 2018, Vol. 39
2018, Vol. 39



2. 中国科学院南京地理与湖泊研究所, 湖泊与环境国家重点实验室, 南京 210008
2. State Key Laboratory of Lake Science and Environment, Nanjing Institute of Geography and Limnology, Chinese Academy of Sciences, Nanjing 210008, China
天然有机质(organic matter, OM)广泛存在于各类水体, 含有丰富的碳、氮、磷等生源要素, 其形成、转化、归宿及其相伴随的营养盐再生过程是水生态系统营养盐生物地球化学循环的重要环节, 在海洋及内陆水体的生态学研究中受到高度重视[1, 2], 在湖泊蓝藻水华暴发过程中势必也扮演着非常重要的营养盐供给者的角色.
水体中的OM一般可分为两个主要的库——溶解态有机质(dissolved organic matter, DOM)和颗粒态有机质(particulate organic matter, POM), 通常将能通过0.45 μm或0.75 μm玻璃纤维滤膜的有机质定义为溶解态, 其余的为颗粒态[3~6].光化学降解、酶解以及细菌降解等这些生物和非生物因素参与的降解转化过程都会使OM的存在形态、组分以及化学特征发生改变, 将POM转化为更具生物可利用性的溶解态有机质以及无机营养盐, 进而参与到湖泊生源要素循环过程中[7, 8].因此, 深入认识水体中POM与DOM的组分及其来源差异, 很可能成为揭示POM-DOM之间转化机制的一个关键环节[9].然而, 国内外关于OM组分及其来源等方面的研究主要集中在DOM方面[10~13], 而对POM组分及其来源等方面的研究尚处于起步阶段;究其原因主要是POM结构复杂及技术手段的限制[14, 15].
Osburn等[9]在前人的基础上运用三维荧光光谱技术(three-dimensional excitation-emission matrix fluorescence spectra, EEMs)结合平行因子分析法(parallel factor analysis, PARAFAC), 总结了POM-DOM PARAFAC三维荧光模型, 对北卡罗莱纳州Neuse河口POM和DOM两种形态的荧光组分进行了解译, 并对POM降解转化过程中新生DOM以及剩余POM的含量及组分变化进行了定性定量的分析, 为研究湖泊水体中OM特征及其来源提供了新的可能. Brym等[16]结合POM-DOM PARAFAC模型分析了在近海沿岸海洋环境中提取的颗粒态有机质的光学和化学特征. Osburn等[17]利用POM-DOM PARAFAC模型揭示了在盐沼泽地及其邻近河口之间交换的溶解态和颗粒态有机质的含量的季节性变化. Larsen等[18]运用POM-DOM PARAFAC模型对恢复和未恢复城市河流中有机沉积物的来源进行了示踪.然而, 目前为止在中国内陆湖泊, POM与DOM荧光组分及其来源差异等方面的研究程度相对较低, 有必要进行系统深入地研究.
湖泊富营养化会导致水体生产力提高, 直接后果是水体内源产生的OM剧烈增加, 而且其来源、组成及循环耦合过程(如:速率、通量、主要过程和机制等)也相应发生着明显的变化[19].例如海洋及河口中POM有机碳所占比重一般约为干重的10%~40%, 而在一些淡水生态系统中, 尤其是水华暴发期, 浮游动植物以及水生植物的残体是构成颗粒态有机质的基本成分, 使得这一比例甚至可达66%[20].微囊藻等藻类由于伪空泡的作用漂浮在水面上, 并且由于风力作用以及岸边大型水生植物如芦苇等的拦截作用而大量堆积在岸边, 这些都使得藻源POM接受太阳辐射的机会大大增多[21].而且相比其他POM来源, 藻源POM降解后可快速释放出大量生物活性很高的DOM和无机营养盐, 供水体中的藻类再次利用, 可能对水华的暴发程度和周期产生重要的影响[20].这暗示着, 富营养化浅水湖泊中POM尤其是藻源等高生物可利用性POM的降解转化过程和水华暴发之间可能存在着关联.
孔繁翔等指出[22], 蓝藻水华的形成可分为休眠、复苏、生长和上浮聚集这4个阶段, 其中冬季蓝藻进入休眠阶段, 春季蓝藻开始复苏, 夏季大量生长, 秋季上浮聚集.目前大部分研究都集中在夏季蓝藻暴发期, 笔者也调查了夏季颗粒态有机质的荧光特征[23], 但是对冬、春蓝藻休眠及复苏季节的相关研究较少.本研究在富营养化浅水湖泊太湖运用POM-DOM PARAFAC模型, 重点探讨了春季蓝藻复苏和冬季POM与DOM的荧光特征并对其可能来源进行分析, 以期为深刻认识富营养化浅水湖泊中溶解态和颗粒态有机质的生物地球化学循环、和其与蓝藻水华暴发之间的因果关系提供科学依据.
1 材料与方法 1.1 样品采集与预处理于2015年2月26日和5月28日分别在梅梁湾(1~6号、17号)、竺山湾(12号、13号)、贡湖湾(7号、10号、11号、16号)、河口区(9号、14号)和湖心区(8号、15号)进行了采样, 分别采集17个样品(图 1), 采样点位同中国科学院太湖生态系统研究站的常规监测点位(编号不同), 南、北部湖区参照Zhou等[24]的方法进行分区, 北部湖区夏季易暴发蓝藻水华即藻型湖区, 南部湖区为沉水植物覆盖即草型湖区.采集5 L水样并置于保温箱中用冰块保存确保低温, 所有采样瓶事先均用10% HCl溶液浸泡、去离子水清洗及现场水润洗, 并于采样结束后直接送至中国科学院南京地理与湖泊研究所实验室放入冰箱内冷藏、冷冻保存.水体温度、水深、浊度等物理指标均在采样现场测定. POC与DOC浓度(mg·L-1)及POM与DOM三维荧光光谱分析均在采样结束后2~3 d内分析完成.

|
图 1 太湖水体采样点分布示意 Fig. 1 Distribution of the sampling sites in Lake Taihu |
共采集5 L水样, 根据水的浑浊程度取100~1 000 mL水样过滤到事先经450℃灼烧并称重的0.7 μm的GF/F滤膜上, 滤后水再用0.2 μm的微孔膜过滤测定DOM的三维荧光;滤膜上的POM荧光物质用10 mL 0.1 mol·L-1的NaOH黑暗条件下振荡24 h, 用HCl中和后调至pH值>6, 经0.2 μm的微孔膜过滤后的滤液用于测定POM的三维荧光, 测定方法参照Osburn等[9]的方法进行.
