 2018, Vol. 39
2018, Vol. 39



2. 中国科学院地球化学研究所, 贵州 550002
2. Institute of Geochemistry, Chinese Academy of Sciences, Guizhou 550002, China
邻苯二甲酸酯类(phthalate esters, PAEs)在日常及工业上被广泛使用, 其中邻苯二甲酸二(2-乙基己基)酯(DEHP)的用量最大, 其次是邻苯二甲酸二丁酯(DBP). DBP作为环境激素的一种, 对生物生存、生态平衡以及人体健康有着巨大的威胁.有研究表明其对水体中藻类(短凯伦藻、短裸甲藻、杜氏盐藻、角毛藻等)有较强的毒性[1~3].
氧化石墨烯(graphene oxide, GO)因其独特的机械、电学、光学及热性能等性质被广泛用于各个领域.一些研究表明, GO进入环境接触生物时, 在基因、细胞、器官、个体尺度上会表现出一定的负面影响, 毒性机制受尺寸、受试生物种类、部位等因素的影响. GO相关纳米材料在一定条件下有一定的细胞毒性, 能改变基因表达, 介导DNA损伤, 影响细胞周期和代谢[4, 5], 导致氧化应激[5~8]、细胞凋亡[5, 7]、免疫毒性[8]、抗菌性[9, 10]、藻抑制性[11, 12]、遗传毒性[13]等.也有一些研究表明, GO对一些生物没有表现出毒性效应或毒性很小.一定浓度的GO对斑马鱼个体没有明显的急性毒性[8], 对人红细胞和皮肤成纤维细胞[14]、Hela细胞[15]、A549细胞表现较低的毒性作用[16].环境中GO与污染物的接触过程复杂, 对生物体的共同作用不能轻视.研究显示, HA的存在抑制了GO或rGO对PAEs的吸附, 增强解吸附作用, 意味着GO能通过吸附环境中的污染物释放到生物体内[17]; GO减少了PCB-52对human-hamster hybrid AL细胞的生长毒性和基因毒性等[18]; 在GO与Cu2+的联合毒性实验中, 因GO的吸附作用减少大型溞体内Cu2+的富集量, 从而减缓Cu2+对大型溞的毒性[19].
新型纳米材料类污染物进入海洋环境后, 与原本存在的传统有机污染物共同作用于海洋生物, 对海洋生态造成潜在危害. GO与污染物对水生生物的联合毒性效应现象多样, 机制尚不明确.青岛大扁藻系单细胞海洋微藻, 海洋中分布广泛, 易被海产珍品幼体摄食, 在海洋生态环境评价中具有代表性和易操作性.本实验以青岛大扁藻为研究对象, 结合藻细胞生长状况、光能利用、细胞通透性、以及氧化胁迫指标, 分析低浓度GO存在时对DBP藻毒性的影响, 以揭示GO存在对海洋生物的风险.
1 材料与方法 1.1 实验材料青岛大扁藻(Platymonas helgolanidica var. tingtaoensis)购自中科院海洋所藻种库.藻种培养:采用f/2培养基于人工智能光照培养箱(GXZ-500Z江南仪器公司, 宁波)中培养.培养条件为:光暗比为14 h:10 h, 昼夜温度比为24℃:22℃, 白天光照强度为6 000 lx, 每天摇晃藻液2次, 正式实验前藻液预培养至对数生长期.纳米材料:高纯度试剂级氧化石墨烯粉末(JCGO-99-1-2)购自南京吉仓纳米科技有限公司.
1.2 实验方法 1.2.1 氧化石墨烯分散液分析天平称取0.005 g氧化石墨烯粉末于100 mL容量瓶, 用灭菌海水配置的培养液定容, 振荡混匀, 配置50 mg·L-1的GO母液.将母液冰水浴超声40 min后, 慢速磁力搅拌2 h, 放置4℃冰箱黑暗条件保存待用.使用前需再超声分散30~40 min, 以保证充分分散, 并置于紫外灯下灭菌30 min.
1.2.2 各毒性指标的测定(1) 藻密度的测定用血球计数板在普通光学显微镜下计数, 每个实验样品计数3次取平均值.
(2) 叶绿素a的测定取4 mL藻液于5 mL的离心管中, 离心(7500g, 10 min, 10℃), 去上清液, 重复1~3次.然后向藻泥中加入4 mL 95%乙醇, 置于黑暗条件下提取18~24 h, 测量665 nm和649 nm波长下吸光度值(D665和D649), 用公式计算叶绿素a、叶绿素b的浓度(CChla和CChlb), 叶绿素a与叶绿素b的加和作为总叶绿素含量, 单位:mg·L-1.
