 2018, Vol. 39
2018, Vol. 39



2. 中国科学院环江喀斯特农业生态系统研究观测站, 环江 547200;
3. 中国科学院大学, 北京 100049;
4. 湖南省邵阳市第二中学, 邵阳 422000
2. Huanjiang Experimental Station of Karst Ecosystem, Chinese Academy of Sciences, Huanjiang 547200, China;
3. University of Chinese Academy of Sciences, Beijing 100049, China;
4. Shaoyang No.2 Middle School, Shaoyang 422000, China
氮是植物生长的必需元素, 也是陆地生态系统最主要的限制性营养元素[1].由于生态系统碳、氮循环之间的密切耦合关系, 土壤氮素供应能力很大程度上决定着全球变化背景下生态系统的固碳潜力及退耕后植被演替的方向和进程[2].同时, 土壤中某些形态氮也是环境中重要的污染物质, 比如排放到水体中的可溶性氮可导致水体富营养化[3], 排放到大气中的NOx可导致光化学烟雾[4].可见, 生态系统氮状况的研究对于评估全球变化背景下生态系统的固碳潜力、预测生态恢复方向与进程及污染防控等一系列重大生态环境问题均具有重要意义.土壤有效氮的生产包括了一系列过程, 如氨化、硝化和氮矿化过程等[5].这些过程是由一系列的真菌和细菌参与的[6].以硝化作用为例, 铵态氮氧化为硝态氮的过程是由自养和异养微生物共同驱动的.相比之下, 有机氮硝化为硝态氮的过程则主要是由异养微生物驱动.一般地, 自养硝化主要是由氨氧化细菌(AOB)和硝氧化细菌(NOB)主导, 而异养硝化则是比较广泛的细菌和真菌主导[7].通过添加生物抑制剂, 自养硝化、异养硝化、真菌硝化和细菌硝化过程是可以被区分开的[8], 而这样的区分对更好地理解生态系统氮循环过程有重要的意义.
生态系统氮素转化过程受多种全球变化因素的影响, 土地利用方式改变是其中重要的影响因素之一[9, 10].目前关于土地利用方式改变后(如:退耕后植被恢复)土壤氮素转化过程变化的研究已有不少报道.然而结果并不一致, Li等[11]的研究发现净氮矿化和硝化速率随演替而下降, 而氨化速率则不断增加; Yan等[12]的研究发现氮净矿化速率在演替初期和后期高, 中期低, 而净硝化速率则呈现随演替进程不断增加, 且不同季节变化趋势差异明显.生态系统类型、土壤母质和土壤类型是导致这些差异的主要原因.然而, 目前的研究主要集中在酸性土壤区域, 对中性偏碱性的喀斯特石灰土壤的氮素转化特征的认识还十分不足.此外, 对氮转化速率的研究多数只关心净(或总)氮转化速率, 对其具体的通路, 如自养、异养硝化过程的研究缺乏深入了解.
喀斯特是地表生态系统的重要组成部分, 其中中国西南喀斯特区是世界最大、最集中连片的喀斯特区[13].喀斯特山区独特的土壤条件(如高pH、高钙与高缓冲能力、特殊水文条件等)决定了生态系统过程对于全球变化的响应有别于其他地区.如前期的研究表明, 该地区的土壤氮含量很高, 硝态氮淋失严重, 意味着该地区土壤氮饱和的可能性极大[14].同时, 该地区正开展大规模的植被恢复工程, 其带来的对土壤氮转化过程的影响也不甚清楚.为此, 本文选取喀斯特地区4种不同土地利用方式的土壤(农田、草地、灌丛、次生林)为研究对象, 通过对喀斯特地区不同植被恢复阶段的氮矿化、氨化、硝化、真菌矿化、细菌矿化、真菌氨化、细菌氨化、真菌硝化、细菌硝化、自养硝化和异养硝化进行测定, 分析植被恢复过程对氮转化速率的影响及机制.本研究一方面可获取区域代表性数据, 另一方面可揭示喀斯特山区土壤肥力的供应和保持机制, 以期为喀斯特生态恢复实践提供理论支撑.
