 2018, Vol. 39
2018, Vol. 39



2. 安徽农业大学农学院, 合肥 230036
2. College of Agronomy, Anhui Agricultural University, Hefei 230036, China
农业是大气温室气体N2O的重要来源且与氮肥施用有关[1, 2].在氮肥施用量较高的我国菜田土壤, N2O排放占全国N2O直接排放量的20%[3].与大田作物相比, 农民习惯于对设施菜田投入更高的养分和水[4].过高的有机肥、化肥投入和频繁灌溉在提高土壤微生物活性的同时, 通常会显著降低土壤孔隙的氧含量, 造成土壤微生物常处于低氧胁迫的环境中.低氧胁迫会显著影响硝化和反硝化过程, 进而影响土壤N2O的排放[5, 6].
氧气是影响土壤N2O排放的重要参数, 一般认为, 厌氧条件下异养反硝化过程是土壤N2O排放的主要途径, 即使很低的氧含量也会显著抑制异养反硝化过程[7, 8], 导致有氧条件下硝化过程是土壤N2O排放的主导途径[9~11].Zhu等[10]的研究结果显示氧气体积分数介于0.5%~3%时, 硝化和硝化细菌的反硝化对N2O的贡献量达到34%~84%, 同样, Frame等[12]认为在0.5%的氧浓度下, 硝化细菌反硝化作用对N2O的贡献量达到43%~87%.但也有研究表明, 即使在2%[13]、3%[14]、甚至5%[15]氧气体积分数下, 仍可以观察到显著的N2O还原现象, 低氧条件下N2O的产生途径始终存在争议.由于氧气在培养过程中低的可控性, 大部分关于氧气对N2O排放影响的研究主要针对单菌株或土壤细胞提取物[16~18], 低氧条件下土壤氧气动态变化与N2O排放间的关系研究较少.此外, NO2-作为N2O产生的重要底物, 与土壤N2O的产生存在相关性[19, 20], 通过NaClO3的添加抑制NO2-氧化菌(NOB)的活性[21], 有助于深入分析低氧条件下N2O的产生机制.
设施菜田中通常会采用添加高碳氮比秸秆的方法改善土壤质量, 并促进N的生物固持[22], 这也同时会影响土壤N2O的排放[23].然而, 与未添加秸秆的土壤相比, 长期添加作物秸秆的土壤是否也有高的N2O产生潜势, 目前还不清楚.此外, 不同碳投入土壤对氧气的响应研究更少.
为了探索低氧条件下设施菜田土壤N2O的产生途径及大量排放的原因, 本文以中国北方典型的设施菜田土壤为研究对象, 在不同初始氧气体积分数下(0%、1%、3%、5%和10%), 利用在线自动监测培养系统(Robot系统)定量测定土壤N2O、NO、N2和CO2产生动态, 并分析其N2O/(NO+N2O+N2)指数, 同时, 动态监测了NO2-、NO3-和NH4+的含量.此外, 通过利用NaClO3抑制NO2-的氧化, 进一步了解低氧条件下N2O产生与土壤NO2-累积间的关系, 以期为菜田土壤N2O减排措施的制定提供理论依据.
1 材料与方法 1.1 采样地点及土壤样品采集供试土壤采自中国农业大学山东寿光蔬菜研究院的定位试验日光温室, 该试验始于2007年, 详细描述见Fan等[24]的研究.该试验是一个裂区试验, 主处理为滴灌和漫灌, 副处理为不添加秸秆、添加小麦秸秆和添加玉米秸秆.本研究选取的土壤样品来自于滴灌(DI)和滴灌+玉米秸秆(DIS)两个处理.滴灌处理每年平均灌溉量为309 mm, 商品鸡粪作为基肥, 平均施用量相当于210 kg·(hm2·季)-1的氮, DIS处理会额外投入高碳氮比的玉米秸秆8 t·hm-2(相当于109 kg·hm-2氮以及3 406 kg·hm-2碳)以调节土壤碳氮平衡.2014年7月番茄收获后采集0~20 cm土壤样品, 充分混匀后过2 mm筛风干备用.