三维荧光光谱采用日立F-7000 FL分光光度计测定, 激发和发射夹缝宽度为5 nm, 激发波长为200~450 nm, 间隔为5 nm, 发射波长250~600 nm, 以1 nm间隔得到荧光光谱, 减去加酸中和过后的NaOH三维荧光光谱以校正水的拉曼散射, 同时利用0.01 mg·L-1硫酸奎宁进行荧光定标, 并用吸光度校正三维荧光光谱的内滤效应[25].
1.2.2 三维荧光光谱的平行因子分析法(PARAFAC)及荧光光谱参数EEM是通过波谱形状或波峰数量进行组分个数和类别的假定, 而PARAFAC法是通过数学统计的方法把OM复杂的荧光数据矩阵分离出不同的组分, 将所有点位的整个三维荧光数据组分解成3个线性项和1个残留数组, 它是基于三线性理论的一种方法[26]. PARAFAC法计算过程可以称为交替最小二乘(ALS)算法:
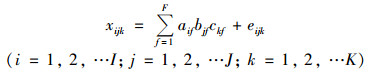
|
式中, Xijk是第i个样点再激发波长为k、发射波长为j处的荧光强度;aif是第i个样点的第f次分析的含量;bjf和ckf分别是与第f次分析后激发波长为j、发射波长为k光谱的线性相关. eijk为系统残差, 是不受模型控制的变量. F为模型中选择的组分因子数量.
本研究在MATLAB中使用DOMFluor工具箱运行PARAFAC法模型对样点的三维荧光数据进行分析.
荧光指数(FI、BIX、HIX)通常用来研究和表征物质的来源及不同来源对OM荧光强度的相对贡献率.荧光指数(fluorescence index, FI)定义为激发波长为370 nm时, 发射波长在450 nm与500 nm处的荧光强度比值, 这个比值反映了芳香氨基酸与非芳香物对荧光强度的相对贡献率, 因而可以作为物质的来源以及OM降解程度的指示指标[25], 陆源和生物来源的f450/500值分别为1.4和1.9[27].
生物源指数(autochthonous index, BIX)是310 nm激发波长下380 nm发射波长处荧光强度与420~435 nm区间最大荧光强度的比值, 可以作为OM溯源的一个指标, BIX值越高, 说明新近自生源组分的比例越高, BIX值越低表示主要来自陆源输入.有研究认为[28], BIX在0.6~0.7之间时, 具有较少的自生成分;BIX在0.7~0.8时具有中度新近自生源特征;在0.8~1.0之间时, 具有较强自生源特征;大于1.0是为生物细菌活动产生.
腐殖化指数(humification index, HIX)用来表征有机质腐殖化的程度或成熟度, 可以通过激发波长为254 nm时, 发射波长从435~480 nm的峰值面积比300~345 nm的荧光峰值面积计算, HIX值越高, 表明腐殖化程度越高.有研究认为[29], 当HIX小于4表示由生物活动产生, 腐殖化程度较弱, 以自生源为主;当HIX值在4~6之间时, 属于较弱腐殖质特征和较强自生源特征;HIX值大于6, 属于强腐殖质特征, 陆源贡献较大.
1.2.3 其他水质参数的测定经高温(450℃)灼烧后的0.7 μm GF/F滤膜过滤水样, 过滤后的水经1020型TOC仪测定DOC浓度;留在滤膜上的颗粒物质经过冷冻干燥后称重, 减去原始滤膜重量, 即为POM的重量;将冻干后的滤膜用研钵磨碎后送至EA 3000的元素分析仪测定POC浓度(mg·L-1).叶绿素a(Chla)、化学需氧量(COD)、总氮(TN)、总磷(TP)等水质参数数据均来源于中国科学院太湖生态系统研究站, 测定方法参照文献[30].
1.3 绘图及数据处理采用ArcGIS 10.2软件绘制参数空间分布图与采样点分布, 均值、标准差以及相关性等采用SPSS 21.0软件分析.相关分析用来检测变量之间的关系;P>0.05表示未达到显著检验水平, 0.01<P<0.05为显著水平, P<0.01为极显著水平.
2 结果与分析 2.1 太湖水体各水质参数的时空分布特征天然有机质降解过程中大量耗氧, 同时释放出N、P、C等有机和无机营养盐, 进而影响各种养分的吸附/解析、相互转化及生物利用等过程[31]. TN、TP、Chla、COD、POC和DOC等水质参数的浓度和时空分布分别见表 1和图 2.春、冬季节TP、Chla、COD和POC具有类似的季节变化规律, 春季浓度及其变幅均大于冬季(t检验, P<0.01);而TN冬季的变幅及其浓度均大于春季(t检验, P<0.01), Xu等[32]也获得了类似的结论;春、冬季节DOC浓度及其变幅无显著差异(t检验, P>0.05).各水质参数的空间分布模式不尽相同, TN、TP、COD和DOC的高值区均分布在竺山湾、梅梁湾及河口区, 呈现出近岸高, 远岸低的分布趋势, 这与Yao等[33]的研究结果一致;而Chla和POC春季高值区集中在梅梁湾区域, 冬季高值区分布在湖心区.
|
|
表 1 太湖各水质参数浓度/mg·L-1 Table 1 Concentrations of water quality parameters in Lake Taihu/mg·L-1 |

|
图 2 太湖水体各水质参数的时空分布 Fig. 2 Spatial-temporal distribution of water quality parameters in Lake Taihu |
荧光特征是描述有机质来源和组成的重要参数[34, 35].入湖河口区和湖心区典型站点POM和DOM的荧光特征见图 3.三维荧光图谱显示, POM和DOM共出现了6种类型的荧光峰(图 3), 包括1个可见光区的类腐殖质荧光C峰, 1个紫外光区的类腐殖质荧光A峰, 1个海源性类富里酸M峰, 以及3个类蛋白荧光B、T和D峰.一般认为C、A峰反映的是外源输入的腐殖酸和富里酸形成的荧光峰值, M峰代表的是微生物及水中藻类降解或人类活动产生的氧化的类腐殖质物质, 而B、T、D峰反映的是生物降解来源的色氨酸和酪氨酸形成的荧光峰值[36~41].但近年来也有部分研究显示浮游植物生长或降解过程中也会产生类腐殖质荧光[42, 43], 而来源于化工厂的城市污水中可能会含有类蛋白荧光[44].