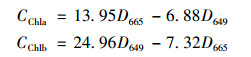
|
(3) 细胞通透性的测量方法乙二酸荧光素(FDA)是一种非极性非荧光的酯化有机物, 可以穿过细胞膜, 自由进入细胞.通过FDA可以在活细胞中酯酶的作用下分解释放出荧光素的特性, 可以利用其荧光强度间接反映细胞膜的通透性[20].具体实验操作:取4 mL藻液于10 mL的离心管中, 离心(7500g, 10min, 15℃), 去上清液, 重复1~3次得到藻泥, 向藻泥中加入FDA 0.7 mL, 黑暗条件下培养20 min, 每隔3~5min摇动一次.充分反应后用磷酸盐缓冲溶液(PBS, pH=7.0)清洗多余FDA, 重复1~3次, 定容到10 mL, 用分子荧光光度计(激发波长485 nm, 发射波长521 nm)测荧光强度.
(4) SOD、ROS的测定方法取4 mL藻液于5 mL离心管中, 离心(7500g, 5 min, 4℃), 去上清液, 重复1~3次得到藻泥, 向藻泥中入加PBS至1 mL, 在超声波细胞破碎仪中冰水浴超声破碎(5 min, 间隔30 s), 得到粗酶液.然后严格按照SOD、ROS试剂盒(南京建成)说明书进行操作, 测得SOD、ROS值.
1.2.3 GO暴露对青岛大扁藻的影响将青岛大扁藻藻液按每瓶100 mL分装到200 mL锥形瓶中.置于光照培养箱中适应24 h, 后进行暴露实验. GO浓度设置:0、0.1、0.2、0.5、1.0、2.0、5.0、10.0 mg·L-1, 每组浓度设3个平行, 然后分别在0、24、48、72、96 h测定藻液的藻密度、叶绿素含量和细胞通透性.
1.2.4 GO对DBP藻毒性的影响将青岛大扁藻藻液按每瓶100 mL分装到200 mL锥形瓶中.置于光照培养箱中适应24 h, 后开始暴露实验. DBP浓度设置:0.1、0.5、1.0、2.0、4.0 mg·L-1, GO根据单独暴露实验结果选择1 mg·L-1作为联合暴露实验浓度, 每个浓度设定DBP单独组和GO添加组, 分别表示为D0.1、D0.5、D1.0、D2.0、D4.0和GD0.1、GD0.5、GD1.0、GD2.0、GD4.0, 每组设3个平行.然后分别在0、24、48、72、96 h测定藻液的藻密度、叶绿素含量、细胞通透性、ROS和SOD.
1.2.5 青岛大扁藻的扫描电镜(SEM)将污染物暴露96 h后的青岛大扁藻样品, 取10 mL离心分离(9000g, 10 min, 4℃), 抽滤, 经冷冻干燥后, 用扫描电镜检测(JSM-6390LV, JEOL, Japan).
1.3 分析方法青岛大扁藻生长抑制率IR按公式计算,
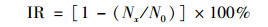
|
式中, Nx是观测时实验组的藻密度, cell·L-1; N0为观测时对照组的藻密度, cell·L-1.利用Origin 9.1和SPSS 19.0软件进行数据分析和方差分析(ANOVA).
2 结果与讨论 2.1 GO的表征其TEM和FTIR表征如图 1.从氧化石墨烯的TEM结果可以看出, 本实验所用的GO是一类只有一个维度在纳米级的纳米材料, 其大小尺寸为1~5 μm, 厚度为0.8~1.2 nm.氧化石墨烯的红外曲线出现了大量含氧官能团的特征峰, 如1717 cm-1处出现羧基(C=O)的特征峰, 3 404 cm-1处特征峰代表着羟基(—OH)的伸缩振动, 1 587 cm-1处特征峰代表着羟基的弯曲振动, 表明本实验所用氧化石墨烯中氧化基团主要是羟基和水分子.