1.2 试验设计与方法为了减少土壤硝态氮(NO3-)背景值含量过高对试验结果的干扰, 同时避免厌氧条件下NO超标现象的发生, 试验开始前将DI、DIS土壤用去离子水清洗4次, 使其NO3-含量低于20 mg·kg-1, 待水分自然蒸发至含水量质量分数10%~15%之间时, 过2 mm筛, 随后将过筛土置于4℃冰箱储存备用.该土壤样品的黏粒、粉粒、沙粒所占比例分别为29%、52%和19%, 洗土后土壤基础理化性质见表 1.
|
|
表 1 土壤基础理化性质 Table 1 Physical and chemical properties of soil |
试验一:以DIS土壤为研究对象, 称取相当于20 g烘干土壤置于120 mL血清瓶, 根据实际测定的土壤NH4+、NO3-含量, 向每个血清瓶补加19 mg·kg-1 NH4+ [(NH4)2SO4]及12 mg·kg-1NO3- (KNO3), 使供试土壤NH4+、NO3-含量为30 mg·kg-1左右, 控制土壤含水量质量分数为25%.所有血清瓶用铝盖密封后用He气通过抽真空-洗气系统(北京帅恩科技有限公司)洗气3次, 分别充入0%、1%、3%、5%以及10%的O2/He混合气体, 随后将所有血清瓶置于20℃恒温水浴槽中培养, 利用Robot系统[25]每隔6 h测定一次培养瓶顶空气体(O2、NO、N2O、N2、CO2)的变化情况.同时, 根据气体监测结果, 选取10个气体产生关键点, 对相同培养条件下的静态培养试验进行破坏性采样, 测定培养过程中无机氮(NH4+、NO3-和NO2-)的动态变化.总培养时间为216 h, 每个处理3个重复.
试验二:以DI土壤为研究对象, 称取相当于20 g烘干土壤置于120 mL血清瓶, (NH4)2SO4及KNO3添加量与试验一相同, 只是增加了添加NaClO3(20 mmol·L-1)[26]的处理, 调节土壤含水量质量分数为25%.所有血清瓶用铝盖密封后用He气通过抽真空-洗气系统(北京帅恩科技有限公司)洗气3次, 分别充入0%、1%、5%以及10%的O2/He混合气体, 随后将所有血清瓶置于20℃恒温水浴槽中培养.利用Robot自动监测系统每隔8 h测定O2、NO、N2O、N2和CO2产生量, 总培养时间为224 h.
1.3 气体动态监测及土壤无机氮素含量、pH值测定Robot自动培养系统由自动进样和气体分析模块组成.其中自动进样模块包括双向旋转的蠕动泵(Gilson Model 222, Gilson, 法国)和顶空自动采样器(CTC GC-Pal).气体分析模块包括气相色谱(Agilent 7890A)和氮氧化物分析仪(Model 200E, 美国).而气相色谱中包含热导检测器(TCD)、电子捕获检测器(ECD)和火焰离子化检测器(FID)这3个检测器, 能够监测N2O(ECD、TCD)、N2(TCD)、CO2(TCD)和O2(TCD)气体浓度的变化.氮氧化物分析仪用来定量监测NO气体浓度.相关Robot自动培养系统的运行模块细节可参见Molstad等[25]的描述.
土壤样品中NH4+和NO3-的含量用1mol·L-1 KCl溶液浸提后用连续流动分析仪(TRACCS2000, 德国)测定; 土壤亚硝态氮测定方法按照国标HJ634-2012方法.土壤全氮、全碳含量利用碳氮分析仪测定(Thermo Scientific Flash 2000 NC Analyzer, 美国).有机碳含量采用重铬酸钾-硫酸溶液氧化滴定法测定.速效磷含量采用NaHCO3浸提-钼锑抗吸光光度法测定.速效钾含量采用NH4OAc浸提-火焰光度法测定.土壤pH值测定的水土比为2.5:1, 用pH计测定.
1.4 数据处理不同氧气体积分数下N2O/(N2O+NO+N2)指数计算公式为[18]:
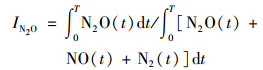
|
(1) |
式中, N2O(t)、N2(t)、NO(t)均为N2O测量值达到高峰时间(t)下的累积产生量.质量平衡是指培养结束时土壤无机氮(NH4+、NO3-)、气体产生量(N2O、N2)之和与培养开始时土壤无机氮(NH4+、NO3-)、气体产生量(N2O、N2)之和两者间的差值.数据采用SPSS 20.0进行单因素、多因素方差分析.采用Sigmaplot 12.5作图, 图表中数据均为:平均值±标准误差.
