 2018, Vol. 39
2018, Vol. 39



2. 山西财经大学环境经济学院, 太原 030006
2. Department of Environment and Economics, Shanxi University of Finance and Economics, Taiyuan 030006, China
群落构建研究对于解释物种共存机制和物种多样性维持机制至关重要, 一直是生态学研究的中心论题[1].在Vellend等[2]提出的概念模型中, 物种组成和多样性格局受到漂变(drift)、扩散(dispersal)、成种(speciation)和选择(selection)这4个基本生态过程的影响.其中, 前三者一定条件下倾向于随机过程, 而选择为确定性过程[3].目前的研究认为随机过程和确定性过程可以同时作用于群落构建, 二者之间形成此消彼长的连续体(continuum)[4].不同的生境、研究尺度、群落物种数量、生态位重叠及物种迁移能力等可能影响确定过程和随机过程对群落动态格局的相对重要性[5].但是, Clark[6]对这种连续体假说提出质疑, 他认为“随机性只是统计模型的一种属性, 而不是本质, 模型确实代表一种连续体, 但是没有证据表明群落结构动态就是由这个连续体引起的”.因此, 需要大量的研究工作对连续体假说进行验证.并且这一理论的依据主要来自植物群落, 关于动物和微生物群落的研究还需继续深入.
微生物个体微小, 扩散能力较强, 利用微生物对生态学理论(群落构建理论)进行验证, 取得了长足的进展.已有研究表明, 基于生态位理论的确定过程和基于中性理论的随机过程对微生物群落的构建有很大影响[7, 8].有学者研究温带森林土壤中微生物发现, 土壤根际微生物群落与植物群落的组成[9]、生产力[10]、根系密度[11]等环境因子密切相关.而真菌群落的分布更多地和土壤pH[12]、土壤营养[13]和海拔高度[14]等密切相关.另一方面, 随机作用(扩散)也对微生物群落产生很大影响.如对全球尺度和区域尺度[12]的外生菌根真菌的研究表明, 扩散过程影响了真菌群落的构建.在冰川消融的年代序列, 不但细菌和真菌群落展现出不同的构建过程[15], 而且真菌群落在不同的演替阶段也展现出不同的构建过程:确定过程的相对重要性会有阶段性的变化[8].然而, 关于微生物群落构建机制的研究在不同研究尺度和生境下的研究结论不尽一致[16, 17].因此, 需要大量的研究工作, 为群落构建机制的普适性提供一定的理论依据和数据支持.
本文选取关帝山庞泉沟自然保护区华北落叶松林土壤真菌群落为研究对象, 通过高通量测序技术, 探讨确定过程和随机过程的相对重要性及其对真菌群落结构的影响机制, 阐释亚高山针叶林土壤真菌群落的构建机制, 以期为生物多样性保护和森林管理措施的制定提供依据, 并为在不同尺度下研究生态系统演替和阐释微生物群落的构建机制提供理论基础.
1 材料与方法 1.1 研究区概况庞泉沟自然保护区位于山西省吕梁山脉的关帝山腹地, 是以保护世界珍禽褐马鸡(Crossoptilon mantchuricum)及其栖息地华北落叶松(Larix principis-rupprechtii)、云杉(Picea spp.)等天然次生林为主的森林和野生动物类型的自然保护区.样地设于具有明显海拔梯度差异的亚高山针叶林, 森林群落类型多样、结构差异明显, 是研究微生物群落结构和功能的理想场所.其地理位置111°22′~111°33′E, 37°45′~37°55′N, 海拔1 600~2 831 m, 主要森林类型是由针叶林华北落叶松、白扦(Picea meyeri)、青扦(Picea wilsonii)、油松(Pinus tabuliformis)以及白桦(Betula platyphylla)等树种组成的纯林和混交林.气候为暖温带大陆性山地气候, 年平均气温4.2℃, 年平均降水量822.6 mm, 土壤属于山地土壤类型, 从低到高垂直带谱为:山地褐土、山地淋溶褐土、山地棕壤和亚高山草甸土.