|
(a)GO分散液的透射电镜图,(b)氧化石墨烯粉末的FTIR图 图 1 GO材料的表征 Fig. 1 Characterization of GO |
图 2中显示了青岛大扁藻在0.1~10 mg·L-1GO的单独作用下的0~96 h藻密度变化曲线以及96 h EC50的拟合评估. GO对青岛大扁藻的生长在72 h前表现出略微的刺激作用, 藻密度高于空白组, 整个暴露过程对青岛大扁藻生长没有表现出明显的抑制作用. 96 h剂量-抑制率图显示0.1~10 mg·L-1GO对青岛大扁藻的藻密度的抑制率均低于10%, EC50, 96 h > 100 mg·L-1(R2=0.965, P < 0.01), 说明GO对青岛大扁藻毒性很小.之前的研究表明, 石墨烯对斜生栅藻(Scenedesmus obliquus)的72 h IC50为150 mg·L-1左右, 其所用的石墨烯与本实验使用的GO尺寸相近, 结果也相近, 10 mg·L-1时对藻的生长抑制低于10%[21]. GO对羊角月牙藻(Raphidocelis subcapitata)的96 h EC50为20 mg·L-1, 其平均尺寸大小(粒径120~200 nm, 厚3.5 nm)比本实验所用的GO(粒径1~5 μm, 厚0.8~1.2 nm)小得多, 表现出的毒性比青岛大扁藻大[22].与Tang等[23]的研究结果相近, 其研究表明10 mg·L-1以下GO(粒径588 nm)对铜绿微囊藻(Microcystis aeruginosa)的生长没有抑制作用, 但两者作用机制是否相同仍需进一步研究.本实验表明在96 h内没有观察到明显的生长抑制, Hu等[12]的研究显示GO(粒径1~5 μm, 厚0.8~1 nm)对小球藻(Chlorella vulgaris)的生长抑制出现在144 h.

|
(a) 0~96 h各组藻细胞生长情况;(b)GO暴露96 h的抑制率-浓度曲线;红线为拟合曲线,灰色部分为95%置信区间 图 2 GO对青岛大扁藻的生长抑制作用 Fig. 2 Effect of GO on the cell density and growth inhibition in algae |
光合色素是客观反映藻类利用光照能力的一类重要物质, 藻类的光合色素会随着环境的改变和生长发育阶段的不同而发生变化, 其含量的高低可以表征藻类的存活情况和生物量的大小. 图 3显示了0~96 h青岛大扁藻在GO暴露组和空白组叶绿素a、叶绿素b、总叶绿素的含量变化情况, 以及96 h不同浓度GO暴露组相对空白组的叶绿素a含量水平. 图 3(a)~3(c)显示了叶绿素a、叶绿素b、总叶绿素含量与藻的生长状况一致, 整个暴露过程, 叶绿素a、叶绿素b、总叶绿素的含量相比空白组没有明显差异. 图 3(d)可以看到96 h GO的暴露对叶绿素a的含量有略微的抑制, 抑制率在10%以内, 但是相对于空白组没有显著的差异.与Tang等[23]的研究结果相似, 其结果表明GO浓度在低于10 mg·L-1时对铜绿微囊藻的光合作用参数(电子产率, 叶绿素a产量, 光合作用效率, 最大电子转移量)没有显著性影响.向昆仑等[24]研究了高毒性污染物双酚A对青岛大扁藻的影响, 发现双酚A能明显抑制青岛大扁藻的光合色素产量.这说明0.1~10 mg·L-1GO对青岛大扁藻的光合作用没有明显影响, 也反映了GO对青岛大扁藻的低毒性.

|
图 3 GO对青岛大扁藻光合色素的影响 Fig. 3 Effect of GO on the photosynthetic pigments of algae |
GO作为一种2D材料, 多项研究发现其独特的片层结构可能对细胞膜造成物理性的割伤、刺破等损害, 导致细胞通透性改变.实验中的细胞通透性在一定程度上可以反映细胞膜的完整性, 为消除细胞密度变化的影响, 以FDA法实测荧光强度与藻细胞数的比值作为单个细胞通透性水平. 图 4显示了0~96 h在GO暴露的情况下单个藻细胞通透性水平(初始空白组为1)的变化情况以及96 h单个藻细胞通透性水平(空白组为1)随GO浓度的变化. 图 4(a)中GO暴露组藻细胞通透性水平随时间呈现升高的趋势, 空白组变化不明显, 5 mg·L-1和10 mg·L-1 GO暴露组藻细胞通透性一直保持较高水平, 最高时分别达到初始空白组的2.6和3.2倍, 说明GO的暴露造成青岛大扁藻细胞通透性改变, 且时间越长, 影响越大. 图 4(b)中显示了96 h不同浓度GO暴露单个藻细胞通透性水平随浓度的增加显著升高(P < 0.05), 0.1~10 mg·L-1分别增加到空白组的1.2~2.2倍. GO(厚度约为0.8~1 nm)的锋利边缘结构在与细胞的相互接触过程中, 会直接对细胞产生一定的物理性损伤(划破细胞壁, 刺穿细胞膜等), 影响细胞通透性[9]. GO浓度越高, 在培养体系中与藻细胞接触的概率越大, 培养过程中摇晃混匀也可能对此造成影响. Ouyang等[20]的研究也发现1和10 mg·L-1的片状GO(粒径1~5 μm, 厚0.8~1 nm)和0.01~10 mg·L-1的量子点GO (粒径20~50 nm, 厚4.8~5.2 nm)暴露后显著改变了小球藻细胞通透性.细胞表面物理损伤是导致藻密度和叶绿素产量略微抑制的主要原因, 但低浓度GO引起的物理性损伤对藻种群的生长没有明显的影响.

