, 单质硫在体系中并没有被检测到[图 1(b)].富铁沉积物中, Fe(Ⅲ)与SO42-同时作为电子受体具有竞争关系, Fe(Ⅲ)抑制了SO42-的还原[20].因此, 在凹山湿地沉积物中硫酸盐的还原过程主要由硫酸盐还原菌参与作用, 没有发生S-ANAMMOX生物化学过程. NH4++C2H2组和NH4++Fe(Ⅲ)+C2H2组体系由于C2H2的抑制作用, N2主要为Feammox过程产生的, 如反应方程式(1)所示.此外, 本实验中含C2H2实验组中产氮气速率减少了0.19 mg·(kg·d)-1, 同Ding等[8]的报道数据接近. Yang等[10]的研究发现在培养24 h热带森林土壤后Feammox产生氮气速率大约为0.32 mg·(kg·d)-1, Li等[12]的研究发现在培养24 h潮汐带湿地沉积物中Feammox产生氮气速率大约为0.24~0.36 mg·(kg·d)-1, 且潮汐对Feammox有着一定的影响, 大潮时期Feammox活性更强.

|
(1) |
在NH4+组和NH4++C2H2组中, Fe(Ⅲ)还原速率提高, Fe(Ⅱ)生成量高于空白对照组(图 2), 说明氨氮的加入能够促进Fe(Ⅲ)还原[13].此外, 在NH4++Fe(Ⅲ)组和NH4++Fe(Ⅲ)+C2H2组体系中, Fe(Ⅱ)生成量分别为(3 394.10±286.34) mg·kg-1和(2 730.81±442.11) mg·kg-1, Fe(Ⅲ)还原速率显著提高.在NH4+组和NH4++C2H2组体系中Fe(Ⅱ)生成量分别为(1 139.97±123.93) mg·kg-1和(1 043.50±54.69) mg·kg-1, 加入外源铁体系的Fe(Ⅱ)生成量明显高于其他没有加入外源铁体系组(图 2). NH4++Fe(Ⅲ)组比NH4+组高出197.36%, NH4++Fe(Ⅲ)+C2H2组比NH4++C2H2组高出161.70%, 说明水铁矿能够被微生物利用进而促进了Feammox生物化学过程[10, 12, 21].本实验中NH4++C2H2组和NH4++Fe(Ⅲ)+C2H2组体系中Feammox产生氮气量为(25.35±3.32) mg·kg-1和(38.18±1.23) mg·kg-1, 说明铁氧化物的生物可利用性和还原速率能够对Feammox起到明显的调控作用[9], 氨氮氧化速率从1.69 mg·(kg·d)-1增强到2.72 mg·(kg·d)-1.文献报道Feammox种群对土壤中生物可利用铁氧化物含量有很大的依赖性[9~12]. Fe(Ⅲ)还原程度和速率与比表面积有着显著的关系[22, 23]. Fe(Ⅲ)可利用性很高, 能够促进Feammox生化过程同时还原生成Fe(Ⅱ)[9, 11, 13].

|
图 2 培养15 d后反应体系中盐酸可提取态Fe(Ⅱ)含量 Fig. 2 HCl-extractable Fe(Ⅱ) content in the reaction system after 15 days |
如图 1所示, NH4+组体系的N2量明显高于NH4++Fe(Ⅲ)组的反应体系(P < 0.01), 表明水铁矿的加入影响沉积物生物化学过程, 进而显著降低系统的氨氮流失量.以Fe(Ⅲ)为电子受体的厌氧自养型微生物群落与以NO2-为电子受体的ANAMMOX菌群在电子传递上存在竞争关系.[24] Fe(Ⅱ)与反硝化产生的NO2-通过表面催化作用将Fe(Ⅱ)氧化成Fe(Ⅲ), 矿化成针铁矿[25], 使其附着在细菌表面, 阻止了基质NO2-和NO3-进入细胞内, 即形成钝化, 进而抑制了ANAMMOX和反硝化过程[22, 24, 26, 27].由于实验中NH4++Fe(Ⅲ)组和NH4++Fe(Ⅲ)+C2H2组中以NO2-为电子受体的ANAMMOX过程可能受到铁还原过程抑制. NH4++Fe(Ⅲ)组的氮气主要产生于反硝化和Feammox过程, 其中反硝化速率较低, 约为0.19 μg·(kg·d)-1, 这主要是由于水铁矿的加入抑制了反硝化微生物与繁殖[26].