1.2 样品采集样地设置:本研究沿海拔梯度(2 000、2 100、2 200和2 400 m)设置4个华北落叶松群落调查样地(E2000、E2100、E2200和E2400, 见图 1.于2016年7月采样, 采用3点混合取样法采集土壤, 即在每一个采样地点设置1 m×1 m的样方, 利用对角线法选取3个点.首先去除地表约5 cm厚的地表植被和覆盖物, 再用土钻钻取0~20 cm深的土壤样品, 放入无菌塑料袋内, 于冰盒内保存, 尽快带回实验室, 并于-20℃下保存.在24 h之内, 将每个样地的3个土壤样品混合均匀并过直径为2 mm的网筛, 除去根系、石块等杂物, 其中一部分风干用于土壤理化性质的测定, 一部分放入-80℃保存, 用于高通量测序.

|
图 1 样地示意 Fig. 1 Schematic diagram of sampling plots |
自然风干的土样用于土壤理化性质的测定.土壤pH用HANNA HI3221测定(1:2.5土/水质量比); 土壤总氮(TN)、总碳(TC)和总硫(TS)用vario MACRO cube元素分析仪(德国)测定; 土壤硝态氮、铵态氮和亚硝态氮用CleverChem 380间断式元素分析仪(德国)测定; 土壤温度、土壤含水率、土壤盐度和电导率在野外采样时使用顺科达土壤温湿度仪TR-6D(中国)测定.
1.3.2 林下植被多样性分析野外采样时期, 记录每个样地内林下植被的种类、株数、盖度和高度.计算公式如下.
群落优势度指数:
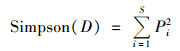
|
群落多样性指数:
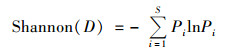
|
Pielou均匀度指数:
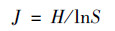
|
式中, S为植物群落丰富度指数; Pi(植物重要值)=(相对盖度+相对高度+相对多度)/3.
1.3.3 土壤真菌群落高通量测序分析每个样地的土壤取3个样本, 每个样本0.5 g.用E.Z.N.A.Soil DNA Kit试剂盒(Sangon公司, 中国)提取土壤微生物宏基因组DNA, 详细步骤参考试剂盒说明书.将每个土样提取出的3个DNA样品混合起来送往上海派森诺生物科技有限公司进行高通量测序.通过Illumina MiSeq测序平台, 利用引物ITS1F(CTTGGTCATTTAGAGGAAGTAA)和ITS2-Rev(GCTGCGTTCTTCATCGATGC)对核糖体ITS高变区进行测序.通常在97%的相似水平下对序列进行OTU的聚类和后续的生物信息分析.
1.4 数据处理各样地土壤理化变量之间的显著性差异均采用单因素方差分析(one-way analysis)和多重比较分析(LSD), 土壤环境因子和真菌优势门的相关性采用Pearson相关性分析, 并通过R语言(V3.4.1)程序包corrplot作可视化分析.土壤环境因子、地上植被多样性与微生物群落间的冗余分析(RDA)使用vegan程序包分析, 真菌群落相对丰度使用ggplot2程序包可视化分析; 韦恩图使用程序包VennDiagram完成; 对相对丰度前50的真菌目进行热图分析, 采用R语言gplots程序包完成.
零模型分析采用Chase等[18, 19]所构建的方法.使用基于β多样性的零模型方法, 定量分析确定过程和随机过程对群落构建的相对重要性.β多样性零偏差方法是通过建立零模型分析观测值相似性偏离期望值相似性的程度[(βobs-βnull)/βnull]来定量分析确定作用与随机作用的相对重要性.