|
(a)单个藻细胞细胞通透性水平相对于空白组0~96 h变化情况; (b)96 h时不同浓度GO组藻细胞通透性水平比较; 不同小写字母表示各组间的差异, P < 0.05 图 4 GO对青岛大扁藻细胞通透性的影响 Fig. 4 Effect of GO on the cell permeability in algae cell |
GO单独暴露的实验表明0.1~10 mg·L-1 GO对青岛大扁藻的生长无明显影响, 本实验选择1 mg·L-1的GO与不同浓度的DBP联合暴露, 以观察低浓度GO的加入对DBP藻毒性的影响.如图 5所示, 显示了DBP单独及与1 mg·L-1 GO同时存在时, 青岛大扁藻0~96 h藻密度变化情况以及DBP浓度变化对藻生长抑制作用的影响. 图 5(a)可以看出DBP单独及与1 mg·L-1 GO同时存在时, 相比于空白组, 青岛大扁藻在0~96 h藻密度受到明显抑制, DBP对青岛大扁藻表现出较高的毒性作用. 图 5(b)中拟合了96 h DBP的浓度-抑制率曲线, 得到DBP对青岛大扁藻的EC50为(11.14±0.80)mg·L-1.不同的藻类对DBP的耐受程度不同, 文献中显示DBP对淡水藻斜生栅藻和小球藻的96h EC50分别为15.3 mg·L-1和3.14 mg·L-1[25].对比96 h DBP单独及与1 mg·L-1GO同时存在的抑制率[图 5(c)], 发现在低浓度DBP(0.2~2 mg·L-1)组, 1 mg·L-1 GO的加入对青岛大扁藻的生长抑制作用没有明显的差异, 在高浓度DBP(4 mg·L-1)组, 1 mg·L-1 GO的加入对藻的96 h生长抑制作用有显著的增强作用(P < 0.05), 抑制率从34%增加到45%. 1 mg·L-1 GO的加入使DBP的96h EC50从(11.14±0.80)mg·L-1下降到(4.93±2.14)mg·L-1, Tang等[23]在研究GO和Cd联合下对铜绿微囊藻的毒性效应时, 发现低浓度(1 mg·L-1)GO作用下没有表现出毒性效应, 但是GO的存在加剧了Cd的毒性. Yu等[26]的研究表明, GO的细胞毒性除了传统的物理损伤和氧化应激等, 还可能是由于GO中的含氧官能团与铁结合导致含铁代谢受阻而引起抑制, 可以通过测量培养体系中的Fe含量研究此过程.

|
(a)为0~96 h各组藻密度的变化情况; (b)、(c)分别为96 h DBP与GO+DBP组藻密度的抑制率-浓度曲线(红线为拟合线, 灰色部分为95%置信区间); (d)为96 h DBP与GO+DBP组藻抑制率对比(空白组为1), *表示显著差异, P < 0.05, 下同 图 5 有无GO添加时DBP对青岛大扁藻密度的影响 Fig. 5 Effect of DBP, with and without the addition of GO, on algae density |
图 6中显示了计算得到的DBP单独及与1 mg·L-1 GO同时存在时, 培养体系中藻细胞产生叶绿素a, 叶绿素b和总叶绿素含量的变化情况, 以及96 h不同浓度DBP有无GO加入相对于空白组单个藻细胞叶绿素a水平对比. 图 6(a)~6(c)可以看出, DBP组和GO+DBP组藻细胞中叶绿素a、叶绿素b和总叶绿素含量要明显少于空白组, 且其变化趋势与藻密度一致.对比GO组与GO+DBP组0~96 h藻细胞中叶绿素a、叶绿素b和总叶绿素含量, 没有显著性差异. 图 6(d)中显示了各组96 h单个藻细胞叶绿素含量水平随DBP浓度变化情况(空白组为1), DBP浓度在0.1~4 mg·L-1时, 均表现出对叶绿素a无差异的高抑制性, 除了4 mg·L-1 DBP组, 其抑制率为21.48%. 0.1~2 mg·L-1 DBP组对藻叶绿素a含量的抑制率分别为85.15%、75.65%、76.79%、66.71%, 平均抑制率为76.08%. 0.1~4 mg·L-1 GO+DBP组对藻叶绿素a含量的抑制率分别为72.4%、82.24%、56.29%、68.03%、63.35%, 平均抑制率为68.46%.对比DBP组和GO+DBP组, GO的加入显著增强了高浓度DBP(4 mg·L-1)对单个藻细胞产生叶绿素a的抑制效应(P < 0.05), 这与GO对高浓度DBP(4 mg·L-1)藻密度的增强抑制效应一致.李芳芳等[27]研究纳米铜颗粒(10~30 nm)和微米铜颗粒(46 μm)对中肋骨条藻的毒性中发现, 铜离子、铜纳米颗粒微米铜颗粒都会抑制藻的光合作用, 且纳米铜颗粒对藻的毒性主要是浸出的铜离子造成的.