2.3 一氧化二氮气(N2O)和硝酸盐的产生土壤生态系统中NO2-和NO3-的主要产生途径是有机质矿化作用和硝化作用, 消耗途径是反硝化过程及厌氧氨氧化过程[8, 28].研究发现厌氧环境下有机大分子矿化作用产生硝酸盐的效率远低于好氧环境, 通常小于1 μmol·L-1[29].本实验条件为厌氧环境, 有机质矿化过程效率较低, 产生的硝酸盐含量可以忽略不计.因此, 沉积物中NO3-的含量改变主要受到反硝化过程的影响.
在空白组和其他实验组, 在任何时间段都没有检测到NO2-.加入NH4Cl溶液的4组体系在培养15 d后, 均检测到了NO3-的产出, 含量约为4.43 mg·kg-1, 平均产生速率约为0.288 μg·(kg·d)-1, 而空白对照组没有检测到. Yang等[10]的研究发现在热带森林土壤中Feammox现象, 并且可以产生N2、NO2-或者NO3-[方程式(1)~(3)].本实验可以发生Feammox生物代谢过程, 以及检测到了NO3-的产出, 说明NH4Cl溶液的加入影响了NO3-的产出, 反应体系中的NO3-主要来源于Feammox过程, 即反应方程式(3).
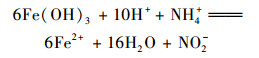
|
(2) |
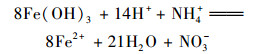
|
(3) |
如图 3所示, NH4+、NH4++C2H2、NH4++Fe(Ⅲ)和NH4++Fe(Ⅲ)+C2H2组体系中均产生了N2O, 其产量从(16.22±1.07)~(31.46±1.67) mg·kg-1, 空白对照组没有检测到N2O的产生.氨氮的加入提高了微生物群落活性, 进而促进了N2O的产出.如前所述, C2H2的加入抑制反硝化过程, 从而导致N2O的累积.另外, NH4++Fe(Ⅲ)+C2H2组体系中N2O产量显著高于NH4++C2H2组体系(P < 0.01), 这表明水铁矿的加入显著促进了N2O的产生.王庆等[26]的研究发现, 水稻土在添加外源铁对N2O的产生和排放产生了显著的影响, 高添加量水铁矿处理导致硝态氮含量明显高于对照和N2O排放速率先低后高于对照, 外源铁抑制了反硝化功能微生物种群数量. Feammox生物代谢过程可能产生N2O[12], 此外在实验中也发现了向土壤添加水铁矿能够促进N2O的产生[27, 30].卢燕宇等[31]经过多年建立农田N2O年度排放数据库, 分析发现N2O排放与温度及降水呈现显著相关性, 与土壤pH值、有机碳及氮含量无显著相关性.因此, 将卢燕宇等[31]利用统计学和经温度因子修正后的N2O排放估算方法运用到本实验数据中, 本实验条件下得到N2O的理论排放量为5.74 mg·kg-1, 而本实验N2O的实际排放量远高于此数值.另外, 本实验反硝化产氮气速率约为0.19 μg·(kg·d)-1, 说明了N2O的产生可能存在新途径, 来自Feammox过程[反应方程式(4)].
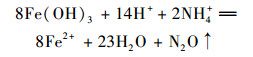
|
(4) |

|
图 3 培养15 d后反应体系中N2O产量 Fig. 3 Production of N2O in the reaction system after 15 days |
在好氧环境中, 硝化过程主导氮的转化, 先将氨氮氧化成NO2-再氧化成NO3-[32].在厌氧环境下, NH4+的浓度是随着有机氮矿化程度增加而得到积累[33]以及NOx-被ANAMMOX和反硝化还原[9].但是, 本实验中NH4+的浓度在4个厌氧体系中均逐渐减少, 而NOx-存在积累现象. 4组体系中均发生了Fe(Ⅲ)的还原, Fe(Ⅱ)随着水铁矿的加入而显著升高(图 2).同时, ANAMMOX及反硝化过程受水铁矿抑制作用, 氨氮的转化率仅仅降低了20%.这表明Fe(Ⅲ)在处理采矿废水湿地沉积物中作为电子受体, 改变了氨氮转化途径, 发生了氮循环新途径——Feammox过程[8~13].