2 结果与分析 2.1 土壤理化性质土壤理化性质测定与分析结果(表 1)显示, 土壤含水率、土壤盐度、电导率、铵态氮、亚硝态氮和pH值在不同样地间差异显著, 土壤温度、TN、TC、TS、硝态氮在样地间差异不显著.低海拔样地E2000土壤含水率(24.1%)、土壤盐度(43 mg·L-1)、电导率(78.33 μS·cm-1)最高, 高海拔样地较小.亚硝态氮在样地E2400(0.167 mg·kg-1)含量最多, 在样地E2000含量最少(0.026 7 mg·kg-1), 铵态氮在样地E2000含量最多(25.6 mg·kg-1), 在样地E2400含量最少(11.12 mg·kg-1).所有样地的pH值都小于7, 为弱酸性的土壤, 且pH值在样地E2100最大(7.23).结果表明, 沿海拔梯度土壤理化因子差异显著, 形成了一定的生态梯度.本研究基于这一生态梯度, 针对华北落叶松林土壤真菌群落结构动态的驱动因素进行分析.
|
|
表 1 土壤环境因子及林下植被多样性1) Table 1 Soil environment factors and underground vegetation diversity of the analyzed soil samples |
2.2 林下植被群落多样性
林下植被多样性指数在各样地之间具有显著差异(表 1).辛普森指数在样地E2200最高(1.516), 在样地E2000最低(0.7748);香农-维纳指数在E2400最高(0.746), 在样地E2100最低(0.378);均匀度指数(0.814)在样地E2400最高, 在样地E2000最低(0.503 5).由此可见, 在上述生态梯度上, 高海拔样地的植物多样性指数高于低海拔样地的多样性指数.
2.3 土壤真菌群落组成及其多样性各样地真菌群落α多样性分析结果如表 2所示, 样地E2400的辛普森指数(0.96)和香农-维纳指数(6.28)最高, E2000的辛普森指数(0.85)和香农-维纳指数(4.55)最低.在样地E2400 ACE指数(391.66)和Chao1指数(390.64)最高, 在样地E2000最低.在上述的生态梯度上, 海拔梯度对真菌群落多样性也有一定的影响.
|
|
表 2 土壤真菌群落多样性 Table 2 Diversity of soil fungal communities |
真菌群落系统发育树(图 2)显示, 真菌群落主要包括子囊菌门(Ascomycota)、担子菌门(Basidiomycota)、接合菌门(Zygomycota)、球囊菌门(Glomeromycota)和壶菌门(Chytridiomycota)等.本研究区总共鉴定出7个真菌门和94个真菌属.真菌物种分布格局具有一定的规律性(图 3):子囊菌门和担子菌门在所有样地中相对丰度最大, 为优势菌门.子囊菌门的相对丰度随着海拔的升高而升高, 担子菌门的相对丰度随海拔对升高而降低.在目水平上的韦恩图表明4个样地共有的真菌目为36个, 且相邻两个的样地间也都有共有的真菌目.通过将相对丰度前50的真菌目热图分析, 样地E2400、E2200和E2100的真菌群落结构可以聚为一类, 说明样地E2400、E2200、E2100真菌群落结构较为类似, 与低海拔样地E2000真菌群落有一定差异.海拔梯度对真菌群落结构有显著的影响.

|
图 2 土壤真菌群落系统发育树 Fig. 2 Phylogenetic tree of soil fungal communities |

|
(a)优势真菌门的相对丰度;(b)韦恩图;(c)热图 图 3 优势真菌门和优势真菌目在各样地的分布格局示意 Fig. 3 Schematic diagram of the distribution pattern of dominant fungal phyla and orders across the plots |
通过偏最小二乘法(PLSPM)构建结构方程模型(图 4), 分析土壤理化因子对真菌群落的直接影响和间接影响(拟合优度GOF=0.855 4).其中, 土壤理化因子包括盐度、电导率、碳、铵态氮、硝态氮和亚硝态氮(上述分析可知, 和真菌群落呈显著相关); 林下植被多样性包括辛普森指数、香农指数和均匀度; 真菌群落主要包括担子菌门和子囊菌门(优势菌门).结果表明, 林下植被多样性对真菌群落的直接影响最明显(1.185 8), 其次是海拔梯度(-0.517 3)和土壤因子(-0.466 5).