|
图 6 有无GO添加时DBP对青岛大扁藻光合色素的影响 Fig. 6 Effect of DBP, with and without the addition of GO, on the photosynthetic pigments of algae |
图 7显示了DBP单独及与1 mg·L-1 GO同时存在时青岛大扁藻细胞通透性的变化情况.从图 7(a)中可以看出, 0~96 h DBP单独及与1 mg·L-1 GO同时暴露下藻细胞通透性随时间呈现上升趋势, 在48 h后均高于空白组, DBP的暴露使青岛大扁藻的细胞通透性升高, 可能是由于DBP对细胞膜的破坏, 细胞内含物流出[25]. 图 7(b)可以看出96 h暴露后DBP组与GO+DBP组的藻细胞通透性均明显高于空白组, 0.1~4 mg·L-1DBP组藻细胞通透性分别为空白组的2.2、1.8、2.6、1.1、2.3倍, 平均升高到空白组的2.0倍, 0.1~4 mg·L-1 GO+DBP组藻细胞通透性分别为空白组的1.9、1.6、1.6、1.4、1.6倍, 平均升高到空白组的1.6倍.在DBP浓度为0.1、1和4 mg·L-1时, GO的加入对DBP表现出显著的藻细胞通透性减小作用(P < 0.05), 在DBP浓度为0.5 mg·L-1和2 mg·L-1时没有显著性影响. GO和DBP单独作用于青岛大扁藻时都能使藻细胞通透性增加, 但二者的作用机制不同, DBP对藻细胞通透性的影响主要是通过干扰细胞代谢造成膜损伤, 而GO对藻细胞通透性的影响主要是通过相互接触产生的物理性损伤[28].实验结果显示GO的加入减轻了DBP对藻细胞通透性的增高作用, 可能是由于藻类的胞外分泌物[29]及GO的吸附作用[19], 增强藻细胞间或与GO的相互积聚, 使藻细胞通透性比单独DBP作用时低.

|
图 7 有无GO添加时DBP对青岛大扁藻细胞膜通透性的影响 Fig. 7 Effect of DBP, with and without the addition of GO, on cell permeability |
氧化胁迫是解释有毒物质对生物组织或细胞造成毒性危害机制的主要理论之一.毒性物质进入生物活细胞后会导致大量的自由基产生, ROS含量上升, 造成氧化胁迫. SOD即总超氧化物歧化酶是一类高诱导酶, 是清除O2-自由基等系列反应中的关键酶, 能够将O2-自由基等底物转化为过氧化氢等过氧化物.为了探讨GO的加入是否能影响DBP对藻细胞中ROS和抗氧化系统损伤, 实验中考察了有无GO添加情况下, DBP暴露96 h后, 藻细胞内ROS和SOD的变化, 结果如图 8所示. DBP的暴露使ROS和SOD的水平明显升高, DBP组的ROS分别升高到空白组的1.5、1.9、1.3、1.3、1.7倍, 平均升高1.5倍. GO+DBP组的ROS分别升高到空白组的1.5、1.4、2.2、2.1、2.0倍, 平均升高1.9倍, 说明GO的加入使ROS的产量增加. DBP组的SOD分别升高到空白组的1.2、1.2、1.5、1.3、1.4倍, 平均升高1.3倍, GO+DBP组的SOD分别升高到空白组的1.4、1.2、1.4、1.2、1.9倍, 平均升高1.4倍, 说明GO的加入也使SOD的产量增加.在低浓度DBP组(0.1~2 mg·L-1), GO的加入对SOD水平无明显影响, 对ROS水平有明显增加, 但在高浓度DBP组(4 mg·L-1), GO的加入显著地提高了SOD水平, 而对ROS水平没有明显影响.细胞内ROS和SOD是一对相互反馈的变量, 当藻细胞内ROS升高时, 为了维持代谢平衡, 细胞中的SOD等抗氧化酶会被激活, 用于清除过量的ROS; 当SOD产量增加时, 藻细胞内的ROS会在短时间内被清除, 导致ROS含量下降.上述现象表明GO的加入, 会促进藻细胞体内ROS和SOD的产生, 这可能是GO对藻细胞的物理损伤能造成细胞表面的ROS升高[30], DBP对藻细胞内部结构的损伤导致细胞内ROS的升高, 两者共同作用导致氧化应激作用增强[25, 31].