1 材料与方法 1.1 研究样地概况研究区位于广西环江县(107°53′~108°05′E, 25°06′~25°12′N), 具有典型喀斯特峰丛洼地景观, 属亚热带季风气候区.年平均气温为19.6~21.6℃, 最低温出现在1月, 约为3.4~8.7℃, 最高温出现在7月, 约为23.0~26.7℃.年均降雨量为1 530~1 820 mm, 分干湿季, 4~8月属于湿季, 9月至次年3月属于旱季.研究区基岩主要为石灰岩和白云岩, 土壤主要为石灰土[15].
1.2 土壤样品采集土壤采集时间在2014年7月.选择典型的农田、草地、灌丛和次生林这4种不同的植被恢复阶段.其中农田种植的是玉米.草地的恢复年限为10 a左右, 优势物种以五节芒(Miscanthus floridulus)、类芦(Neyraudia reynaudiana)等为主; 灌丛封育年限为15~20 a, 主要优势种为火棘(Pyracantha fortuneana)、金丝桃(Hypericum monogynum)和小果蔷薇(Rosa cymosa)等.次生林封育年限为30 a以上, 主要的优势种有伞花木(Eurycorymbus cavaleriei)、樟树(Cinnamomum camphora)、厚壳桂(Cryptocarya chinensis)等.
每个植被恢复阶段建立3个10 m×10 m的样方, 每个样方按S型布设5个样点, 采集表层(0~10 cm)土壤样品.将5个样点采取的土壤样品混成一个混合土样, 总共12个土壤样品.新鲜土样放于冰盒中带回实验室, 先用镊子挑去根和石块, 过2 mm筛后分两部分保存.一部分保存在4℃冰箱内用于土壤氮转化速率测定, 另一部分经自然风干后用于土壤理化性质分析(表 1).
|
|
表 1 不同植被恢复阶段下的土壤属性1) Table 1 Soil properties in different vegetation restoration stages |
1.3 实验方法 1.3.1 土壤潜在净氮转化速率测定
土壤潜在净氮转化速率的测定参考文献[16]的方法.具体步骤如下:将新鲜土样分为ABCDE 5组. A组:称5 g土测定土壤NH4+和NO3-浓度作为培养的初始值. B组:将10 g土壤放入250 mL培养瓶中, 不添加任何物质; C组:将10 g土壤放入250 mL培养瓶中, 添加纯度为99%的乙炔.乙炔的注入量为2%(体积比), 用以抑制微生物的自养硝化; D组:将10 g土壤放入250 mL培养瓶中, 添加2 mL的放线菌酮试剂, 用以抑制土壤的真菌活动; E组:将10 g土壤放入250 mL培养瓶中, 添加2 mL的链霉素试剂, 用以抑制土壤的细菌活动.
所有土壤样品, 调节含水量至土壤最大持水率的85%.用聚乙烯膜将B-E组的瓶口包扎.除B组的外, C-E组在膜上扎5个小孔以保证透气性, 然后将B-E组所有土壤置于25℃培养室中培养7 d.培养期间, 每隔两天调节一次含水率.培养结束后, 加入2 mol·L-1 KCl溶液100 mL, 于温度为25℃的恒温振荡机内振荡1 h后进行过滤, 滤液用AA3连续流动分析仪(Fiastar 5000)测定土壤中的NH4+和NO3-含量.土壤潜在净氮转化速率[以N计, mg·(kg·d)-1]的计算如下[16, 17].