|
基于偏最小二乘法(partial least squares path modeling)的结构方程模型分析环境影子对真菌群落的直接影响; 方程的拟合优度(GOF)为0.855 4;图中数字表示相关系数, 数字值越大, 直接影响越大; 正值代表促进作用, 负值代表抑制作用 图 4 环境因子对真菌群落贡献的结构方程模型 Fig. 4 Equation structural model for the contribution of environmental factors to fungal communities |
通过优势真菌门丰度和土壤理化因子间的相关分析[图 5(a)], 子囊菌门与土壤盐度、铵态氮呈显著负相关(P < 0.05), 与亚硝态氮呈显著正相关; 担子菌门与铵态氮呈显著正相关, 与土壤总碳、亚硝态氮呈显著负相关.由此推断, 土壤盐度、铵态氮、亚硝态氮和碳对真菌群落结构有显著影响.

|
图 5 优势真菌门与土壤环境因子相关矩阵和优势真菌门与土壤环境因子及林下植被多样性指数的冗余分析 Fig. 5 Correlation matrix diagram of dominant fungal phyla and soil environmental factors, and ordination diagram of RDA with dominant soil fungal communities and explanatory variables |
对本研究区土壤中的优势真菌门丰度和土壤环境因子及林下植被多样性指数进行冗余分析(RDA)[图 5(b)], 结果显示第1轴和第2轴累积解释率达到98%, 说明土壤理化因子和地上植被多样性对土壤真菌群落结构影响较大.林下植被多样性均匀度、香农指数和土壤温度对子囊菌门、接合菌门和球囊菌门影响较大, 铵态氮和盐度对担子菌门影响较大, 硝态氮对Rozellomycota影响较大.
由此推断, 地上植被多样性和土壤环境因子对真菌群落的构建过程有很重要的影响作用.而地上植被与土壤环境因子都属于确定过程.因此, 确定过程对土壤真菌群落的构建有很大的影响.
2.5 地理距离对土壤真菌群落的影响研究区真菌群落β多样性与地理距离的关系如图 6所示, 二者之间具有较强的相关性(R2=0.870 8), 且真菌群落的β多样性随着地理距离的增加而减小, 说明一定程度上地理距离能够影响森林土壤真菌群落的β多样性.由此推断, 随机过程(扩散限制)对土壤真菌群落构建也有一定的影响作用.

|
图 6 真菌群落的距离衰减分布格局 Fig. 6 Distribution pattern of the distance-delay of the fungi community |
上述分析可推断, 确定过程(环境因子)和随机过程(扩散限制)对真菌群落构建都有一定的影响作用.通过基于β多样性的零模型分析结果如图 7所示, 零偏差值在各个样地都是负值, 表明确定过程驱动了研究区域真菌群落的构建过程.零偏差值可以代表群落的相似性比期望值更加相似(负值), 或者更加相异(正值); 零偏差值越接近零, 表明随机过程驱动群落构建过程; 越偏离零, 表明确定过程驱动了群落的构建.而且观测值的相似性小于预测期望值的相似性, 说明生境过滤是真菌群落构建的主要驱动因子.零偏差值在不同的样地有差异, 说明确定过程与随机过程的相对重要性随着海拔梯度的变化而变化.

|
图 7 真菌群落在不同样地的零模型 Fig. 7 Null model of the fungi community across the plots |
本研究基于局域尺度, 土壤理化因子和植被多样性在海拔梯度上有显著的差异, 形成一定的生态梯度.在此生态梯度上, 探讨确定过程和随机过程的权衡作用对真菌群落构建过程的驱动机制.