|
图 8 有无GO添加时暴露96 h DBP对青岛大扁藻ROS和SOD的影响 Fig. 8 Effect of DBP, with and without the addition of GO, on the ROS and the SOD level in algae after exposure for 96 h |
图 9中显示了GO及DBP暴露下青岛大扁藻的SEM照片, 从图 9(b)~9(c)可以看出GO的加入对青岛大扁藻有一定的覆盖和包裹作用, 图中分别用黄色和蓝色箭头表示.前文中发现低浓度GO对青岛大扁藻的增长没有明显的影响, 但是对藻细胞通透性有显著影响, 随浓度的升高细胞通透性越高.由于浓度越高, GO与藻细胞的接触概率越大, 被划伤切割的概率也越大, 平均每个藻细胞通透性则增大.可以从藻密度、光合色素含量、SOD及ROS的变化中看出, 藻细胞的通透性增强, 使得DBP更容易进入藻细胞, 是导致DBP毒性增强的重要原因. Hu等[12]在对GO与小球藻相互作用的研究中, 所使用的GO在尺寸和浓度设定上与本实验相近, 他们发现GO在藻细胞表面形成泡状、薄膜、垂直嵌入、桥状等微型结构, 这些现象均能够改变藻细胞通透性.前文中提到, GO的加入减轻了DBP对藻细胞通透性的增强, 从图 9(d)中可以看出, DBP对青岛大扁藻的生长不仅有高抑制性, 还能导致青岛大扁藻畸形, 图中红色箭头所示, 畸变藻细胞比空白组藻细胞长3~7倍, 并且GO的存在使藻细胞容易发生图中类似的聚集. GO能对细胞膜产生应力, 影响细胞膜的生成[30], 畸形藻细胞或死亡后的藻细胞残留物均能与GO相互聚集, 聚集后覆盖在一些活细胞表面, 抑制藻细胞的运动, 影响物质的进出, 进而减小活细胞通透性, 改变藻细胞对营养物质的运输和摄取或相关代谢活动, 例如, 研究发现GO的存在可以抑制细胞中与Fe相关的代谢[26].

|
(a)为空白组青岛大扁藻; (b)为1 mg·L-1 GO单独作用组; (c)和(d)为1 mg·L-1 GO与4 mg·L-1DBP共同作用组; 图中黄色箭头表示GO覆盖在藻细胞表面, 蓝色箭头表示GO对藻细胞的包裹作用, 红色箭头表明藻体异常现象 图 9 GO及DBP暴露下青岛大扁藻的SEM图 Fig. 9 SEM images of the algae cells after exposure to DBP and GO |
(1) 低浓度0.1~10 mg·L-1 GO对青岛大扁藻的生长藻密度和光合色素产量无明显影响, 但GO的暴露能显著影响藻细胞通透性, 且随浓度升高而升高, 主要原因是GO与藻细胞的相互接触过程中造成的物理性伤害, GO浓度越高相互接触概率越大, 损伤也越大.
(2) 1 mg·L-1 GO的加入使DBP的EC50, 96 h从(11.14±0.80) mg·L-1降低到(4.93±2.14) mg·L-1, 1 mg·L-1GO对DBP藻毒性表现出一定的增强作用.
(3) 1 mg·L-1 GO的加入对低浓度DBP组(0.1~2 mg·L-1)暴露96 h后的藻密度、叶绿素、细胞通透性水平没有显著性影响, 但对高浓度DBP组(4 mg·L-1)的藻密度、叶绿素a抑制有显著的增强作用; GO的加入还使单个藻细胞氧化应激水平增加; 还发现GO的加入能减缓DBP造成的藻细胞通透性增加, 主要原因是GO的存在促进藻细胞团聚, 影响细胞运动和物质运输, 其分子机制仍需进一步研究.