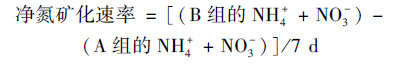
|
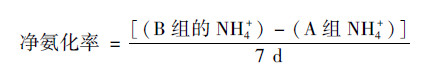
|

|

|
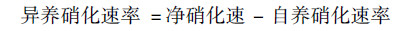
|

|
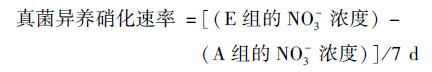
|

|

|
土壤理化指标的测定参考刘光崧等的方法[18].土壤pH采用pH计测定(土水比为1:2.5);土壤有机碳(SOC)采用重铬酸钾氧化-外加热法(油浴)测定; 全氮(TN)采用半微量开氏法-流动注射仪(FIAstar 5000)测定; 交换性K+、Ca2+、Na+、Mg2+采用乙酸铵交换-原子吸收分光光度法测定.土壤机械组成采用激光衍射粒度分析仪(Mastersizer, 2000, Malvern, UK)测定.土壤微生物量C和N (MBC和MBN)采用氯仿熏蒸法测定.此外还测定了与土壤有机氮分解相关的5种酶的活性.这些酶包括β-葡糖苷酶(BG)、α-纤维素酶(CBH)、亮氨酸氨基肽酶(LAP)、β-乙酰葡糖胺糖苷酶(NAG)和脲酶(Urease).这些酶的测定方法和功能见文献[19~21].
1.4 数据处理与分析利用单因素方差分析(One-way ANOVA)检验不同土地利用方式下土壤氮转化速率的差异, 并采用LSD方法进行显著性多重比较, 差异显著性水平为P < 0.05.利用线性回归分析土壤氮转化速率与土壤理化指标之间的相关性.以上分析在SPSS 19.0软件中进行.
2 结果与分析在4个喀斯特生态系统中, 土壤无机氮均以硝态氮为主, 占到总无机氮的(65%~93%)(表 2, A组).土壤培养7d后, 硝态氮在所有处理中均升高, 其中B组的升高幅度最大; C组、D组和E组相比B组有明显的下降.铵态氮在所有处理中也均显著升高, 其中C组、D组和E组的升高幅度大于B组.
|
|
表 2 不同处理下土壤硝态氮和氨态氮浓度的差异1)/mg·kg-1 Table 2 Concentrations of soil nitrate and ammonia under different treatments/mg·kg-1 |
2.1 土壤的净氨化、细菌氨化与真菌氨化速率
4种喀斯特生态系统类型的土壤净氨化速率均较低, 介于-0.16~0.63 mg·(kg·d)-1(图 1).方差分析结果表明, 随着植被的恢复, 土壤净氨化速率显著降低, 甚至在次生林阶段表现为负值(图 1).通过添加细菌和真菌抑制剂计算出来的细菌氨化[0.37~3.88 mg·(kg·d)-1]与真菌氨化速率[0.11~3.37 mg·(kg·d)-1]均高于总的净氨化速率(图 1).细菌氨化速率在次生林阶段显著低于农田、草地和灌丛, 而后3个阶段之间没有显著差异.真菌氨化速率表现出和细菌氨化速率一致的规律(图 1).

|
图 1 不同土地利用方式对土壤的净氨化、细菌与真菌氨化速率的影响 Fig. 1 Effect of different land use types on the net rates of ammonification, fungal and bacterial ammonification |
4种喀斯特生态系统类型的土壤净硝化速率均较高, 介于1.49~16.94 mg·(kg·d)-1, 其中自养硝化[1.24~13.02 mg·(kg·d)-1]相比异养硝化[0.25~3.92 mg·(kg·d)-1]占到总净氨化速率比重的76.8%~83.3%(图 2).

|
图 2 不同土地利用方式对土壤的总硝化、自养与异养硝化和真菌与细菌硝化的影响 Fig. 2 Effects of different land-use types on the net rates of soil nitrification, autotrophic and heterotrophic nitrification and fungal and bacterial nitrification |
细菌硝化介于0.80~4.46 mg·(kg·d)-1之间, 平均约占净氨化速率的41%;真菌硝化速率介于1~6.46 mg·(kg·d)-1之间, 平均约占59%(图 3).以上几种硝化速率类型均表现为随着植被恢复不断增加的趋势, 其中在草地最低, 次生林最高.