培养15 d后NH4+和NH4++Fe(Ⅲ)组体系中氨氮剩余量从20%增加到40%, 其他途径也有所改变(图 4).其中, ANAMMOX和反硝化过程在NH4+组体系中消耗氨氮24.99 mg·kg-1和12.83 mg·kg-1, 占比28%和14%, 而在NH4++Fe(Ⅲ)组体系中ANAMMOX过程完全消失, 反硝化过程也降低到了2.88 mg·kg-1, 占比3%.同时, NH4+组和NH4++Fe(Ⅲ)组体系沉积物氮损失分别达到了76%和51%, 说明水铁矿的加入在一定程度上减少了处理采矿废水湿地沉积物氮损失, 这与水铁矿二次成矿抑制了ANAMMOX和反硝化过程有着显著的关系[22, 25]. Feammox过程在NH4+组体系中消耗氨氮25.35 mg·kg-1, 占比28%, 而在NH4++Fe(Ⅲ)组体系中Feammox过程增加到了38.18 mg·kg-1, 占比42%, 说明水铁矿促进了Feammox过程, 其中氧化氨氮速率从1.68 mg·(kg·d)-1增强到2.72 mg·(kg·d)-1, 对氮损失贡献率从40%增加到了82%.因此, 在凹山湿地沉积物氨氮的厌氧氧化过程中, NH4+被转化成N2、N2O或者NO3-进一步进行脱氮, 而水铁矿的加入改变了氨氮的转化途径和减少了氮损失, 这也为贫瘠湿地的水生生物和植物保留氮源提供一定的参考价值.

|
内环:NH4+, 外环: NH4++Fe(Ⅲ) 图 4 培养15 d后NH4+和NH4++Fe(Ⅲ)组体系中氮平衡和流失途径分析 Fig. 4 Nitrogen mass balance and loss paths in the NH4+ and NH4++Fe(Ⅲ)systems after 15 days |
本实验证明氮气直接产生于Feammox过程, 乙炔抑制剂的添加直接将其与通过反硝化和ANAMMOX产生的氮气所区分.一些证据说明Feammox可能直接产生NO3-及N2O.但是, Feammox直接产生NO3-和N2O的速率需要进一步研究, 这可能对全球变暖气体的产生途径有所影响.湿地中氨氮的转化是以Feammox产生氮气为主要过程, 而不是将还原价态氨氮氧化成高价态的亚硝酸盐或硝酸盐, 再被还原成氮气.因此, Feammox过程对氮损失有着不可忽视的重要性, 在特定环境中比反硝化和ANAMMOX的影响更为显著.比如, 富铁水稻土过量施肥可能会影响湿地植物和水稻的生长[8].
3 结论(1) 培养期间发现了Feammox过程直接产生了氮气, 以及可能直接产生NO3-及N2O.
(2) 乙炔抑制剂很大程度上使ANAMMOX过程得到抑制, 而对Feammox过程没有显著影响, 两个过程产氮量区分效果明显, ANAMMOX产生氮气量为24.99 mg·kg-1, Feammox产生氮气量为25.35 mg·kg-1, 为接下来通过乙炔抑制剂技术研究Feammox过程提供较为可靠的依据.
(3) 水铁矿通过二次成矿抑制了ANAMMOX和反硝化过程, 以及通过促进Feammox过程氧化氨氮速率而改变了氨氮的转化途径和减少了氮损失.但是, 水铁矿在一定程度上抑制了人工湿地去除硫酸盐效率.因此在后续研究中为提高人工湿地去除硫酸盐效率, 需更深一步探讨水铁矿的添加量.
| [1] |
周念清, 王燕, 钱家忠, 等. 湿地氮循环及其对环境变化影响研究进展[J]. 同济大学学报(自然科学版), 2010, 38(6): 865-869. Zhou N Q, Wang Y, Qian J Z, et al. Advances in research on nitrogen cycle in wetlands and its influence on environmental change[J]. Journal of Tongji University (Natural Science), 2010, 38(6): 865-869. |
| [2] | Dørge J. Modelling nitrogen transformations in freshwater wetlands[J]. Ecological Modelling, 1994, 75-76: 409-420. DOI:10.1016/0304-3800(94)90036-1 |
| [3] | Mitsch W J, Gosselink J G. Wetlands (5th ed.)[J]. New York:NJ John Wiley and Sons Inc., 2015, 259-420. |
| [4] |
任伊滨, 任南琪, 李志强. 冻融对小兴安岭湿地土壤微生物碳、氮和氮转换的影响[J]. 哈尔滨工程大学学报, 2013, 34(4): 530-535. Ren Y B, Ren N Q, Li Z Q. Effects of freeze-thaw cycles on soil microbial carbon, nitrogen and nitrogen conversion in Xiaoxing'anling boreal wetland of China[J]. Journal of Harbin Engineering University, 2013, 34(4): 530-535. |
| [5] | Yang X R, Li H, Nie S A, et al. Potential contribution of anammox to nitrogen loss from paddy soils in southern China[J]. Applied and Environmental Microbiology, 2015, 81(3): 938-947. DOI:10.1128/AEM.02664-14 |
| [6] | Deng F Y, Hou L J, Liu M, et al. Dissimilatory nitrate reduction processes and associated contribution to nitrogen removal in sediments of the Yangtze Estuary[J]. Journal of Geophysical Research:Biogeosciences, 2015, 120(8): 1521-1531. DOI:10.1002/2015JG003007 |
| [7] | Shen L D, Wu H S, Gao Z Q, et al. Distribution and activity of anaerobic ammonium-oxidising bacteria in natural freshwater wetland soils[J]. Applied Microbiology and Biotechnology, 2016, 100(7): 3291-3300. DOI:10.1007/s00253-015-7191-z |