|
6号采样点位于梅梁湾入湖河口区; 10号采样点位于大浦河河口区; 8号采样点位于湖心区 图 3 入湖河口区及湖心区典型站点三维荧光图谱 Fig. 3 Examples of EEM fluorescence spectra of OM from typical sampling sites at the river inflow and in the central part of Lake Taihu |
各个峰的位置及其峰值荧光强度如图 3所示. POM和DOM的三维荧光光谱中, 荧光峰、荧光中心位置和荧光强度均存在一定差异. POM的三维荧光光谱中出现了5种类型的荧光峰(T、D、M、A和C峰), 而在DOM的三维荧光光谱中发现了4种类型的荧光峰(B、D、A和C峰), 这说明POM具有更复杂的组分构成;He等[6]也获得了类似的结论.有研究表明, 对于同一种荧光峰, 其对应激发波长越大(红移), 所含芳香性越强, 分子量越大, 聚合度越高[45]; POM类酪氨酸荧光峰D峰激发波长为380 nm(DOM三维荧光光谱中D峰激发波长为350 nm), 出现明显的“红移”现象, 说明POM类酪氨酸荧光物质的结构和组成与DOM之间存在差异, 即POM类酪氨酸荧光物质的分子量、缩聚度和芳香度大于DOM. DOM的荧光强度要强于POM;典型站点的POM和DOM三维荧光光谱特征表明, 由河口区至湖心区的POM和DOM的荧光峰类型相似, 但荧光强度逐渐降低.从季节变化来看, POM春季荧光强度大于冬季, 而DOM冬季荧光强度大于春季.在DOM三维荧光光谱中, 类蛋白荧光强度(B和D峰)明显大于类腐殖质荧光强度(A和C峰).
2.3 太湖OM荧光组分的PARAFAC模型解析将2015年2月和5月采集的共34个POM和34个DOM三维荧光样品全部放入PARAFAC模型中进行计算, 一共得到5种荧光组分(C1~C5).太湖POM和DOM样品平行因子分析PARAFAC所得5个组分模型对所有EEMs数据信息的解释率可达99.6%以上, 并能很好地通过对半分析检验(图 4).这5种荧光组分的激发波长和发射波长见表 2.由图 4和表 2可知, 组分C1和C2均为类色氨酸物质, 对应的是传统意义上的T和B峰, 这类组分反映的是生物降解来源形成的荧光峰值, 主要由微生物和浮游植物的作用所产生, 也受到生活污水和工业废水等外源输入所携带的微生物影响[36, 37];组分C3反映的是海源性类富里酸M峰, 可能来源于微生物及水中藻类降解或人类活动产生的氧化的类腐殖质物质[38, 39];组分C4被认为是外源输入的腐殖酸和富里酸形成的荧光峰值, 对应的是A峰和C峰, 与类富里酸荧光和腐殖质结构中的羟基及羧基有关[40, 41];组分C5为生物降解来源的类酪氨酸D峰, 与芳环氨基酸结构有关[36, 40].

|
图 4 太湖POM和DOM各荧光组分 Fig. 4 Spectral shapes of the five components identified by PARAFAC |
|
|
表 2 POM和DOM的5种荧光组分的相对含量/% Table 2 Relative concentrations of the five component determined using the combined POM-DOM PARAFAC model/% |
POM和DOM荧光组分的相对含量见表 2. POM 5种荧光组分(C1p~C5p)的相对含量分别在3.44%~24.26%、0.00%~18.55%、15.26%~62.80%、1.21%~40.62%和7.68%~32.07%之间变化, 均值分别为14.38%±0.92%、7.39%±0.91%、43.53%±2.29%、15.68%±1.75%和19.02%±0.86%.春、冬季节太湖北部湖区C1p~C5p的分布存在明显的空间差异;春季C1p~C5p高值区均分布在梅梁湾区域, 而冬季C1p~C5p高值区均集中在湖心区.太湖北部湖区春、冬季节POM的季节差异并不明显, 均以荧光组分C3p为主.
DOM 5种荧光组分(C1d~C5d)的相对含量分别在16.61%~38.17%、27.80%~48.60%、5.59%~17.86%、2.70%~18.60%和0.00%~37.58%之间变化, 均值分别为30.64%±0.91%、39.69%±0.83%、9.72%±0.46%、5.04%±0.49%和14.91%±.1.43%.与POM不同, 春、冬季节太湖北部湖区C1d~C5d高值区均分布在竺山湾区域, 且均以类蛋白组分C2d、C1d和C5d为主.
2.4 各荧光组分之间及其与各水质参数的关系如表 3所示, 建立POM(C1p~C5p)和DOM(C1d~C5d)各荧光组分间的相关关系, 以分析各荧光组分间的关系.相关分析显示, 春季C1d与C2d、C3d与C4d之间有较强的相关性, 且显著正相关(P<0.01), 说明C1d与C2d这两种类色氨酸在组成及分布上相似, C3d与C4d这两种类腐殖质在组成及分布上亦相似, 周永强等[46]对洪湖、东湖与梁子湖CDOM组成特征分析中也获得了类似的结论.同理, C1p与C4p(P<0.05)、C2p与C5p(P<0.01)之间也具有较强的相关性, 说明C1p与C4p这两种物质在组成和来源上具有某种联系, C2p与C5p这两种类蛋白物质之间亦具有某种联系.值得注意的是, C3d(P<0.01)、C4d(P<0.05)与C3p均具有较好的相关关系, 表明POM与DOM在组分来源上具有某种联系;He等研究表明[6], POM与DOM之间通过生物因素或非生物因素易发生动态转化.与春季类似, 冬季荧光组分C1d与C2d、C3d与C4d、C1p与C2p、C1d与C2p亦具有极好的相关性(P<0.01).
|
|
表 3 POM和DOM的5种荧光组分的相对含量/% Table 3 Relative concentrations of the five component determined using the combined POM-DOM PARAFAC model/% |
春季C1d(P<0.01)和冬季C4d(P<0.01)、C5d(P<0.05)与TN之间存在相关关系, 春季C1d(P<0.01)、C3d(P<0.01)、C4d(P<0.01)、C3p(P<0.01)、C4p(P<0.01)和冬季C4d(P<0.01)与TP之间具有极显著相关关系, 表明OM与N、P元素的迁移转化过程密切相关.春季C2d(P<0.05)、C5d(P<0.05)、C3p(P<0.05)与COD均具有相关性, 证实了荧光强度的分布情况一定程度上可以反映水体中有机物的分布, 季乃云等[47]在研究大沽河到胶州湾的DOM荧光特征变化时也获得了类似的结论.春季C2p(P<0.01)、C3p(P<0.01)、C5p(P<0.05)与POC均具有较好的相关关系, 这表明类蛋白和类腐殖质是太湖POC的重要组成部分, 荧光强度可以作为一个反映水体中POC含量的有效参数, 这与方芳等[48]在三峡水库小江回水区溶解性有机物三维荧光光谱分析结果一致.春季C1d(P<0.01)、C3d(P<0.01)、C4d(P<0.01)、C1p(P<0.05)、C2p(P<0.05)、C3p(P<0.05)和冬季C3d(P<0.05)、C4d(P<0.05)与DOC均存在相关关系, 说明类蛋白和类腐殖质是太湖DOC的重要组成部分.