|
图 3 异养硝化、自养硝化、细菌硝化和真菌硝化占总净硝化的占比 Fig. 3 Contributions of heterotrophic, autotrophic, bacterial, and fungal nitrification to the total net nitrification |
土壤净氮矿化速率介于2.00~16.78 mg·(kg·d)-1, 在次生林阶段显著低于农田、草地和灌丛(图 4).细菌氮矿化[0.37~3.88 mg·(kg·d)-1]与真菌氮矿化速率[0.11~3.37 mg·(kg·d)-1]也表现出相同的规律.而且除了次生林外, 真菌和细菌氮矿化均高于总的净氮矿化速率.

|
图 4 不同土地利用方式对土壤的氮矿化、细菌和真菌氮矿化的影响 Fig. 4 Effects of different land-use types on the net rates of soil nitrogen-mineralization, bacterial and fungal nitrogen-mineralization |
相关性分析的结果显示, 土壤氮转化速率与土壤有机碳、总氮、硝态氮、微生物量C、微生物量N及5种与氮获取相关的酶的活性关系密切, 而与土壤pH、碳氮比、铵态氮、土壤钾钙钠镁的含量无显著相关(表 3).其中, 土壤有机碳与净氮矿化、真菌矿化、细菌矿化、净硝化、异养硝化、自养硝化、真菌硝化和细菌硝化呈显著正相关; 土壤总氮与净氮矿化、真菌矿化、细菌矿化、净氨化、净硝化、异养硝化、自养硝化、真菌硝化和细菌硝化呈显著正相关; 而土壤硝态氮与测定的所有氮转化速率均成正相关.微生物量C与净氮矿化、细菌矿化、净氨化、细菌氨化、净硝化、自养硝化成正相关.微生物量N与净氮矿化、净氨化成正相关. 5种酶中, LAP的相关性最强, 与净氮矿化、真菌矿化、细菌矿化、细菌氨化、净硝化、异养硝化、自养硝化和细菌硝化均呈显著正相关.
|
|
表 3 土壤氮转化速率与土壤属性的相关分析结果1)(P值) Table 3 Results of correlation analysis for the relationships between soil nitrogen transformation rate and soil properties |
3 讨论 3.1 喀斯特地区土壤氮转化的特征
喀斯特钙质土壤的无机氮主要以硝态氮形式存在, 而铵态氮所占比例很低(表 2).该结果与其他钙质土壤地区的研究一致[22], 而与酸性土壤的研究相反:一般酸性土壤中的无机氮以铵态氮为主[23].偏碱性的钙质土壤硝态氮含量高的原因可能是较高的硝化速率导致的[24].一般认为, 硝化作用在pH < 5的时候很低, 而随着土壤pH值的升高(从4升至8)硝化作用不断增加[25].以此相符, 本研究中喀斯特钙质土壤的硝化速率很高, 相反地, 氨化速率非常低, 甚至在次生林中表现出负值.较高的硝化速率使得氨化产生的铵态氮很快被消耗.当总硝化速率超过了总氨化速率时, 净氨化速率则会表现为负值.
研究发现在添加了真菌或细菌抑制剂后, 无论是真菌还是细菌氨化都比总的净氨化速率高.这和一般的认知相反.一般地添加抑制剂后, 氨化速率都会下降.比如, Trap等[16]的研究发现, 在添加抑制剂后, 真菌氨化和细菌氨化均比净氨化速率低.导致本研究中真菌氨化和细菌氨化均高于对照的净氨化速率的原因还不清楚.推测可能是由于添加生物抑菌剂在杀死硝化细菌的同时也杀死了氨化细菌.由于喀斯特地区的硝化速率较高, 相同比例的硝化细菌被杀死后, 可能导致硝化速率下降, 使得铵态氮累积, 最终表现为净氨化速率的上升.本研究确实发现添加真菌和细菌抑制剂后铵态氮增加了, 但硝化速率和硝态氮浓度均下降(表 2), 这个结果支持了上述的解释.
本研究还发现自养硝化相比异养硝化占更重要的地位.这和目前的认知是一致的.自养硝化细菌在低pH的环境中含量很低, 所以一般认为在酸性土壤中自养硝化很弱, 相反地, 在中性或碱性土壤(pH为7~8), 自养硝化速率达到最高[4].此外, 在喀斯特生态系统中细菌和真菌对硝化的贡献均较高, 分别占41%和59%.之前的研究结果表明, 喀斯特地区土壤的细菌约占总微生物量的11.3%~63.6%[26], 略低于真菌的比例.这部分支持了以上的结果.然而, 进一步的研究有待揭示起作用的真菌与细菌的具体种类及其作用机制.