2 结果与分析 2.1 DIS土壤不同初始氧气体积分数下无机氮、气体的动态变化由图 1可见, 在完全厌氧条件下, DIS土壤中NO3-在培养48 h后全部被还原, 土壤NO2-含量呈先增加后下降的趋势.土壤NO和N2O产生量分别在培养54 h和90 h时达到最大累积量, N2产生速率在114 h时达到峰值73.06 nmol·(g·h)-1.在120 h时, 土壤中NO3-被完全还原.

|
图中所有的气体、无机氮含量均以N计,下同 图 1 不同初始氧气体积分数下DIS土壤气体、无机氮的动态变化 Fig. 1 Dynamic changes in nitrogenous gases and mineral nitrogen contents under different initial volume fractions of oxygen in DIS soil |
初始氧气体积分数为1%时, 培养前期(1~32 h)土壤NH4+含量呈下降趋势, 而NO2-和NO3-的含量分别增加了6.49 mg·kg-1和2.58 mg·kg-1.从48 h开始(氧气体积分数为0.36%), 土壤NO3-含量显著降低, NO2-含量在64 h时达到峰值16.71 mg·kg-1, 随后于80 h开始降低.随着氧气的逐渐消耗, NO测量值在102 h时达到高峰, 同时, N2O产生速率在64 h(氧气体积分数为0.019%)和128 h(氧气体积分数为0%)达到峰值, 分别为16.00 nmol·(g·h)-1和13.62 nmol·(g·h)-1.N2产生速率在152 h时达到峰值, 为26.50 nmol·(g·h)-1.
当初始氧气体积分数为3%时, 培养开始后96 h NO2-含量接近峰值, 为25.10 mg·kg-1, 该过程中NH4+的减少量等于NO2-的增加量; 96 h(氧气体积分数为1.16%)至176 h(氧气体积分数为0%)的培养过程中, NO2-、NO3-含量无显著变化, 其中NO2-最高达到27.11 mg·kg-1, NH4+含量从5.42 mg·kg-1降至1.83 mg·kg-1; 176 h之后NO2-、NO3-含量呈降低趋势, NH4+则相反.N2O产生速率分别于108 h时(氧气体积分数为0.078%)、176 h(氧气体积分数为0%)达到峰值77.91 nmol·(g·h)-1及7.08 nmol·(g·h)-1.NO、N2产生速率从96 h开始稳定增加, 至176 h时分别达到高峰值6.55 nmol·(g·h)-1及4.48 nmol·(g·h)-1, 随后下降.3%氧气体积分数下N2产生率峰值显著小于初始氧气体积分数为0%和1%的处理, 导致该氧水平下N2O/(N2O+NO+N2)指数显著增加(表 2).
|
|
表 2 不同初始氧气体积分数下N2O/(N2O+NO+N2)指数1) Table 2 N2O/(N2O+NO+N2) index under different initial volume fractions of oxygen |
当初始氧气体积分数为5%时, 96 h(氧气体积分数为1.76%)时, NH4+含量由0 h的22.10 mg·kg-1降至1.27 mg·kg-1, 此时NO2-增长至高峰值25.73mg·kg-1, 随后降低; 整个培养过程中NO3-从0 h的24.80 mg·kg-1增长至培养结束时的71.0 mg·kg-1.与5%相比, 10%初始氧气体积分数下氨氧化速率显著增加(P<0.05), 80 h时NH4+降至最低, 此时NO2-达到峰值26.61 mg·kg-1.
由图 1中5%和10%氧梯度下气体产生动态可见, 初始氧气体积分数高于5%后, N2O产生量显著降低(P<0.01).60 h时两个氧梯度下培养瓶顶空氧气体积分数分别降至2.50%和7.59%, 此时NO累积量和N2O产生速率同时出现峰值, 分别为4.69 nmol·g-1、2.61 nmol·g-1和1.59 nmol·(g·h)-1、0.38 nmol·(g·h)-1, 随后NO累积量和N2O产生速率降低.在5%初始氧气体积分数下, 从96 h开始至培养结束(氧气体积分数为0.099%), N2O产生速率始终维持在1.43~1.59 nmol·(g·h)-1, NO、N2O测量值及N2产生速率逐渐增加, 培养结束时分别达到5.79 nmol·g-1、311.14 nmol·g-1和2.83 nmol·(g·h)-1.尽管5%初始氧气体积分数下N2O产生量显著降低(P<0.01), 但对N2O/(N2O+NO+N2)指数无显著影响(表 2).