富营养化水体中存在大量的浮游植物, 浮游植物量的多少常以叶绿素a的浓度反映, 而浮游植物降解是产生荧光物质的重要来源之一[49], 故本研究进一步对叶绿素a与荧光组分的相关性进行了分析.分析结果显示, 春季C5d与Chla之间存在正相关关系(P<0.01), 而C2d(P<0.05)、C3p(P<0.01)与Chla之间具有负相关关系.冬季C4d与Chla之间也存在负相关关系(P<0.05).
3 讨论 3.1 POM和DOM可能来源途径从图 3三维荧光图谱可以发现, 河口区和湖心区荧光峰值出现的位置和强度的差异证明两种形态有机质中具有不同的荧光物质, 暗示河口区和湖心区OM的来源不一样.本研究中6号和10号采样点均位于河流的入湖口, 河流带来了大量的工业废水、生活污水以及土壤成分, 所以在河口区春、冬季节POM和DOM的来源主要为陆源, 类腐殖质荧光A峰和C峰比较强, 这与Baker等[44]对河流中DOM三维荧光研究结果类似, 河流中类腐殖质荧光显著大于类蛋白荧光. Baker等[44]在研究受污染河流DOM时还发现, 可以用类蛋白荧光来指示河流的受污染程度, 因为在洁净水体中类蛋白荧光峰很少出现, 在纯腐殖质样品的三维荧光图谱中亦没有类蛋白荧光峰.河口区6号和10号采样点由于受城市污水中类蛋白荧光的影响, POM在T、D峰的荧光峰值以及DOM在B、D峰的荧光峰值均较高.此外, 6号和10号采样点所在的河口区同时位于易暴发蓝藻水华的太湖西、北部, 再加上河口区氮、磷营养盐丰富, 生物活动剧烈, 生物降解的类蛋白应该也会有很大的贡献, 而并非全部来自河流输入.
8号采样点位于远离河口区的湖心区, 受河流影响减少, 由于流水的不断稀释, 外源输入的贡献有所降低, 从荧光峰值中也可以看出这一点, 河口区陆源类腐殖质荧光峰值明显强于湖心区, 王志刚等[50]在巢湖的研究得出了同样的观点.在湖心区, 受河流影响减弱, 类腐殖质荧光强度明显降低, 而类蛋白荧光强度却并没有等比例降低, 这表明在湖心区浮游植物的降解作用产生了新的类蛋白组分.根据傅平青等[51]对云南洱海的研究发现, 沉积物孔隙水中有较强的类蛋白荧光组分, 这是由于生物以及微生物残骸中蛋白质成分被分解后进入孔隙水中导致的. Coble等[35]对海洋沉积物孔隙水的研究中也发现了强的类蛋白荧光, 因此太湖北部湖区中类蛋白荧光也有可能是太湖沉积物孔隙水中类蛋白荧光组分的释放.
荧光指数(FI、BIX、HIX)通常用来研究和表征物质的来源及不同来源对OM荧光强度的相对贡献率.本研究中, 春季DOM荧光指数FI、BIX和HIX分别在1.83~2.43、0.91~1.26和0.26~1.96之间, 均值分别为1.97±0.03、1.16±0.02和0.50±0.10, 这表明春季DOM受内源贡献较大.冬季DOM荧光指数FI、BIX和HIX分别在1.65~2.50、1.00~1.27和0.23~0.93之间, 均值分别为2.02±0.05、1.19±0.02和0.43±0.04, 整体处于较强的自生源特征.春季POM荧光指数FI、BIX和HIX分别在1.66~2.80、0.30~1.55和1.40~4.81之间, 均值分别为2.26±0.08、1.00±0.09和2.92±0.25, 说明春季太湖北部湖区POM整体处于弱腐殖质特征, 且以自生源为主.冬季POM荧光指数FI、BIX和HIX分别在1.64~2.16、1.06~1.35和0.69~1.88之间, 均值分别为1.82±0.03、1.18±0.02和1.28±0.10, 这表明冬季POM受内源贡献较大.相比之下, 入湖河流的河口区(图 5中圈出部分), FI和BIX相对较小, HIX相对较大, 趋近陆源特征, 表明河口区受陆源输入影响相对显著一些.综上所述, 3种荧光指数对表征OM来源均具有良好的指示性, 都显示出春、冬季节太湖北部湖区水体POM和DOM均以自生源为主, 河口区受陆源贡献较大;江俊武等[52]认为太湖水体DOM主要为内源贡献, 河口区外源输入相对明显一些.曾庆飞等[53]使用稳定同位素方法也证实太湖水体中0.7~120 μm粒径间的POM主要为内源藻类贡献.

|
图 5 太湖POM和DOM样本的FI-HIX and BIX-HIX分布 Fig. 5 FI-HIX and BIX-HIX distributions of POM and DOM in Lake Taihu |
近年来, 有关浮游植物与类蛋白和类腐殖质荧光强度关系的说法不一.有报道指出, 高叶绿素a含量的水域对应着高类蛋白荧光强度[54~57], 高类腐殖质荧光强度对应着高叶绿素a含量[56~59].不过Mayer等[60]对Maine河口的研究表明, 高类蛋白荧光对应着高叶绿素a含量, 却也发现有时类蛋白荧光和叶绿素a呈负相关;任保卫等[61]通过研究胶州湾围隔实验中溶解有机物三维荧光特征时发现, 类酪氨酸荧光强度与叶绿素a浓度呈明显的负相关.但也有不少研究发现荧光组分与叶绿素a之间并没有显著的相关性, 如刘学利等[62]在研究东平湖溶解性有机质时发现, 各荧光组分与叶绿素a相关性均未达到显著水平;吕丽莎等[63]在研究东海溶解有机物时也发现, 荧光组分与叶绿素a之间无明显相关性.分析其原因, 可能与浮游植物所处的生长阶段有关;Petersen[64]在培养球等鞭金藻时发现, 衰老和死亡细胞的破碎降解是有机物类蛋白荧光的一个重要来源;Rochelle-Newall等[65]通过对中肋骨条藻和微小原甲藻等几种海洋微藻培养过程中有机物类腐殖质荧光变化的研究表明, 类腐殖质荧光在微藻进入稳定生长期后才开始逐渐增大, 其主要是来源于浮游植物释放的有机物质经由细菌进一步降解后的产物;这就表明, 某区域虽然具有相对高的浮游植物生物量, 如果此时正处于浮游植物的指数生长期, 类蛋白和类腐殖质荧光可能会低;相反, 如果正是浮游植物衰亡期, 叶绿素a含量不一定很高, 但可能会具有较高的类蛋白和类腐殖质荧光.本研究中, 如表 3所示, 冬季C4d与Chla之间存在负相关关系;春季C5d与Chla之间存在正相关关系, 而C2d、C3p与Chla之间具有负相关关系.