3.2 植被恢复对喀斯特土壤氮素转化速率的影响随着植被恢复, 土壤净氮矿化速率(包括其子通路)和净硝化速率(包括其子通路)从农田到森林不断增加, 而净氨化作用(包括其子通路)不断下降.氮矿化速率和硝化速率随着植被恢复不断上升的规律与之前的研究一致.比如, 刘欣等的研究发现[27], 随着植被的正向演替(草丛灌丛次生林原生林), 土壤氮净矿化速率、净硝化速率均呈增加趋势.然而, 其测定的净氨化速率也呈现增加的趋势, 这与本研究的结果相反.目前关于喀斯特地区土壤氮素转化特征随植被恢复变化的研究较少.因此, 到底氨化速率随植被恢复后如何改变还需进一步的研究.本研究推测净氨化作用下降可能是由于总硝化作用的上升幅度高过了总氨化作用, 从而导致氨氮的生成赶不上其消耗, 最终表现为净氨化作用的下降.虽然本研究并没有测定总氨化和总硝化速率, 但是相关分析显示, 土壤净氨化与土壤硝态氮成显著负相关关系.这个结果部分支持了上述解释.
而导致净氮矿化和硝化速率上升的原因可能有两个.第一个原因与土壤碳氮水平有关.一般认为土壤中的有机碳和氮的增加会导致总氮矿化或者硝化的作用增加, 因为其增加了微生物的能量和底物来源[27].本研究中土壤有机碳, 有机碳与净矿化硝化有显著的正相关关系, 说明土壤有机碳、氮含量的增加确实对土壤净矿化消化速率的上升有显著贡献.与本研究的结果一致, 刘欣等[27]及周练川等[28]也发现相似的结果.第二, 微生物量及酶活性也是导致土壤氮矿化和硝化速率上升的关键因素.微生物是土壤生物化学循环的主要驱动者[29], 因此土壤微生物量的大小直接决定了土壤氮素转化的速率.本研究发现土壤氮矿化和硝化速率与土壤微生物量C有显著的正相关关系, 支持了上述观点.同时, 土壤有机氮的释放是在一系列氮获取酶的催化下进行的[20].这些酶对土壤有机氮的降解被认为是土壤氮循环的主要限速步骤[20].通过测定与土壤有机氮分解相关的酶的活性(包括BG、CBH、LAP、NAG、Urease), 发现这些酶的活性均与土壤氮转化速率有极强的相关性, 尤其是蛋白质水解酶(LAP酶)相关性最高(表 3), 说明蛋白质的降解可能是土壤氮矿化的最重要的限速步骤.
4 结论(1) 喀斯特区土壤硝化作用强烈, 导致土壤无机氮主要以硝态氮为主.
(2) 喀斯特土壤硝化作用主要以自养硝化为主, 真菌和细菌均对土壤硝化有重要贡献.
(3) 随着植被恢复, 土壤净氮矿化速率(包括其子通路)和净硝化速率(包括其子通路)从农田到森林不断增加, 而净氨化作用(包括其子通路)不断下降.