2.2 DI和DI+NaClO3处理在不同初始氧气体积分数下气体、无机氮的动态变化添加NaClO3的处理反应前后土壤pH没有发生显著变化.在初始氧气体积分数为0%和1%时, NaClO3的添加显著降低土壤中NO2-的含量.在初始氧气体积分数为5%和10%时, 添加NaClO3后土壤NO2-含量持续增加, 培养结束时NO2-峰值与不添加NaClO3处理无显著差异; 两种氧气体积分数下NO3-增量为不添加NaClO3处理的53%左右, NH4+含量从培养初期的24.70 mg·kg-1缓慢降至培养末期的5.78 mg·kg-1(图 2).

|
图 2 DI土壤不添加和添加NaClO3处理无机氮的动态变化 Fig. 2 Dynamic changes in mineral nitrogen content of DI soil with or without sodium chlorate (NaClO3) addition |
在0%和1%初始氧气体积分数下, NaClO3的添加促使NO高峰期分别提前了16 h和8 h, 且高峰值仅分别为不添加NaClO3处理的20.0%和32.8%;添加NaClO3的处理N2O出现高峰的时间分别提前了32 h和24 h, 且N2O产生量峰值降低.与NO和N2O不同的是, 添加NaClO3并未影响土壤N2的产生量.与未添加NaClO3的处理相比, 在5%初始氧气体积分数下, NO、N2O与N2这3种气体的产生量峰值分别降低了53.7%、59.7%、55.9%, 在10%初始氧气体积分数下分别降低了35.4%、21.7%、38.7%(图 3).

|
图 3 DI土壤不添加/添加NaClO3处理NO、N2O、N2的动态变化 Fig. 3 Dynamic changes in NO, N2O, and N2 of DI soil with or without NaClO3 addition |
质量平衡计算结果表明, 各处理均存在氮的净矿化, 其中DIS土壤的净矿化量在5.9~17.8 mg·kg-1之间, 显著低于DI土样的净矿化量(P<0.01), 后者矿化量介于12.2~20.5 mg·kg-1之间.比较两种土壤在不同氧气体积分数下的N2O(图 1和图 3)产生峰值和CO2(表 3)累积产生量可以看出, 当初始氧气体积分数≤1%时, DIS土壤N2O和CO2产生量显著高于DI土壤(P<0.01).在5%初始氧气体积分数下, 两种土样N2O和CO2产生量无显著差异.初始氧气体积分数为10%时, 培养结束时DIS和DI土壤中氧气体积分数分别为4.09%和2.14%, DI土壤N2O和CO2产生量显著高于DIS土壤(P<0.01).
|
|
表 3 不同初始氧气体积分数下DIS和DI土壤CO2累积产生量/nmol·g-1 Table 3 Cumulative CO2 content in the soil with or without straw addition under different initial volume fractions of oxygen/nmol·g-1 |
3 讨论
在高氮投入的设施菜田, 碳氮比会严重失调[4], 高碳氮比玉米秸秆的投入可能会对土壤氮素的转化产生一定的影响[22].本研究中, DI土壤的矿化量普遍高于DIS, 尤其是在0%和1%初始氧气体积分数下, 这主要受土壤C/N比的影响, 低C/N比的DI土壤更易发生矿化作用[27].长期添加作物秸秆的土壤有高的N2O产生潜势[22, 23], 当氧气体积分数≤1%时, 这一现象尤为显著.在10%初始氧气体积分数下, DI土壤N2O/(NO+N2O+N2)指数和CO2产生量显著高于DIS, 这是由于DI土壤较高的矿化作用和本底NH4+含量显著增加了其氧气消耗速度, 较低的土壤氧含量会增加土壤微生物活性, 促进土壤CO2和N2O的产生[28].