| [1] | Liu N, Wen F L, Li F M, et al. Inhibitory mechanism of phthalate esters on Karenia brevis[J]. Chemosphere, 2016, 155: 498-508. |
| [2] |
别聪聪, 李锋民, 李媛媛, 等. 邻苯二甲酸二丁酯对短裸甲藻活性氧自由基的影响[J]. 环境科学, 2012, 33(2): 442-447. Bie C C, Li F M, Li Y Y, et al. Effects of allelochemical dibutyl phthalate on Gymnodinium breve reactive oxygen species[J]. Environmental Science, 2012, 33(2): 442-447. |
| [3] | Gao J, Chi J. Biodegradation of phthalate acid esters by different marine microalgal species[J]. Marine Pollution Bulletin, 2015, 99(1-2): 70-75. DOI:10.1016/j.marpolbul.2015.07.061 |
| [4] | Liu Y Y, Luo Y, Wu J, et al. Graphene oxide can induce in vitro and in vivo mutagenesis[J]. Scientific Reports, 2013, 3: 3469. DOI:10.1038/srep03469 |
| [5] | Li Y P, Wu Q L, Zhao Y L, et al. Response of microRNAs to in vitro treatment with graphene oxide[J]. ACS Nano, 2014, 8(3): 2100-2110. DOI:10.1021/nn4065378 |
| [6] | Hu C W, Wang Q, Zhao H T, et al. Ecotoxicological effects of graphene oxide on the protozoan Euglena gracilis[J]. Chemosphere, 2015, 128: 184-190. DOI:10.1016/j.chemosphere.2015.01.040 |
| [7] | Souza J P, Baretta J F, Santos F, et al. Toxicological effects of graphene oxide on adult zebrafish (Danio rerio)[J]. Aquatic Toxicology, 2017, 186: 11-18. DOI:10.1016/j.aquatox.2017.02.017 |
| [8] | Chen M J, Yin J F, Liang Y, et al. Oxidative stress and immunotoxicity induced by graphene oxide in zebrafish[J]. Aquatic Toxicology, 2016, 174: 54-60. DOI:10.1016/j.aquatox.2016.02.015 |
| [9] | Akhavan O, Ghaderi E. Toxicity of graphene and graphene oxide nanowalls against bacteria[J]. ACS Nano, 2010, 4(10): 5731-5736. DOI:10.1021/nn101390x |
| [10] | Hu W B, Peng C, Luo W J, et al. Graphene-based antibacterial paper[J]. ACS Nano, 2010, 4(7): 4317-4323. DOI:10.1021/nn101097v |
| [11] | Pretti C, Oliva M, Di Pietro R, et al. Ecotoxicity of pristine graphene to marine organisms[J]. Ecotoxicology and Environmental Safety, 2014, 101: 138-145. DOI:10.1016/j.ecoenv.2013.11.008 |
| [12] | Hu X G, Lu K C, Mu L, et al. Interactions between graphene oxide and plant cells:Regulation of cell morphology, uptake, organelle damage, oxidative effects and metabolic disorders[J]. Carbon, 2014, 80: 665-676. DOI:10.1016/j.carbon.2014.09.010 |
| [13] |
段伟艳, 杜永祥, 孟范平, 等. 氧化石墨烯对双壳类动物文蛤的亚致死毒性研究[J]. 中国环境科学, 2017, 37(7): 2755-2764. Duan W Y, Du Y X, Meng F P, et al. The sublethal toxicity of graphene oxide to bivalve Meretrix meretrix[J]. China Environmental Science, 2017, 37(7): 2755-2764. |
| [14] | Liao K H, Lin Y S, Macosko C W, et al. Cytotoxicity of graphene oxide and graphene in human erythrocytes and skin fibroblasts[J]. ACS Applied Materials & Interfaces, 2011, 3(7): 2607-2615. |
| [15] | Zhang H, Peng C, Yang J Z, et al. Uniform ultrasmall graphene oxide nanosheets with low cytotoxicity and high cellular uptake[J]. ACS Applied Materials & Interfaces, 2013, 5(5): 1761-1767. |
| [16] | Chang Y L, Yang S T, Liu J H, et al. In vitro toxicity evaluation of graphene oxide on A549 cells[J]. Toxicology Letters, 2011, 200(3): 201-210. DOI:10.1016/j.toxlet.2010.11.016 |
| [17] | Lu L, Wang J, Chen B L. Adsorption and desorption of phthalic acid esters on graphene oxide and reduced graphene oxide as affected by humic acid[J]. Environmental Pollution, 2018, 232: 505-513. DOI:10.1016/j.envpol.2017.09.078 |