冬季, 浮游植物进入消亡时期, Chla含量降低[22], 类腐殖质荧光C4d与Chla存在负相关性(P<0.05), 这可能是因为浮游植物降解过程中产生了难降解的类腐殖质物质, 从而造成了类腐殖质荧光组分C4d随着Chla浓度的降低而升高的现象;Karmer等[42]的研究指出, 浮游植物原位降解产生的DOM中包含大约25%的类腐殖质荧光.
冬季蓝藻休眠到春季蓝藻复苏的过程中, 浮游植物量增加[22], Chla浓度升高(图 2), 荧光组分C2d、C3p的相对含量呈下降趋势(表 2), 相关分析显示C2d、C3p与Chla之间均存在负相关关系, C2d、C3p随着Chla浓度的增加而降低, 说明POM与DOM对蓝藻复苏起着重要的作用;冯胜等[66]的研究指出, 在微生物作用下的微囊藻降解实验进行至21 d时, 发现降解瓶上层颜色变绿, 伴随有部分新的浮游藻类产生. C5d相对含量升高(表 2), C5d与Chla具有正相关关系, C5d随着Chla浓度的增加而增加, 表明浮游植物是DOM类酪氨酸组分的重要来源, 与江俊武等[52]报道的类蛋白物质与Chla存在相关性相吻合.
4 结论(1) 春、冬季节太湖北部湖区POM和DOM的荧光峰、荧光中心位置和荧光强度均存在明显的差别;POM有着比DOM更复杂的组分构成.综合POM和DOM的荧光强度和荧光指数可知, 春、冬季节太湖北部湖区POM和DOM均以自生源为主, 河口区受陆源贡献较大.
(2) 通过POM-DOM PARAFAC三维荧光联合模型得出POM(C1p~C5p)和DOM(C1d~C5d)均含有5类荧光组分. POM和DOM的荧光组分特征存在明显的差别;春、冬季节POM均以类腐殖质荧光C3为主, 高值区分别集中在梅梁湾区域和湖心区;而DOM均以类蛋白组分C2、C1和C5为主, 高值区均分布在竺山湾区域.
(3) 春季C1d与C2d(P<0.01)、C3d与C4d(P<0.01)、C1p与C4p(P<0.05)、C2p与C5p(P<0.01)均具有较强的相关性.值得注意的是, C3d(P<0.01)、C4d(P<0.05)与C3p均具有较好的相关关系, 表明POM与DOM在组分来源上具有某种联系.与春季类似, 冬季荧光组分C1d与C2d、C3d与C4d、C1p与C2p、C1d与C2p亦具有极好的相关性(P<0.01).相关分析结果显示, 春季C5d与Chla之间存在正相关关系(P<0.01), 而C2d(P<0.05)、C3p(P<0.01)与Chla之间具有负相关关系;冬季C4d与Chla之间也存在负相关关系(P<0.05).
| [1] | Simon M, Grossart H P, Schweitzer B, et al. Microbial ecology of organic aggregates in aquatic ecosystems[J]. Aquatic Microbial Ecology, 2002, 28(2): 175-211. |
| [2] | Turner J T. Zooplankton fecal pellets, marine snow and sinking phytoplankton blooms[J]. Aquatic Microbial Ecology, 2002, 27: 57-102. DOI:10.3354/ame027057 |
| [3] | Verdugo P, Alldredge A L, Azam F, et al. The oceanic gel phase:a bridge in the DOM-POM continuum[J]. Marine Chemistry, 2004, 92(1-4): 67-85. DOI:10.1016/j.marchem.2004.06.017 |
| [4] | Roulet N, Moore T R. Environmental chemistry:Browning the waters[J]. Nature, 2006, 444(7117): 283-284. DOI:10.1038/444283a |
| [5] | Azam F, Malfatti F. Microbial structuring of marine ecosystems[J]. Nature Reviews Microbiology, 2007, 5(10): 782-791. DOI:10.1038/nrmicro1747 |
| [6] | He W, Chen M L, Schlautman M A, et al. Dynamic exchanges between DOM and POM pools in coastal and inland aquatic ecosystems:a review[J]. Science of the Total Environment, 2016, 551-552: 415-428. DOI:10.1016/j.scitotenv.2016.02.031 |
| [7] | McCallister S L, Bauer J E, Ducklow H W, et al. Sources of estuarine dissolved and particulate organic matter:a multi-tracer approach[J]. Organic Geochemistry, 2006, 37(4): 454-468. DOI:10.1016/j.orggeochem.2005.12.005 |
| [8] | Mayer L M, Schick L L, Hardy K R, et al. Photodissolution and other photochemical changes upon irradiation of algal detritus[J]. Limnology and Oceanography, 2009, 54(5): 1688-1698. DOI:10.4319/lo.2009.54.5.1688 |
| [9] | Osburn C L, Handsel L T, Mikan M P, et al. Fluorescence tracking of dissolved and particulate organic matter quality in a river-dominated estuary[J]. Environmental Science & Technology, 2012, 46(16): 8628-8636. |
| [10] | Lou T, Xie H X. Photochemical alteration of the molecular weight of dissolved organic matter[J]. Chemosphere, 2006, 65(11): 2333-2342. DOI:10.1016/j.chemosphere.2006.05.001 |
| [11] |
张运林, 秦伯强, 龚志军. 太湖有色可溶性有机物荧光的空间分布及其与吸收的关系[J]. 农业环境科学学报, 2006, 25(5): 1337-1342. Zhang Y L, Qin B Q, Gong Z J. Spatial distribution of chromophoric dissolved organic matter fluorescence and its relation with absorption in Lake Taihu[J]. Journal of Agro-Environment Science, 2006, 25(5): 1337-1342. |
| [12] | Gondar D, Thacker S A, Tipping E, et al. Functional variability of dissolved organic matter from the surface water of a productive lake[J]. Water Research, 2008, 42(1-2): 81-90. DOI:10.1016/j.watres.2007.07.006 |
| [13] | Zhang Y L, Van Dijk M A, Liu M L, et al. The contribution of phytoplankton degradation to chromophoric dissolved organic matter (CDOM) in eutrophic shallow lakes:Field and experimental evidence[J]. Water Research, 2009, 43(18): 4685-4697. DOI:10.1016/j.watres.2009.07.024 |
| [14] | Baker A, Elliott S, Lead J R. Effects of filtration and pH perturbation on freshwater organic matter fluorescence[J]. Chemosphere, 2007, 67(10): 2035-2043. DOI:10.1016/j.chemosphere.2006.11.024 |
| [15] | Santín C, Yamashita Y, Otero X L, et al. Characterizing humic substances from estuarine soils and sediments by excitation-emission matrix spectroscopy and parallel factor analysis[J]. Biogeochemistry, 2009, 96(1-3): 131-147. DOI:10.1007/s10533-009-9349-1 |
| [16] | Brym A, Paerl H W, Montgomery M T, et al. Optical and chemical characterization of base-extracted particulate organic matter in coastal marine environments[J]. Marine Chemistry, 2014, 162: 96-113. DOI:10.1016/j.marchem.2014.03.006 |