氧气会通过改变土壤功能基因的表达、酶活性等直接影响土壤氮素转化和N2O的产生[15, 29].本研究中, 氧耗竭时会观察到N2O产生速率峰值, 但N2产生速率峰值随着氧气体积分数的升高显著降低, 导致1%和3%初始氧气体积分数下N2O/(NO+N2O+N2)指数显著增加.这可能由于与其它反硝化酶相比, N2O还原酶N2OR的活性表达时间相对滞后, 且其活性对氧气更敏感[30, 31].同时, 考虑到供试土壤的采样时间在收获期后, 土壤有效性碳缺乏会导致电子供体不足, 也可能是导致N2O不能被有效还原的重要因素[32].
氧气会通过影响中间产物(NO2-、NO等)的累积量, 间接地影响NOx的产生.Nadeem等[33, 34]的研究认为, 有氧条件下NO会刺激反硝化调节蛋白NNR(对O2及NO敏感)的活性, 进而诱导nirS、nosZ基因的表达, 发生有氧条件下的异养反硝化反应.本研究虽未对相关功能基因进行定量分析, 但NO的显著增加或者产生速率高峰总是伴随着氧耗竭现象的发生, 且初始氧气体积分数高于5%后NO测量值显著降低, 表明本研究中有氧条件下异养反硝化途径对N2O的贡献量较少[35].同时也说明, 本研究中NO产生量的大小及产生速率高峰出现的时间可以作为判断N2O产生途径的依据.
土壤中NO2-的过渡性累积会导致N2O排放量或N2O/(NO+N2O+N2)指数显著增加[20, 36].Dandie等[37]认为NO3-、NO2-的还原与N2O还原相比存在竞争优势, 导致土壤中N2O的大量产生.此外, 大量研究认为, 出于解毒机制, NO2-会诱导部分氨氧化细菌发生硝化细菌的反硝化反应, 终产物以N2O的形式排放[20, 27, 38].在1%和3%初始氧气体积分数下, 从有氧向厌氧过渡过程中, 土壤NO2-持续累积且先于NO出现第一个N2O产生速率高峰.同时, 添加NaClO3后, NH4+氧化的产物全部转化为NO2-, 5%、10%初始氧气体积分数下土壤中持续增加的NO2-与N2O产生量两者之间线性相关(图 4).由此可见, NO2-的持续累积对N2O的产生有诱导效应, 且土壤NO2-累积量的高低可能主要受土壤初始底物浓度的影响.5%初始氧气体积分数下NO、N2O测量值显著降低, 但N2O/(NO+N2O+N2)指数显著高于0%以及10%初始氧梯度的处理, 表明尽管有氧条件下土壤N2O的产生量显著低于厌氧条件, 但NO2-诱导的硝化细菌反硝化过程会显著增加土壤的N2O/(NO+N2O+N2)指数.Rassamee等[11]的研究结果显示氧气体积分数高于6.5 mg·L-1(12%氧气体积分数)时, 即使NO2-存在, 只会产生很少的N2O, 这是由于氧高于6.83%后NO2-会被迅速氧化为NO3-[39], 这一观点可以由未添加NaClO3处理的DIS土壤在10%初始氧气体积分数下无机氮、气体变化情况得以验证.

|
图 4 有氧条件下添加/不添加NaClO3处理NO2-与N2O的关系 Fig. 4 Relationship between NO2- and N2O with or without NaClO3 addition under aerobic conditions |
NaClO3的添加没有出现N2O产生量显著高于未添加处理的现象, 这与Schneider等[19]的研究结果不一致, 其认为有氧条件下氨氧化过程(NH4+氧化为NO2-)N2O的排放因子是亚硝酸氧化过程(NH4+氧化为NO3-)的4倍.这是由于本试验中虽然CO2产生量不受NaClO3添加的影响, 但NaClO3不仅抑制了NO2-的氧化, 也同时对NH4+氧化过程产生了抑制效应[21].与大部分氨氧化抑制剂作用机理相同, NaClO3会通过降低NH4+氧化速率减少NO2-的积累以及N2O的产生[20, 36, 40], 造成添加NaClO3处理的DI土壤N2O产生量显著低于不添加处理, 这也同时证实了硝化细菌反硝化过程对N2O排放的贡献.厌氧条件下添加NaClO3后所有中间产物(NO2-、NO、N2O)累积量降低, 但N2累积量无差异, 这可能是由于NaClO3的添加刺激了ROC (rapid, complete onset)类菌株的活性, 此类菌被认为可以快速、完全地表达所有的反硝化功能基因, 而基本不会出现中间产物的过渡积累[16, 17].