| [1] | LeBauer D S, Treseder K K. Nitrogen limitation of net primary productivity in terrestrial ecosystems is globally distributed[J]. Ecology, 2008, 89(2): 371-379. DOI:10.1890/06-2057.1 |
| [2] | Reich P B, Hungate B A, Luo Y Q. Carbon-nitrogen interactions in terrestrial ecosystems in response to rising atmospheric carbon dioxide[J]. Annual Review of Ecology, Evolution, and Systematics, 2006, 37: 611-636. DOI:10.1146/annurev.ecolsys.37.091305.110039 |
| [3] | Ryther J H, Dunstan W M. Nitrogen, phosphorus, and eutrophication in the coastal marine environment[J]. Science, 1971, 171(3975): 1008-1013. DOI:10.1126/science.171.3975.1008 |
| [4] | Fabian P. Photochemischer smog und seine einwirkung auf die biosphäre[J]. Forstwissenschaftliches Centralblatt, 1987, 106(1): 223-235. DOI:10.1007/BF02741152 |
| [5] |
方运霆, 莫江明, Gundersen P, 等. 森林土壤氮素转换及其对氮沉降的响应[J]. 生态学报, 2004, 24(7): 1523-1531. Fang Y T, Mo J M, Gundersen P, et al. Nitrogen transformations in forest soils and its responses to atmospheric nitrogen deposition:a review[J]. Acta Ecologica Sinica, 2004, 24(7): 1523-1531. |
| [6] | Hayatsu M, Tago K, Saito M. Various players in the nitrogen cycle:diversity and functions of the microorganisms involved in nitrification and denitrification[J]. Soil Science and Plant Nutrition, 2008, 54(1): 33-45. DOI:10.1111/j.1747-0765.2007.00195.x |
| [7] | De Boer W, Kowalchuk G A. Nitrification in acid soils:micro-organisms and mechanisms[J]. Soil Biology and Biochemistry, 2001, 33(7-8): 853-866. DOI:10.1016/S0038-0717(00)00247-9 |
| [8] | Brierley E D R, Wood M. Heterotrophic nitrification in an acid forest soil:isolation and characterisation of a nitrifying bacterium[J]. Soil Biology and Biochemistry, 2001, 33(10): 1403-1409. DOI:10.1016/S0038-0717(01)00045-1 |
| [9] | Booth M S, Stark J M, Rastetter E. Controls on nitrogen cycling in terrestrial ecosystems:a synthetic analysis of literature data[J]. Ecological Monographs, 2005, 75(2): 139-157. DOI:10.1890/04-0988 |
| [10] | Barnes R T, Raymond P A. Land-use controls on sources and processing of nitrate in small watersheds:insights from dual isotopic analysis[J]. Ecological Applications, 2010, 20(7): 1961-1978. DOI:10.1890/08-1328.1 |
| [11] | Li M, Zhou X H, Zhang Q F, et al. Consequences of afforestation for soil nitrogen dynamics in central China[J]. Agriculture, Ecosystems & Environment, 2014, 183: 40-46. |
| [12] | Yang Y H, Luo Y Q, Finzi A C. Carbon and nitrogen dynamics during forest stand development:a global synthesis[J]. New Phytologist, 2011, 190(4): 977-989. DOI:10.1111/nph.2011.190.issue-4 |
| [13] |
彭晚霞, 王克林, 宋同清, 等. 喀斯特脆弱生态系统复合退化控制与重建模式[J]. 生态学报, 2008, 28(2): 811-820. Peng W X, Wang K L, Song T Q, et al. Controlling and restoration models of complex degradation of vulnerable Karst ecosystem[J]. Acta Ecologica Sinica, 2008, 28(2): 811-820. |
| [14] | Wen L, Li D J, Yang L Q, et al. Rapid recuperation of soil nitrogen following agricultural abandonment in a karst area, southwest China[J]. Biogeochemistry, 2016, 129(3): 341-354. DOI:10.1007/s10533-016-0235-3 |
| [15] | Zhang W, Zhao J, Pan F J, et al. Changes in nitrogen and phosphorus limitation during secondary succession in a karst region in southwest China[J]. Plant and Soil, 2015, 391(1-2): 77-91. DOI:10.1007/s11104-015-2406-8 |