| [17] | Osburn C L, Mikan M P, Etheridge J R, et al. Seasonal variation in the quality of dissolved and particulate organic matter exchanged between a salt marsh and its adjacent estuary[J]. Journal of Geophysical Research:Biogeosciences, 2015, 120(7): 1430-1449. DOI:10.1002/2014JG002897 |
| [18] | Larsen L, Harvey J, Skalak K, et al. Fluorescence-based source tracking of organic sediment in restored and unrestored urban streams[J]. Limnology and Oceanography, 2015, 60(4): 1439-1461. DOI:10.1002/lno.10108 |
| [19] |
吴敬禄, 王苏民. 云南程海富营养化过程的碳氧稳定同位素示踪[J]. 第四纪研究, 2003, 23(5): 557-564. Wu J L, Wang S M. Stable isotopic tracing of historical progressive eutrophication from Lake Chenghai, Yunnan in China[J]. Quaternary Sciences, 2003, 23(5): 557-564. |
| [20] | Grossart H P. Ecological consequences of bacterioplankton lifestyles:changes in concepts are needed[J]. Environmental Microbiology Reports, 2010, 2(6): 706-714. DOI:10.1111/emi4.2010.2.issue-6 |
| [21] | Zhang Y L, van Dijk M A, Liu M L, et al. The contribution of phytoplankton degradation to chromophoric dissolved organic matter (CDOM) in eutrophic shallow lakes:field and experimental evidence[J]. Water Research, 2009, 43(18): 4685-4697. DOI:10.1016/j.watres.2009.07.024 |
| [22] |
孔繁翔, 马荣华, 高俊峰, 等. 太湖蓝藻水华的预防、预测和预警的理论与实践[J]. 湖泊科学, 2009, 21(3): 314-328. Kong F X, Ma R H, Gao J F, et al. The theory and practice of prevention, forecast and warning on cyanobacteria bloom in Lake Taihu[J]. Journal of Lake Sciences, 2009, 21(3): 314-328. DOI:10.18307/2009.0302 |
| [23] |
吕伟伟, 姚昕, 张保华, 等. 太湖颗粒态有机质的荧光特征及环境指示意义[J]. 环境科学, 2018, 39(5): 2056-2066. Lv W W, Yao X, Zhang B H, et al. Fluorescent characteristics and environmental significance of particulate organic matter in Lake Taihu, China[J]. Environmental Science, 2018, 39(5): 2056-2066. |
| [24] | Zhou Y Q, Zhang Y L, Shi K, et al. Lake Taihu, a large, shallow and eutrophic aquatic ecosystem in China serves as a sink for chromophoric dissolved organic matter[J]. Journal of Great Lakes Research, 2015, 41(2): 597-606. DOI:10.1016/j.jglr.2015.03.027 |
| [25] | McKnight D M, Boyer E W, Westerhoff P K, et al. Spectrofluorometric characterization of dissolved organic matter for indication of precursor organic material and aromaticity[J]. Limnology and Oceanography, 2001, 46(1): 38-48. DOI:10.4319/lo.2001.46.1.0038 |
| [26] | Yao X, Zhu G W, Cai L L, et al. Geochemical characteristics of amino acids in sediments of Lake Taihu, a large, shallow, eutrophic freshwater lake of china[J]. Aquatic Geochemistry, 2012, 18(3): 263-280. DOI:10.1007/s10498-012-9160-9 |
| [27] | Jaffé R, Boyer J N, Lu X, et al. Source characterization of dissolved organic matter in a subtropical mangrove-dominated estuary by fluorescence analysis[J]. Marine Chemistry, 2004, 84(3-4): 195-210. DOI:10.1016/j.marchem.2003.08.001 |
| [28] | Huguet A, Vacher L, Relexans S, et al. Properties of fluorescent dissolved organic matter in the Gironde Estuary[J]. Organic Geochemistry, 2009, 40(6): 706-719. DOI:10.1016/j.orggeochem.2009.03.002 |
| [29] | Ohno T. Fluorescence inner-filtering correction for determining the humification index of dissolved organic matter[J]. Environmental Science & Technology, 2002, 36(4): 742-746. |
| [30] |
金相灿, 屠清瑛. 湖泊富营养化调查规范[M]. 第二版. 北京: 中国环境科学出版社, 1990. Jin X C, Tu Q Y. The standard methods in lake eutrophication investigation[M]. 2nd ed.. Beijing: China Environmental Science Press, 1990. |
| [31] |
吴丰昌, 王立英, 黎文, 等. 天然有机质及其在地表环境中的重要性[J]. 湖泊科学, 2008, 20(1): 1-12. Wu F C, Wang L Y, Li W, et al. Natural organic matter and its significance in terrestrial surface environment[J]. Journal of Lake Sciences, 2008, 20(1): 1-12. DOI:10.18307/2008.0101 |
| [32] | Xu H, Paerl H W, Qin B Q, et al. Nitrogen and phosphorus inputs control phytoplankton growth in eutrophic Lake Taihu, China[J]. Limnology and Oceanography, 2010, 55(1): 420-432. DOI:10.4319/lo.2010.55.1.0420 |
| [33] | Yao X, Zhang Y L, Zhu G W, et al. Resolving the variability of CDOM fluorescence to differentiate the sources and fate of DOM in Lake Taihu and its tributaries[J]. Chemosphere, 2011, 82(2): 145-155. DOI:10.1016/j.chemosphere.2010.10.049 |
| [34] | Wu F C, Tanoue E. Isolation and partial characterization of dissolved copper-complexing ligands in streamwaters[J]. Environmental Science & Technology, 2001, 35(18): 3646-3652. |