4 结论(1) 有氧条件下土壤N2O的产生量显著低于厌氧条件, 且有氧条件下异养反硝化途径对N2O的贡献量较少.长期添加作物秸秆的土壤有高的N2O产生潜势, 当氧气体积分数≤1%时, 这一现象尤为显著.
(2) 低氧条件下(初始氧气体积分数介于1%~5%时)N2O/(NO+N2O+N2)指数显著高于厌氧以及好氧(O2≥10%)处理.不完全的反硝化和NO2-诱导的硝化细菌的反硝化共同作用, 显著增加了低氧条件下土壤N2O的产生量和N2O/(N2O+NO+N2)指数.
(3) 在田间实际条件下, 通过优化施肥保证作物正常的水肥需求的同时, 避免饱和灌溉以维持土壤较高的孔隙含氧量, 对降低土壤N2O产生量、N2O/(NO+N2O+N2)指数有重要意义.
| [1] | Reay D S, Davidson E A, Smith K A, et al. Global agriculture and nitrous oxide emissions[J]. Nature Climate Change, 2012, 2(6): 410-416. DOI:10.1038/nclimate1458 |
| [2] | Liu Q H, Qin Y M, Zou J W, et al. Annual nitrous oxide emissions from open-air and greenhouse vegetable cropping systems in China[J]. Plant and Soil, 2013, 370(1-2): 223-233. DOI:10.1007/s11104-013-1622-3 |
| [3] | Zheng X H, Han S H, Huang Y, et al. Re-quantifying the emission factors based on field measurements and estimating the direct N2O emission from Chinese croplands[J]. Global Biogeochemical Cycles, 2004, 18(2): GB2018. |
| [4] |
王敬国. 设施菜田退化土壤修复与资源高效利用[M]. 北京: 中国农业大学出版社, 2011. Wang J G. Management of degraded vegetable soils in greenhouses[M]. Beijing: China Agricultural University Press, 2011. |
| [5] | Pijuan M, Torà J, Rodríguez-Caballero A, et al. Effect of process parameters and operational mode on nitrous oxide emissions from a nitritation reactor treating reject wastewater[J]. Water Research, 2014, 49: 23-33. DOI:10.1016/j.watres.2013.11.009 |
| [6] | Pan M, Wen X G, Wu G X, et al. Characteristics of nitrous oxide (N2O) emission from intermittently aerated sequencing batch reactors (IASBRs) treating slaughterhouse wastewater at low temperature[J]. Biochemical Engineering Journal, 2014, 86: 62-68. DOI:10.1016/j.bej.2014.03.003 |
| [7] | Wunderlin P, Mohn J, Joss A, et al. Mechanisms of N2O production in biological wastewater treatment under nitrifying and denitrifying conditions[J]. Water Research, 2012, 46(4): 1027-1037. DOI:10.1016/j.watres.2011.11.080 |
| [8] | Hu H W, Chen D L, He J Z. Microbial regulation of terrestrial nitrous oxide formation:understanding the biological pathways for prediction of emission rates[J]. FEMS Microbiology Reviews, 2015, 39(5): 729-749. DOI:10.1093/femsre/fuv021 |
| [9] | Baggs E M. A review of stable isotope techniques for N2O source partitioning in soils:recent progress, remaining challenges and future considerations[J]. Rapid Communications in Mass Spectrometry, 2008, 22(11): 1664-1672. DOI:10.1002/(ISSN)1097-0231 |
| [10] | Zhu X, Burger M, Doane T A, et al. Ammonia oxidation pathways and nitrifier denitrification are significant sources of N2O and NO under low oxygen availability[J]. Proceedings of the National Academy of Sciences of the United States of America, 2013, 110(16): 6328-6333. DOI:10.1073/pnas.1219993110 |
| [11] | Rassamee V, Sattayatewa C, Pagilla K, et al. Effect of oxic and anoxic conditions on nitrous oxide emissions from nitrification and denitrification processes[J]. Biotechnology and Bioengineering, 2011, 108(9): 2036-2045. DOI:10.1002/bit.v108.9 |
| [12] | Frame C H, Casciotti K L. Biogeochemical controls and isotopic signatures of nitrous oxide production by a marine ammonia-oxidizing bacterium[J]. Biogeosciences, 2010, 7(2): 2695-2709. |
| [13] | Morley N, Baggs E M. Carbon and oxygen controls on N2O and N2 production during nitrate reduction[J]. Soil Biology and Biochemistry, 2010, 42(10): 1864-1871. DOI:10.1016/j.soilbio.2010.07.008 |