| [18] | Liu Y, Wang X, Wang J, et al. Graphene oxide attenuates the cytotoxicity and mutagenicity of PCB 52 via activation of genuine autophagy[J]. Environmental Science & Technology, 2016, 50(6): 3154-3164. |
| [19] |
倪凌峰, 李轶, 郭燕飞. 氧化石墨烯存在下铜离子对大型溞的毒性研究[J]. 生态毒理学报, 2016, 11(5): 71-78. Ni L F, Li Y, Guo Y F. Toxicity of copper ions on Daphnia magna in the presence of graphene oxide[J]. Asian Journal of Ecotoxicology, 2016, 11(5): 71-78. |
| [20] | Ouyang S H, Hu X G, Zhou Q X. Envelopment-Internalization synergistic effects and metabolic mechanisms of graphene oxide on single-cell Chlorella vulgaris are dependent on the nanomaterial particle size[J]. ACS Applied Materials & Interfaces, 2015, 7(32): 18104-18112. |
| [21] |
张鹏. 石墨烯对植物的毒性效应及机制研究[D]. 杭州: 浙江工商大学, 2015. 19-30. Zhang P. Mechanisms of graphene toxicity to plants[D]. Hangzhou: Zhejiang Gongshang University, 2015. 19-30. http://cdmd.cnki.com.cn/Article/CDMD-10353-1016026866.htm |
| [22] | Nogueira P F M, Nakabayashi D, Zucolotto V. The effects of graphene oxide on green algae Raphidocelis subcapitata[J]. Aquatic Toxicology, 2015, 166: 29-35. DOI:10.1016/j.aquatox.2015.07.001 |
| [23] | Tang Y L, Tian J L, Li S Y, et al. Combined effects of graphene oxide and Cd on the photosynthetic capacity and survival of Microcystis aeruginosa[J]. Science of the Total Environment, 2015, 532: 154-161. DOI:10.1016/j.scitotenv.2015.05.081 |
| [24] |
向昆仑, 王晶晶, 唐思, 等. 双酚A对青岛大扁藻的干扰效应[J]. 生态毒理学报, 2015, 10(3): 262-267. Xiang K L, Wang J J, Tang S, et al. Interference effects of bisphenol A on Platymonas helgolanidica[J]. Asian Journal of Ecotoxicology, 2015, 10(3): 262-267. |
| [25] | Gu S R, Zheng H, Xu Q Q, et al. Comparative toxicity of the plasticizer dibutyl phthalate to two freshwater algae[J]. Aquatic Toxicology, 2017, 191: 122-130. DOI:10.1016/j.aquatox.2017.08.007 |
| [26] | Yu Q L, Zhang B, Li J R, et al. Graphene oxide significantly inhibits cell growth at sublethal concentrations by causing extracellular iron deficiency[J]. Nanotoxicology, 2017, 11(9-10): 1102-1114. DOI:10.1080/17435390.2017.1398357 |
| [27] |
李芳芳, 潘容, 张偲, 等. 纳米铜粉对中肋骨条藻的毒性效应[J]. 中国环境科学, 2015, 35(9): 2874-2880. Li F F, Pan R, Zhang C, et al. Inhibition effects of copper nanoparticles on the growth of Skeletonema costatum[J]. China Environmental Science, 2015, 35(9): 2874-2880. |
| [28] | Johnston H J, Hutchison G R, Christensen F M, et al. Identification of the mechanisms that drive the toxicity of TiO2 particulates:the contribution of physicochemical characteristics[J]. Particle and Fibre Toxicology, 2009, 6(1): 33. DOI:10.1186/1743-8977-6-33 |
| [29] | Iswarya V, Sharma V, Chandrasekaran N, et al. Impact of tetracycline on the toxic effects of titanium dioxide (TiO2) nanoparticles towards the freshwater algal species, Scenedesmus obliquus[J]. Aquatic Toxicology, 2017, 193: 168-177. DOI:10.1016/j.aquatox.2017.10.023 |
| [30] | Song C, Yang C M, Sun X F, et al. Influences of graphene oxide on biofilm formation of gram-negative and gram-positive bacteria[J]. Environmental Science and Pollution Research, 2018, 25(3): 2853-2860. DOI:10.1007/s11356-017-0616-8 |
| [31] |
刘春晓, 王平, 李海燕, 等. DBP对铜绿微囊藻生长和抗氧化酶的影响[J]. 环境科学与技术, 2015, 38(2): 7-12. Liu C X, Wang P, Li H Y, et al. Effects of dibutyl phthalate on growth and enzymatic antioxidants of Microcytis aeruginosa[J]. Environmental Science & Technology, 2015, 38(2): 7-12. |