通过构建结构方程模型(PLS-PM)分析, 显示林下植被多样性对真菌群落的直接影响最明显, 其次是海拔梯度和土壤因子.Yang等[20]通过最小二乘回归(PLSR)研究青藏高原自然草原土壤真菌多样性, 发现植物丰富度和真菌丰富度显著相关; 也有研究表明, 在区域尺度上, 植物群落的组成可以预测真菌群落的分布, 而且相比于非生物因素, 植物群落对真菌群落的影响更大[21].本研究的结果与上述类似, 在局域尺度, 植被多样性对真菌群落直接影响最显著.一方面, 林下植被凋落物通过被土壤中的腐生微生物分解给土壤提供养分, 这些养分又给真菌的生长繁殖提供营养, 营养元素在真菌与林下植被之间形成循环; 另一方面, 某些土壤真菌能够与植物共生或形成菌根真菌, 这种种间合作的方式提高了合作土壤真菌的竞争能力, 从而在有限的资源里提高群落的丰富度.另外也有研究表明, 在真菌分解实验中枯枝落叶具有增效作用[22].因此林下植被的多样性变化趋势与土壤真菌群落结构显著相关, 植被多样性对土壤真菌遗传多样性有显著的作用[23].
对土壤理化因子和优势真菌门进行相关性分析和冗余分析, 表明土壤盐度、铵态氮、亚硝态氮和总碳等因子对真菌群落有显著影响.前人研究也表明森林生态系统中真菌群落的分布与某些非生物因子, 比如土壤营养等密切相关[13].这可能与下述几点有关:寒温性华北落叶松林凋落物较多, 凋落物中多含难以分解的木质素、单宁、树脂和蜡质等物质, 而子囊菌门和担子菌门是土壤真菌群落中主要的分解者[24, 25], 因此在本研究区相对丰度较高.担子菌门相对丰度随着海拔的升高而降低, 可能由于它对高海拔样地的土壤环境的异质性适应较差.另一方面, 真菌群落对土壤含水率的变化很敏感, 土壤含水率主要影响植物病原体以及共生体等与植物相关的真菌[26].E2400样地土壤铵态氮浓度最低, 这可能是因为高海拔针叶林下比低海拔土壤更加冷湿, 不利于氮矿化和硝化作用的进行[27].
海拔梯度对真菌群落结构有较大的影响.Bahram[14]研究伊朗Hyrcanian森林真菌群落指出, 真菌群落物种丰富度随着海拔的升高而降低, 且在局域尺度和区域尺度上, 海拔是外生菌根真菌的主要驱动因素.本研究中真菌群落的丰富度随着海拔的升高而升高, 一方面这可能是因为本研究区是暖温带亚高山华北落叶松林, 海拔梯度对真菌群落组成造成中度干扰, 提高了群落丰富度.
距离衰减关系, 即群落分类相异性随地理距离的增加而增加, 是微生物群落普遍的生物地理分布格局[28].有研究发现, 即使在局域尺度, 微生物群落也展现出距离衰减的分布格局[29].本研究中, 真菌群落表现出距离衰减的生物地理分布格局, 即群落的相异性随着地理距离的增加而增加, 这与前人的研究结果相类似.扩散限制对真菌群落的构建产生一定的影响, 由此推断随机过程对真菌群落也有一定的影响.
有研究认为群落构建是物种的筛选过程, 环境因子和生物间的相互作用则是多个嵌套的筛子(nested sieves), 区域物种库中只有具有特定性状的物种, 才能通过这些嵌套筛子的过滤选入局域群落[30~32].生境过滤作用由于是将适宜物种筛入相同生态位的过程, 增加物种间的相似性, 因而结果会使得功能性状的趋同聚集(trait convergence)[33, 34].即群落相似性越高, 越能体现生境过滤作用对群落构建的影响作用.Tucker等指出[35], 零模型的零偏差值可以代表群落的相似性比期望值更加相似(负值), 或者更加相异(正值).由于零模型是分析观测值相似性偏离期望值相似性的程度[(βobs-βnull)/βnull], 且零偏差值在各个样地都是负值, 说明群落的相似性比期望值更相似, 因此笔者推断生境过滤作用是本研究区真菌群落构建的驱动因子.相关研究指出, 真菌群落的构建由确定过程(pH、盐度、土壤肥力和有机碳)[36~39]、随机过程[40](扩散限制)或者两者权衡作用[41]的影响.也有研究指出, 在原生演替时期, 细菌群落更多地由确定过程驱动, 而真菌群落更倾向于随机过程.不同时空尺度和类型的群落可能受不同的生态过程驱动[42].本研究基于局域尺度, 环境梯度明显, 生境过滤是真菌群落构建的驱动因子.