| [14] | Miyahara M, Kim S W, Fushinobu S, et al. Potential of aerobic denitrification by Pseudomonas stutzeri TR2 to reduce nitrous oxide emissions from wastewater treatment Plants[J]. Applied and Environmental Microbiology, 2010, 76(14): 4619-4625. DOI:10.1128/AEM.01983-09 |
| [15] | Figueroa-González I, Quijano G, Laguna I, et al. A fundamental study on biological removal of N2O in the presence of oxygen[J]. Chemosphere, 2016, 158: 9-16. DOI:10.1016/j.chemosphere.2016.05.046 |
| [16] | Bergaust L, Mao Y J, Bakken L R, et al. Denitrification response patterns during the transition to anoxic respiration and posttranscriptional effects of suboptimal pH on nitrogen oxide reductase in Paracoccus denitrificans[J]. Applied and Environmental Microbiology, 2010, 76(19): 6387-6396. DOI:10.1128/AEM.00608-10 |
| [17] | Liu B B, Frostegård Å, Shapleigh J P. Draft genome sequences of five strains in the genus Thauera[J]. Genome Announcements, 2013, 1(1): e00052-12. |
| [18] | Liu B B, Frostegård Å, Bakken L R. Impaired reduction of N2O to N2 in acid soils is due to a posttranscriptional interference with the expression of nosZ[J]. mBio, 2014, 5(3): 01383-14. |
| [19] | Schneider Y, Beier M, Rosenwinkel K H. Influence of operating conditions on nitrous oxide formation during nitritation and nitrification[J]. Environmental Science and Pollution Research, 2014, 21(20): 12099-12108. DOI:10.1007/s11356-014-3148-5 |
| [20] | Ma L, Shan J, Yan X Y. Nitrite behavior accounts for the nitrous oxide peaks following fertilization in a fluvo-aquic soil[J]. Biology and Fertility of Soils, 2015, 51(5): 563-572. DOI:10.1007/s00374-015-1001-8 |
| [21] | Bauhus J, Meyer A C, Brumme R. Effect of the inhibitors nitrapyrin and sodium chlorate on nitrification and N2O formation in an acid forest soil[J]. Biology and Fertility of Soils, 1995, 22(4): 318-325. |
| [22] |
宋贺, 王敬国, 陈清, 等. 设施菜田不同碳氮管理对反硝化菌结构和功能的影响[J]. 微生物学通报, 2014, 41(11): 2283-2292. Song H, Wang J G, Chen Q, et al. Impacts of different carbon and nitrogen managements on the structure and function of denitrifier community in a greenhouse vegetable cropping system[J]. Microbiology China, 2014, 41(11): 2283-2292. |
| [23] | Gao J M, Xie Y X, Jin H Y, et al. Nitrous oxide emission and denitrifier abundance in two agricultural soils amended with crop residues and urea in the North China plain[J]. PLoS One, 2016, 11(5): e0154773. DOI:10.1371/journal.pone.0154773 |
| [24] | Fan Z B, Lin S, Zhang X M, et al. Conventional flooding irrigation causes an overuse of nitrogen fertilizer and low nitrogen use efficiency in intensively used solar greenhouse vegetable production[J]. Agricultural Water Management, 2014, 144: 11-19. DOI:10.1016/j.agwat.2014.05.010 |
| [25] | Molstad L, Dörsch P, Bakken L R. Robotized incubation system for monitoring gases (O2, NO, N2O N2) in denitrifying cultures[J]. Journal of Microbiological Methods, 2007, 71(3): 202-211. DOI:10.1016/j.mimet.2007.08.011 |
| [26] | Belser L W, Mays E L. Specific inhibition of nitrite oxidation by chlorate and its use in assessing nitrification in soils and sediments[J]. Applied and Environmental Microbiology, 1980, 39(3): 505-510. |