| [35] | Coble P G. Characterization of marine and terrestrial DOM in seawater using excitation-emission matrix spectroscopy[J]. Marine Chemistry, 1996, 51(4): 325-346. DOI:10.1016/0304-4203(95)00062-3 |
| [36] | Murphy K R, Stedmon C A, Waite T D, et al. Distinguishing between terrestrial and autochthonous organic matter sources in marine environments using fluorescence spectroscopy[J]. Marine Chemistry, 2008, 108(1-2): 40-58. DOI:10.1016/j.marchem.2007.10.003 |
| [37] | Yamashita Y, Jaffé R, Maie N, et al. Assessing the dynamics of dissolved organic matter (DOM) in coastal environments by excitation emission matrix fluorescence and parallel factor analysis (EEM-PARAFAC)[J]. Limnology and Oceanography, 2008, 53(5): 1900-1908. DOI:10.4319/lo.2008.53.5.1900 |
| [38] | Stedmon C A, Markager S. Tracing the production and degradation of autochthonous fractions of dissolved organic matter by fluorescence analysis[J]. Limnology and Oceanography, 2005, 50(5): 1415-1426. DOI:10.4319/lo.2005.50.5.1415 |
| [39] | Williams C J, Yamashita Y, Wilson H F, et al. Unraveling the role of land use and microbial activity in shaping dissolved organic matter characteristics in stream ecosystems[J]. Limnology and Oceanography, 2010, 55(3): 1159-1171. DOI:10.4319/lo.2010.55.3.1159 |
| [40] | Kowalczuk P, Cooper W J, Durako M J, et al. Characterization of dissolved organic matter fluorescence in the South Atlantic Bight with use of PARAFAC model:relationships between fluorescence and its components, absorption coefficients and organic carbon concentrations[J]. Marine Chemistry, 2010, 118(1-2): 22-36. DOI:10.1016/j.marchem.2009.10.002 |
| [41] | Stedmon C A, Markager S. Resolving the variability in dissolved organic matter fluorescence in a temperate estuary and its catchment using PARAFAC analysis[J]. Limnology and Oceanography, 2005, 50(2): 686-697. DOI:10.4319/lo.2005.50.2.0686 |
| [42] | Kramer G D, Herndl G J. Photo-and bioreactivity of chromophoric dissolved organic matter produced by marine bacterioplankton[J]. Aquatic Microbial Ecology, 2004, 36: 239-246. DOI:10.3354/ame036239 |
| [43] |
姚昕, 张运林, 朱广伟, 等. 湖泊草、藻来源溶解性有机质及其微生物降解的差异[J]. 环境科学学报, 2014, 34(3): 688-694. Yao X, Zhang Y L, Zhu G W, et al. Different degradation mechanism of dissolved organic matter derived from phytoplankton and macrophytes in Lake Taihu, China[J]. Acta Scientiae Circumstantiae, 2014, 34(3): 688-694. |
| [44] | Baker A. Fluorescence excitation-emission matrix characterization of river waters impacted by a tissue mill effluent[J]. Environmental Science & Technology, 2002, 36(7): 1377-1382. |
| [45] |
傅平青, 刘丛强, 吴丰昌. 溶解有机质的三维荧光光谱特征研究[J]. 光谱学与光谱分析, 2005, 25(12): 2024-2028. Fu P Q, Liu C Q, Wu F C. Three-dimensional excitation emission matrix fluorescence spectroscopic characterization of dissolved organic matter[J]. Spectroscopy and Spectral Analysis, 2005, 25(12): 2024-2028. DOI:10.3321/j.issn:1000-0593.2005.12.031 |
| [46] |
周永强, 张运林, 牛城, 等. 基于EEMs及PARAFAC的洪湖、东湖与梁子湖CDOM组成特征分析[J]. 光谱学与光谱分析, 2013, 33(12): 3286-3292. Zhou Y Q, Zhang Y L, Niu C, et al. Characterizing chromophoric dissolved organic matter (CDOM) in Lake Honghu, Lake Donghu and Lake Liangzihu using excitation-emission matrices (EEMs) fluorescence and parallel factor analysis (PARAFAC)[J]. Spectroscopy and Spectral Analysis, 2013, 33(12): 3286-3292. DOI:10.3964/j.issn.1000-0593(2013)12-3286-07 |
| [47] |
季乃云. 胶州湾荧光溶解有机物及其与环境因子关系的研究[D]. 青岛: 中国科学院研究生院(海洋研究所), 2005. Ji N Y. Fluorescent dissolved organic matter and its relationship with environmental factors in Jiaozhou Bay[D]. Qingdao: The Institute of Oceanology, Graduate University of Chinese Academy of Sciences, 2005. |
| [48] |
方芳, 翟端端, 郭劲松, 等. 三峡水库小江回水区溶解有机物的三维荧光光谱特征[J]. 长江流域资源与环境, 2010, 19(3): 323-328. Fang F, Zhai D D, Guo J S, et al. Three dimensional fluorescence spectroscopy characterization of dissolved organic matter in Xiaojiang River Backwater Area, Three Gorges Reservoir[J]. Resources and Environment in the Yangtze Basin, 2010, 19(3): 323-328. |
| [49] |
李崇明, 黄真理, 张晟, 等. 三峡水库藻类"水华"预测[J]. 长江流域资源与环境, 2007, 16(1): 1-6. Li C M, Huang Z L, Zhang S, et al. Risk forecast of algal bloom in the Three Gorges Reservoir[J]. Resources and Environment in the Yangtze Basin, 2007, 16(1): 1-6. |
| [50] |