4 结论本文以关帝山庞泉沟自然保护区华北落叶松针叶林土壤真菌群落为研究对象, 在局域尺度探讨真菌群落构建机制.研究发现, 土壤理化因子和植被多样性中沿海拔梯度上差异显著, 形成一定的生态梯度, 确定过程(生境过滤)驱动本研究区真菌群落的构建.
| [1] |
柴永福, 岳明. 植物群落构建机制研究进展[J]. 生态学报, 2016, 36(15): 4557-4572. Chai Y F, Yue M. Research advances in plant community assembly mechanisms[J]. Acta Ecologica Sinica, 2016, 36(15): 4557-4572. |
| [2] | Vellend M. Conceptual synthesis in community ecology[J]. The Quarterly Review of Biology, 2010, 85(2): 183-206. DOI:10.1086/652373 |
| [3] | Hubbell S P. The unified neutral theory of biodiversity and biogeography[J]. Plant Systematics & Evolution, 2002, 85(11): 3172-3174. |
| [4] |
牛克昌, 刘怿宁, 沈泽昊, 等. 群落构建的中性理论和生态位理论[J]. 生物多样性, 2009, 17(6): 579-593. Niu K C, Liu Y N, Shen Z H, et al. Community assembly:the relative importance of neutral theory and niche theory[J]. Biodiversity Science, 2009, 17(6): 579-593. |
| [5] | Chase J M, Myers J A. Disentangling the importance of ecological niches from stochastic processes across scales[J]. Philosophical Transactions of the Royal Society B:Biological Science, 2011, 366(1576): 2351-2363. DOI:10.1098/rstb.2011.0063 |
| [6] | Clark J S. Beyond neutral science[J]. Trends in Ecology & Evolution, 2009, 24(1): 8-15. |
| [7] | Ferrenberg S, O'neill S P, Knelman J E, et al. Changes in assembly processes in soil bacterial communities following a wildfire disturbance[J]. The ISME Journal, 2013, 7(6): 1102-1111. DOI:10.1038/ismej.2013.11 |
| [8] | Tian J Q, Qiao Y C, Wu B, et al. Ecological succession pattern of fungal community in soil along a retreating glacier[J]. Frontiers in Microbiology, 2017, 8: 1028. DOI:10.3389/fmicb.2017.01028 |
| [9] | Roy M, Rochet J, Manzi S, et al. What determines Alnus-associated ectomycorrhizal community diversity and specificity? A comparison of host and habitat effects at a regional scale[J]. New Phytologist, 2013, 198(4): 1228-1238. DOI:10.1111/nph.12212 |
| [10] | Kranabetter J M, Durall D M, MacKenzie W H. Diversity and species distribution of ectomycorrhizal fungi along productivity gradients of a southern boreal forest[J]. Mycorrhiza, 2009, 19(2): 99-111. DOI:10.1007/s00572-008-0208-z |
| [11] | Peay K G, Kennedy P G, Bruns T D. Rethinking ectomycorrhizal succession:are root density and hyphal exploration types drivers of spatial and temporal zonation?[J]. Fungal Ecology, 2011, 4(3): 233-240. DOI:10.1016/j.funeco.2010.09.010 |
| [12] | Põlme S, Bahram M, Yamanaka T, et al. Biogeography of ectomycorrhizal fungi associated with alders (Alnus spp.) in relation to biotic and abiotic variables at the global scale[J]. New Phytologist, 2013, 198(4): 1239-1249. DOI:10.1111/nph.12170 |
| [13] | Cox F, Barsoum N, Lilleskov E A, et al. Nitrogen availability is a primary determinant of conifer mycorrhizas across complex environmental gradients[J]. Ecology Letters, 2010, 13(9): 1103-1113. DOI:10.1111/j.1461-0248.2010.01494.x |
| [14] | Bahram M, Põlme S, Kõljalg U, et al. Regional and local patterns of ectomycorrhizal fungal diversity and community structure along an altitudinal gradient in the Hyrcanian forests of northern Iran[J]. New Phytologist, 2012, 193(2): 465-473. DOI:10.1111/j.1469-8137.2011.03927.x |