| [27] | Eberwein J R, Oikawa P Y, Allsman L A, et al. Carbon availability regulates soil respiration response to nitrogen and temperature[J]. Soil Biology and Biochemistry, 2015, 88: 158-164. DOI:10.1016/j.soilbio.2015.05.014 |
| [28] | Banerjee S, Helgason B, Wang L F, et al. Legacy effects of soil moisture on microbial community structure and N2O emissions[J]. Soil Biology and Biochemistry, 2016, 95: 40-50. DOI:10.1016/j.soilbio.2015.12.004 |
| [29] | Bueno E, Mesa S, Bedmar E J, et al. Bacterial adaptation of respiration from oxic to microoxic and anoxic conditions:redox control[J]. Antioxidants & Redox Signaling, 2012, 16(8): 819-852. |
| [30] | Zheng J Q, Doskey P V. Simulated rainfall on agricultural soil reveals enzymatic regulation of short-term nitrous oxide profiles in soil gas and emissions from the surface[J]. Biogeochemistry, 2016, 128(3): 327-338. DOI:10.1007/s10533-016-0210-z |
| [31] | Qu Z, Bakken L R, Molstad L, et al. Transcriptional and metabolic regulation of denitrification in Paracoccus denitrificans allows low but significant activity of nitrous oxide reductase under oxic conditions[J]. Environmental Microbiology, 2016, 18(9): 2951-2963. DOI:10.1111/1462-2920.13128 |
| [32] | Chen Y G, Wang D B, Zheng X, et al. Biological nutrient removal with low nitrous oxide generation by cancelling the anaerobic phase and extending the idle phase in a sequencing batch reactor[J]. Chemosphere, 2014, 109: 56-63. DOI:10.1016/j.chemosphere.2014.02.011 |
| [33] | Nadeem S, Dörsch P, Bakken L R. Autoxidation and acetylene-accelerated oxidation of NO in a 2-phase system:implications for the expression of denitrification in ex situ experiments[J]. Soil Biology and Biochemistry, 2013, 57: 606-614. DOI:10.1016/j.soilbio.2012.10.007 |
| [34] | Nadeem S, Dörsch P, Bakken L R. The significance of early accumulation of nanomolar concentrations of NO as an inducer of denitrification[J]. FEMS Microbiology Ecology, 2013, 83(3): 672-684. DOI:10.1111/fem.2013.83.issue-3 |
| [35] | Guo G X, Deng H, Qiao M, et al. Effect of long-term wastewater irrigation on potential denitrification and denitrifying communities in soils at the watershed scale[J]. Environmental Science & Technology, 2013, 47(7): 3105-3113. |
| [36] | Maharjan B, Venterea R T. Nitrite intensity explains N management effects on N2O emissions in maize[J]. Soil Biology and Biochemistry, 2013, 66: 229-238. DOI:10.1016/j.soilbio.2013.07.015 |
| [37] | Dandie C E, Wertz S, Leclair C L, et al. Abundance, diversity and functional gene expression of denitrifier communities in adjacent riparian and agricultural zones[J]. FEMS Microbiology Ecology, 2011, 77(1): 69-82. DOI:10.1111/fem.2011.77.issue-1 |
| [38] | Yu R, Kampschreur M J, Van Loosdrecht M C, et al. Mechanisms and specific directionality of autotrophic nitrous oxide and nitric oxide generation during transient anoxia[J]. Environmental Science & Technology, 2010, 44(4): 1313-1319. |
| [39] | Peng L, Ni B J, Ye L, et al. The combined effect of dissolved oxygen and nitrite on N2O production by ammonia oxidizing bacteria in an enriched nitrifying sludge[J]. Water Research, 2015, 73: 29-36. DOI:10.1016/j.watres.2015.01.021 |
| [40] | Abed R M M, Lam P, De Beer D, et al. High rates of denitrification and nitrous oxide emission in arid biological soil crusts from the Sultanate of Oman[J]. The ISME Journal, 2013, 7(9): 1862-1875. DOI:10.1038/ismej.2013.55 |