| [8] | Ding L J, An X L, Li S, et al. Nitrogen loss through anaerobic ammonium oxidation coupled to iron reduction from paddy soils in a chronosequence[J]. Environmental Science & Technology, 2014, 48(18): 10641-10647. |
| [9] | Clément J C, Shrestha J, Ehrenfeld J G, et al. Ammonium oxidation coupled to dissimilatory reduction of iron under anaerobic conditions in wetland soils[J]. Soil Biology and Biochemistry, 2005, 37(12): 2323-2328. DOI:10.1016/j.soilbio.2005.03.027 |
| [10] | Yang W H, Weber K A, Silver W L. Nitrogen loss from soil through anaerobic ammonium oxidation coupled to iron reduction[J]. Nature Geoscience, 2012, 5(8): 538-541. DOI:10.1038/ngeo1530 |
| [11] | Shrestha J, Rich J J, Ehrenfeld J G, et al. Oxidation of ammonium to nitrite under iron-reducing conditions in wetland soils:Laboratory, field demonstrations, and push-pull rate determination[J]. Soil Science, 2009, 174(3): 156-164. DOI:10.1097/SS.0b013e3181988fbf |
| [12] | Li X F, Hou L J, Liu M, et al. Evidence of nitrogen loss from anaerobic ammonium oxidation coupled with ferric iron reduction in an intertidal wetland[J]. Environmental Science & Technology, 2015, 49(19): 11560-11568. |
| [13] | Sawayama S. Possibility of anoxic ferric ammonium oxidation[J]. Journal of Bioscience and Bioengineering, 2006, 101(1): 70-72. DOI:10.1263/jbb.101.70 |
| [14] |
陶巍. 某矿区酸性矿山废水污染土壤及湿地处理系统的调查研究[D]. 合肥: 合肥工业大学, 2017. 44-55. Tao W. The investigation on soil contaminated by acid mine wastewater and a wetland treatment system in a mining area[D]. Hefei: Hefei University of Technology, 2017. 44-55. http://cdmd.cnki.com.cn/Article/CDMD-10359-1017058462.htm |
| [15] |
李石磊. 气相分子吸收光谱法检测气液固三相硫化物的研究[D]. 合肥: 合肥工业大学, 2014. 12-29. Li S L. Determination of sulfide sulfur in gas-liquid-solid phase by gas-phase molecular absorption spectrometry[D]. Hefei: Hefei University of Technology, 2014. 12-29. http://cdmd.cnki.com.cn/Article/CDMD-10359-1015666408.htm |
| [16] |
孙志高, 刘景双, 杨继松, 等. 三江平原典型小叶章湿地土壤硝化-反硝化作用与氧化亚氮排放[J]. 应用生态学报, 2007, 18(1): 185-192. Sun Z G, Liu J S, Yang J S, et al. Nitrification-denitrification and N2O emission of typical Calamagrostis angustifolia wetland soils in Sanjiang plain[J]. Chinese Journal of Applied Ecology, 2007, 18(1): 185-192. |
| [17] | Barnes R T, Smith R L, Aiken G R. Linkages between denitrification and dissolved organic matter quality, Boulder Creek watershed, Colorado[J]. Journal of Geophysical Research:Biogeosciences, 2015, 117(G1): G01014. |
| [18] | Yoshinari T, Hynes R, Knowles R. Acetylene inhibition of nitrous oxide reduction and measurement of denitrification and nitrogen fixation in soil[J]. Soil Biology and Biochemistry, 1977, 9(3): 177-183. DOI:10.1016/0038-0717(77)90072-4 |
| [19] | Jensen M M, Thamdrup B, Dalsgaard T. Effects of specific inhibitors on anammox and denitrification in marine sediments[J]. Applied and Environmental Microbiology, 2007, 73(10): 3151-3158. DOI:10.1128/AEM.01898-06 |
| [20] | Lovley D R, Phillips E J P. Competitive mechanisms for inhibition of sulfate reduction and methane production in the zone of ferric iron reduction in sediments[J]. Applied and Environmental Microbiology, 1987, 53(11): 2636-2641. |
| [21] |