| [15] | Rime T, Hartmann M, Frey B. Potential sources of microbial colonizers in an initial soil ecosystem after retreat of an alpine glacier[J]. The ISME Journal, 2016, 10(7): 1625-1641. DOI:10.1038/ismej.2015.238 |
| [16] | Ranjard L, Dequiedt S, Prévost-Bouré N C, et al. Turnover of soil bacterial diversity driven by wide-scale environmental heterogeneity[J]. Nature Communications, 2013, 4: 1434. DOI:10.1038/ncomms2431 |
| [17] | Friedman J, Higgins L M, Gore J. Community structure follows simple assembly rules in microbial microcosms[J]. Nature Ecology & Evolution, 2017, 1(5): 109. |
| [18] | Chase J M. Drought mediates the importance of stochastic community assembly[J]. Proceedings of the National Academy of Sciences of the United States of America, 2007, 104(44): 17430-17434. DOI:10.1073/pnas.0704350104 |
| [19] | Chase J M, Kraft N J B, Smith K G, et al. Using null models to disentangle variation in community dissimilarity from variation in α-diversity[J]. Ecosphere, 2011, 2(2): 24. |
| [20] | Yang T, Adams J M, Shi Y, et al. Soil fungal diversity in natural grasslands of the Tibetan Plateau:associations with plant diversity and productivity[J]. New Phytologist, 2017, 215(2): 756-765. DOI:10.1111/nph.2017.215.issue-2 |
| [21] | Ke P J, Miki T, Ding T S. The soil microbial community predicts the importance of plant traits in plant-soil feedback[J]. New Phytologist, 2015, 206(1): 329-341. DOI:10.1111/nph.13215 |
| [22] | Gartner T B, Cardon Z G. Decomposition dynamics in mixed-species leaf litter[J]. Oikos, 2004, 104(2): 230-246. DOI:10.1111/oik.2004.104.issue-2 |
| [23] | Rousk J, Brookes P C, Bååth E. Contrasting soil pH effects on fungal and bacterial growth suggest functional redundancy in carbon mineralization[J]. Applied and Environmental Microbiology, 2009, 75(6): 1589-1596. DOI:10.1128/AEM.02775-08 |
| [24] | Yelle D J, Ralph J, Lu F C, et al. Evidence for cleavage of lignin by a brown rot basidiomycete[J]. Environmental Microbiology, 2008, 10(7): 1844-1849. DOI:10.1111/j.1462-2920.2008.01605.x |
| [25] | Bossuyt H, Denef K, Six J, et al. Influence of microbial populations and residue quality on aggregate stability[J]. Applied Soil Ecology, 2001, 16(3): 195-208. DOI:10.1016/S0929-1393(00)00116-5 |
| [26] | Kivlin S N, Hawkes C V. Tree species, spatial heterogeneity, and seasonality drive soil fungal abundance, richness, and composition in Neotropical rainforests[J]. Environmental Microbiology, 2016, 18(12): 4662-4673. DOI:10.1111/1462-2920.13342 |
| [27] |
沙丽清, 孟盈, 冯志立, 等. 西双版纳不同热带森林土壤氮矿化和硝化作用研究[J]. 植物生态学报, 2000, 24(2): 152-156. Sha L Q, Meng Y, Feng Z L, et al. Nitrification and net N mineralization rate of soils under different tropical forests in Xishuangbanna, Southwest China[J]. Acta Phytoecologica Sinica, 2000, 24(2): 152-156. |
| [28] | Martiny J B H, Bohannan B J M, Brown J H, et al. Microbial biogeography:putting microorganisms on the map[J]. Nature Reviews Microbiology, 2006, 4(2): 102-112. DOI:10.1038/nrmicro1341 |