| [16] | Trap J, Bureau F, Akpa-Vinceslas M, et al. Changes in humus forms and soil N pathways along a 130-year-old pure beech forest chronosequence[J]. Annals of Forest Science, 2011, 68(3): 595-606. DOI:10.1007/s13595-011-0063-5 |
| [17] | Trap J, Bureau F, Vinceslas-Akpa M, et al. Changes in soil N mineralization and nitrification pathways along a mixed forest chronosequence[J]. Forest Ecology and Management, 2009, 258(7): 1284-1292. DOI:10.1016/j.foreco.2009.06.021 |
| [18] |
刘光崧, 蒋能慧, 张连第, 等. 土壤理化分析与剖面描述[M]. 北京: 中国标准出版社, 1996: 137-183. Liu G S, Jiang N H, Zhang L D, et al. Soil physical and chemical analysis & description of soil profiles[M]. Beijing: Standards Press of China, 1996: 137-183. |
| [19] | Chen H, Luo P, Wen L, et al. Determinants of soil extracellular enzyme activity in a karst region, southwest China[J]. European Journal of Soil Biology, 2017, 80: 69-76. DOI:10.1016/j.ejsobi.2017.05.001 |
| [20] | Chen H, Li D J, Zhao J, et al. Effects of nitrogen addition on activities of soil nitrogen acquisition enzymes:a meta-analysis[J]. Agriculture, Ecosystem & Environment, 2018, 252: 126-131. |
| [21] |
刘璐, 陈浩, 李德军, 等. 喀斯特山区植被恢复过程中土壤水解酶和氧化酶活性的响应[J]. 环境科学学报, 2017, 37(9): 3528-3534. Liu L, Chen H, Li D J, et al. Changes of soil hydrolytic and oxidized enzyme activities under the process of vegetation restoration in a karst area, southwest China[J]. Acta Scientiae Circumstantiae, 2017, 37(9): 3528-3534. |
| [22] | Zhu T B, Zeng S M, Qin H L, et al. Low nitrate retention capacity in calcareous soil under woodland in the karst region of southwestern China[J]. Soil Biology and Biochemistry, 2016, 97: 99-101. DOI:10.1016/j.soilbio.2016.03.001 |
| [23] | Chen H, Gurmesa G A, Zhang W, et al. Nitrogen saturation in humid tropical forests after 6 years of nitrogen and phosphorus addition:hypothesis testing[J]. Functional Ecology, 2016, 30(2): 305-313. DOI:10.1111/1365-2435.12475 |
| [24] | Sibbald I R, Bowland J P, Robblee A R, et al. The influence of the nitrogen source on the food intake and nitrogen retention of weanling rats[J]. The Journal of Nutrition, 1957, 62(2): 185-200. DOI:10.1093/jn/62.2.185 |
| [25] | McGrath S P, Rorison I H. The influence of nitrogen source on the tolerance of Holcus lanatus L.* and Bromus erectus huds.to manganese[J]. New Phytologist, 1982, 91(3): 443-452. DOI:10.1111/nph.1982.91.issue-3 |
| [26] |
宋敏, 邹冬生, 杜虎, 等. 不同土地利用方式下喀斯特峰丛洼地土壤微生物群落特征[J]. 应用生态学报, 2013, 24(9): 2471-2478. Song M, Zou D S, Du H, et al. Characteristics of soil microbial populations in depressions between karst hills under different land use patterns[J]. Chinese Journal of Applied Ecology, 2013, 24(9): 2471-2478. |
| [27] |
刘欣, 黄运湘, 袁红, 等. 植被类型与坡位对喀斯特土壤氮转化速率的影响[J]. 生态学报, 2016, 36(9): 2578-2587. Liu X, Huang Y X, Yuan H, et al. Effects of vegetation type and slope position on soil nitrogen transformation rate in Karst regions[J]. Acta Ecologica Sinica, 2016, 36(9): 2578-2587. |
| [28] |
周炼川, 陈效民, 李孝良, 等. 西南喀斯特地区典型石漠化阶段土壤氮素变异研究[J]. 农业环境科学学报, 2009, 28(8): 1753-1757. Zhou L C, Chen X M, Li X L, et al. Variability of soil nitrogen in karst regions under typical rocky desertification stage, southwest China[J]. Journal of Agro-Environment Science, 2009, 28(8): 1753-1757. |
| [29] | Cusack D F, Silver W L, Torn M S, et al. Changes in microbial community characteristics and soil organic matter with nitrogen additions in two tropical forests[J]. Ecology, 2011, 92(3): 621-632. DOI:10.1890/10-0459.1 |