王志刚, 刘文清, 李宏斌, 等. 三维荧光光谱法分析巢湖CDOM的空间分布及其来源[J]. 环境科学学报, 2006, 26(2): 275-279. Wang Z G, Liu W Q, Li H B, et al. Analysis of CDOM spatial distributions variations in Chaohu Lake and its sources by three dimensional fluorescence excitation-emission matrix[J]. Acta Scientiae Circumstantiae, 2006, 26(2): 275-279. |
| [51] |
傅平青, 吴丰昌, 刘丛强, 等. 高原湖泊溶解有机质的三维荧光光谱特性初步研究[J]. 海洋与湖沼, 2007, 38(6): 512-520. Fu P Q, Wu F C, Liu C Q, et al. Three-dimensional excitation emission matrix fluorescence spectroscopy of dissolved organic matter from Chinese highland lakes[J]. Oceanologia et Limnologia Sinica, 2007, 38(6): 512-520. |
| [52] |
江俊武, 李帅东, 沈胤胤, 等. 夏季太湖CDOM光学特性空间差异及其来源解析[J]. 环境科学研究, 2017, 30(7): 1020-1030. Jiang J W, Li S D, Shen Y Y, et al. Spatial differences of optical properties of CDOM and their source apportionment in Taihu Lake in summer[J]. Research of Environmental Sciences, 2017, 30(7): 1020-1030. |
| [53] |
曾庆飞, 孔繁翔, 张恩楼, 等. 利用稳定同位素技术研究外源物质输入对太湖微食物链的贡献[J]. 环境科学, 2007, 28(8): 1670-1674. Zeng Q F, Kong F X, Zhang E L, et al. Effects of anthropogenic organic matter inputs on stable carbon and nitrogen isotopes in organisms from microbial food chain in Taihu Lake[J]. Environmental Science, 2007, 28(8): 1670-1674. |
| [54] | Coble P G, Del Castillo C E, Avril B. Distribution and optical properties of CDOM in the Arabian Sea during the 1995 Southwest Monsoon[J]. Deep Sea Research Part Ⅱ:Topical Studies in Oceanography, 1998, 45(10-11): 2195-2223. DOI:10.1016/S0967-0645(98)00068-X |
| [55] | Smart P L, Finlayson B L, Rylands W D, et al. The relation of fluorescence to dissolved organic carbon in surface waters[J]. Water Research, 1976, 10(9): 805-811. DOI:10.1016/0043-1354(76)90100-7 |
| [56] |
季乃云, 赵卫红, 王江涛, 等. 大沽河-胶州湾段溶解有机物类腐殖质荧光特征变化[J]. 环境科学, 2006, 27(6): 1073-1077. Ji N Y, Zhao W H, Wang J T, et al. Change of humic-like fluorescence characteristics of dissolved organic matter from Dagu River to Jiaozhou Bay[J]. Environmental Science, 2006, 27(6): 1073-1077. |
| [57] |
周楠, 赵卫红, 苗辉. 獐子岛附近海域溶解有机物的荧光特征[J]. 光谱学与光谱分析, 2012, 32(4): 1007-1011. Zhou N, Zhao W H, Miao H. Fluorescence characteristics of dissolved organic matter in the sea nearby Zhangzi Island[J]. Spectroscopy and Spectral Analysis, 2012, 32(4): 1007-1011. |
| [58] |
陈锡超, 纪颖琳, 胡青, 等. "引江济太"水系有色溶解有机质的特征与来源[J]. 湖泊科学, 2010, 22(1): 63-69. Chen X C, Ji Y L, Hu Q, et al. Characteristics and sources of CDOM in the drainage system of Water Diversion from Yangtze River to Lake Taihu[J]. Journal of Lake Sciences, 2010, 22(1): 63-69. |
| [59] |
鲁景亮, 孙松, 张光涛, 等. 2011年夏季胶州湾表层溶解有机物荧光特征的时空变化[J]. 海洋科学, 2014, 38(4): 1-6. Lu J L, Sun S, Zhang G T, et al. Temporal and spatial variation of fluorescence characteristics of dissolved organic matters during summer of 2011 in Jiaozhou Bay[J]. Marine Sciences, 2014, 38(4): 1-6. |
| [60] | Mayer L M, Schick L L, Loder Ⅲ T C. Dissolved protein fluorescence in two Maine estuaries[J]. Marine Chemistry, 1999, 64(3): 171-179. DOI:10.1016/S0304-4203(98)00072-3 |
| [61] |
任保卫, 赵卫红, 王江涛, 等. 胶州湾围隔实验中溶解有机物三维荧光特征[J]. 环境科学, 2007, 28(4): 712-718. Ren B W, Zhao W H, Wang J T, et al. Three-dimensional fluorescence characteristic of dissolved organic matter in marine mesocosm experiment in Jiaozhou Bay, China[J]. Environmental Science, 2007, 28(4): 712-718. |
| [62] |
刘学利, 姚昕, 董杰, 等. 东平湖可溶性有机物的荧光特征及环境意义[J]. 生态与农村环境学报, 2016, 32(6): 933-939. Liu X L, Yao X, Dong J, et al. Fluorescence features of chromophoric dissolved organic matter in Dongping Lake and their environmental significance[J]. Journal of Ecology and Rural Environment, 2016, 32(6): 933-939. |
| [63] |
吕丽莎, 赵卫红, 苗辉. 三维荧光结合平行因子分析在东海溶解有机物研究中的应用[J]. 光谱学与光谱分析, 2013, 33(3): 653-658. Lü L S, Zhao W H, Miao H. Application of excitation-emission matrix spectrum combined with parallel factor analysis in dissolved organic matter in East China Sea[J]. Spectroscopy and Spectral Analysis, 2013, 33(3): 653-658. |
| [64] | Petersen H T. Determination of an Isochrysis galbana algal bloom by L-tryptophan fluorescence[J]. Marine Pollution Bulletin, 1989, 20(9): 447-451. DOI:10.1016/0025-326X(89)90065-9 |
| [65] | Rochelle-Newall E J, Fisher T R. Production of chromophoric dissolved organic matter fluorescence in marine and estuarine environments:an investigation into the role of phytoplankton[J]. Marine Chemistry, 2002, 77(1): 7-21. DOI:10.1016/S0304-4203(01)00072-X |
| [66] |
冯胜, 刘义, 余广彬. 微生物作用下的微囊藻降解试验研究[J]. 现代农业科技, 2009(1): 253-255. Feng S, Liu Y, Yu G B. Microbial degradation test of microcystis[J]. Modern Agricultural Sciences and Technology, 2009(1): 253-255. |