| [29] | Langenheder S, Lindström E S, Tranvik L J. Structure and function of bacterial communities emerging from different sources under identical conditions[J]. Applied and Environmental Microbiology, 2006, 72(1): 212-220. |
| [30] | Keddy P A. Assembly and response rules:two goals for predictive community ecology[J]. Journal of Vegetation Science, 1992, 3(2): 157-164. DOI:10.2307/3235676 |
| [31] | Weiher E, Keddy P A. Assembly rules, null models, and trait dispersion:new questions from old patterns[J]. Oikos, 1995, 74(1): 159-164. DOI:10.2307/3545686 |
| [32] | Weiher E, Clarke G D P, Keddy P A. Community assembly rules, morphological dispersion, and the coexistence of plant species[J]. Oikos, 1998, 81(2): 309-322. DOI:10.2307/3547051 |
| [33] | Bazzaz F A. Habitat selection in plants[J]. The American Naturalist, 1991, 137: S116-S130. DOI:10.1086/285142 |
| [34] | Lavorel S, Garnier E. Predicting changes in community composition and ecosystem functioning from plant traits:revisiting the Holy Grail[J]. Functional Ecology, 2002, 16(5): 545-556. DOI:10.1046/j.1365-2435.2002.00664.x |
| [35] | Tucker C M, Shoemaker L G, Davies K F, et al. Differentiating between niche and neutral assembly in metacommunities using null models of β-diversity[J]. Oikos, 2016, 125(6): 778-789. DOI:10.1111/oik.2016.v125.i6 |
| [36] | Fierer N, Jackson R B. The diversity and biogeography of soil bacterial communities[J]. Proceedings of the National Academy of Sciences of the United States of America, 2006, 103(3): 626-631. DOI:10.1073/pnas.0507535103 |
| [37] | Lozupone C A, Knight R. Global patterns in bacterial diversity[J]. Proceedings of the National Academy of Sciences of the United States of America, 2007, 104(27): 11436-11440. DOI:10.1073/pnas.0611525104 |
| [38] | Hartman W H, Richardson C J, Vilgalys R, et al. Environmental and anthropogenic controls over bacterial communities in wetland soils[J]. Proceedings of the National Academy of Sciences of the United States of America, 2008, 105(46): 17842-17847. DOI:10.1073/pnas.0808254105 |
| [39] | Nemergut D R, Costello E K, Hamady M, et al. Global patterns in the biogeography of bacterial taxa[J]. Environmental Microbiology, 2011, 13(1): 135-144. DOI:10.1111/emi.2011.13.issue-1 |
| [40] | Peay K G, Garbelotto M, Bruns T D. Evidence of dispersal limitation in soil microorganisms:isolation reduces species richness on mycorrhizal tree islands[J]. Ecology, 2010, 91(12): 3631-3640. DOI:10.1890/09-2237.1 |
| [41] | Wu B, Tian J Q, Bai C M, et al. The biogeography of fungal communities in wetland sediments along the Changjiang River and other sites in China[J]. The ISME Journal, 2013, 7(7): 1299-1309. DOI:10.1038/ismej.2013.29 |
| [42] | Dini-Andreote F, Pylro V S, Baldrian P, et al. Ecological succession reveals potential signatures of marine-terrestrial transition in salt marsh fungal communities[J]. The ISME Journal, 2016, 10(8): 1984-1997. DOI:10.1038/ismej.2015.254 |