程冰如. 针铁矿促进硝酸盐生物脱氮的初步研究[D]. 安徽: 合肥工业大学, 2015. 14-29. Cheng B R. Preliminary study on enhanced biological nitrate reduction with goethite[D]. Hefei: Hefei University of Technology, 2015. 14-29. http://cdmd.cnki.com.cn/Article/CDMD-10359-1015666379.htm |
| [22] | Schädler S, Burkhardt C, Hegler F, et al. Formation of cell-iron-mineral aggregates by phototrophic and nitrate-reducing anaerobic Fe(Ⅱ)-oxidizing bacteria[J]. Geomicrobiology Journal, 2009, 26(2): 93-103. DOI:10.1080/01490450802660573 |
| [23] |
谢晶晶, 庆承松, 陈天虎, 等. 几种铁(氢)氧化物对溶液中磷的吸附作用对比研究[J]. 岩石矿物学杂志, 2007, 26(6): 535-538. Xie J J, Qing C S, Chen T H, et al. A comparison of phosphate adsorption capacity between different iron hydroxides/oxides[J]. Acta Petrologica et Mineralogica, 2007, 26(6): 535-538. |
| [24] | Coby A J, Picardal F W. Inhibition of NO3- and NO2- reduction by microbial Fe(Ⅲ) reduction:evidence of a reaction between NO2- and cell surface-bound Fe2+[J]. Applied and Environmental Microbiology, 2005, 71(9): 5267-5274. DOI:10.1128/AEM.71.9.5267-5274.2005 |
| [25] |
刘承帅, 李芳柏, 陈曼佳, 等. Fe(Ⅱ)催化水铁矿晶相转变过程中Pb的吸附与固定[J]. 化学学报, 2017, 75(6): 621-628. Liu C S, Li F B, Chen M J, et al. Adsorption and stabilization of lead during Fe(Ⅱ)-catalyzed phase transformation of ferrihydrite[J]. Acta Chimica Sinica, 2017, 75(6): 621-628. |
| [26] |
王庆, 杨会翠, 王玲, 等. 添加水铁矿对水稻土N2O释放及反硝化微生物的影响[J]. 土壤学报, 2016, 53(5): 1306-1315. Wang Q, Yang H C, Wang L, et al. Effect of amending ferrihydrite on N2O emission and denitrifying microorganisms in paddy soil[J]. Acta Pedologica Sinica, 2016, 53(5): 1306-1315. |
| [27] | Zhu X, Silva L C R, Doane T A, et al. Iron:the forgotten driver of nitrous oxide production in agricultural soil[J]. PLoS One, 2013, 8(3): e60146. DOI:10.1371/journal.pone.0060146 |
| [28] | Burns L C, Stevens R J, Laughlin R J. Determination of the simultaneous production and consumption of soil nitrite using15N[J]. Soil Biology and Biochemistry, 1995, 27(6): 839-844. DOI:10.1016/0038-0717(94)00219-Q |
| [29] | Hansen L S, Blackburn T H. Aerobic and anaerobic mineralization of organic material in marine sediment microcosms[J]. Marine Ecology Progress Series, 1991, 75: 283-291. DOI:10.3354/meps075283 |
| [30] | Huang B, Yu K W, Gambrell R P. Effects of ferric iron reduction and regeneration on nitrous oxide and methane emissions in a rice soil[J]. Chemosphere, 2009, 74(4): 481-486. DOI:10.1016/j.chemosphere.2008.10.015 |
| [31] |
卢燕宇, 黄耀, 郑循华. 农田氧化亚氮排放系数的研究[J]. 应用生态学报, 2005, 16(7): 1299-1302. Lu Y Y, Huang Y, Zheng X H. N2O emission factor for agricultural soils[J]. Chinese Journal of Applied Ecology, 2005, 16(7): 1299-1302. |
| [32] | Caffrey J M, Bano N, Kalanetra K, et al. Ammonia oxidation and ammonia-oxidizing bacteria and archaea from estuaries with differing histories of hypoxia[J]. The ISME Journal, 2007, 1(7): 660-662. DOI:10.1038/ismej.2007.79 |
| [33] | Santos I R, Cook P L M, Rogers L, et al. The "salt wedge pump":convection-driven pore-water exchange as a source of dissolved organic and inorganic carbon and nitrogen to an estuary[J]. Limnology and Oceanography, 2012, 57(5): 1415-1426. DOI:10.4319/lo.2012.57.5.1415 |