 2018, Vol. 39
2018, Vol. 39



华北平原是中国主要的冬小麦(Trticum aestivums L.)产区, 产量占国内总产量的2/3[1], 研究该区域冬小麦生产对于土壤有机碳的影响具有重要科学和实践价值.农田土壤的有机碳主要来自作物, 除根和茎叶残留之外, 活体植物分泌的根系淀积物(根系脱落物和根系分泌物)也是土壤有机碳的重要来源[2, 3].来自植物凋落物的碳比较容易定量[2], 但对于根系淀积物利用传统洗根法则较为困难, 主要由于以下原因:①在一定时间内, 根系淀积碳含量远低于土壤中已存在的其他有机物质(如植物凋落物、半分解的有机物和土壤生物的代谢产物等)[2]; ②通过根释放到土壤的有机物半衰期为0.5~10 d, 根系淀积碳在土壤中很快被分解[2]; ③根系淀积物只存在于靠近根表面一个很小的区域内, 分离非常困难[2].碳同位素标记法是利用碳同位素在植物体内和土壤有机物中的差别, 被很多学者广泛用于定量根系淀积物和根际呼吸的碳量(区分土壤呼吸), 进而定量植物地上部转移到根系和土壤的总光合碳量[2, 4~6].通过标记植物法所获得的试验结果比洗根法或根系生长评估法高3~7倍[2].
在诸多碳同位素中, 13C不需安全防护, 且13C与12C的性质差异, 比14C与12C要小, 因而13C被视为更可靠的示踪物[7].与连续标记法相比, 脉冲标记比较容易掌握, 设备要求简单、费用低, 能够提供植物各生育时期光合碳的分配信息[8, 9].虽然一次脉冲标记得到的碳同位素分配信息不能代表植物整个生育期, 但只要在植物生育期内进行一系列标记, 就可以合理地估计植物输入地下各组分的碳量[2, 6, 10~13].因此, 本研究选择13CO2脉冲标记方法, 来定量分析华北平原冬小麦生长对于地下部分碳的贡献.
目前, 国内外同行在冬小麦碳同位素脉冲标记研究方面获得了很多数据, 但由于标记方法和试验条件不同, 所获得的试验结果有很大差异, 且可比性差, 更不能直接应用于国内的农田土壤.此外, 由于技术条件的限制, 国内冬小麦碳标记的定量输入的研究开展较少, 仅有屈波等[14]和齐鑫等[15]分别使用14CO2和13CO2脉冲标记法的研究.因此, 本研究选择在长示踪期条件下, 探究冬小麦不同生育时期光合碳在植物-土壤系统中的分配, 定量冬小麦向地下各组分转移的光合碳, 以期为我国农田土壤碳分配增加数据积累, 加深对冬小麦-土壤系统中地下部碳平衡过程的理解, 对于我国农业固碳减排具有重要意义.
1 材料与方法 1.1 冬小麦种植试验于中国农业大学桓台试验站田间进行, 站点坐标为E117°59′, N36°57′, 海拔高度为18 m.所选盆(PVC柱)大小为20 cm×30 cm(直径×高度).供试土壤取自试验站农田(0~20 cm), 土壤有机碳的δ13C值为-24‰.供试土壤为潮土类沙壤土质[16], 相关土壤参数为:土壤有机碳8.4 g·kg-1, 土壤无机碳5.2 g·kg-1, 全氮0.67 g·kg-1, pH值=8.2(水土比为2.5:1), 速效钾174 mg·kg-1, 速效磷5.2 mg·kg-1.土壤风干后, 磨碎、挑根, 再过5 mm筛.每盆装风干土8.7 kg(土深23 cm左右).磷肥和钾肥(KH2PO4)作为底肥一次性施入, 氮肥作为底肥、拔节肥2次施入尿素(含氮量为46%), 两次施用量的比例均为50%, 用量分别为P2O5 78 mg·kg-1、K2O 43 mg·kg-1和N 90 mg·kg-1.供试作物为冬小麦(Triticum aestivums L., 鲁源502).冬小麦种子播种前放在清水中浸泡10h, 然后再浅埋入土壤中. 2014年10月15日播种, 每盆10粒, 出苗一周后, 每盆留6株.用称重法分别控制土壤水分为田间持水量的75%.小麦生长期间, 根据病虫害情况, 喷洒必要的农药.
1.2 13C脉冲标记分别在播种后75 d(分蘖期)、168 d(拔节期)、188 d(花期)和202 d(灌浆期)进行4次标记.每次选取4盆小麦, 放入标记室进行13C脉冲标记(图 1). ①标记前, 连接CO2吸收装置, 让标记过程中根际呼吸产生的13CO2汇聚在盆内; ②将盆内土壤和隔板间的空气与标记室空气隔开, 把隔板和PVC柱的接合处用凡士林密封, 并在小麦基部与隔板的间隙中涂上凡士林密封; ③检查密闭性:把气球套在一空心管上, 在另一端管口用吸耳球鼓气, 根据气球膨胀状态判定密封性好坏.然后把空心管管口堵住; ④将风扇、小麦植株和13C丰度为99%的Na213CO3放入标记室内, 然后将整个标记室密封; ⑤标记于上午09:00点开始, 用注射器向装有Na213CO3的烧杯中注入一定量1 mol·L-1的H2SO4溶液(根据Na213CO3计算好硫酸的用量), 标记开始.此后每隔一段时间, 根据CO2检测仪的读数来确定是否加入硫酸, 当低于200×10-6时, 向烧杯中注入1次硫酸, 以维持CO2体积分数在360×10-6左右, 标记时间持续7 h; ⑥标记结束后, 标记罩子内残留的13CO2气体通过气泵和NaOH溶液回收(回收0.5 h)[6, 17].

|
1.小风扇; 2.CO2浓度检测仪; 3.注射1 mo·L-1 H2SO4溶液; 4.1 mol·L-1 Na213CO3溶液; 5.盆栽小麦; 6.土壤 图 1 标记装置 Fig. 1 Labeling apparatus |
脉冲标记的示踪时间应足够长, 以保证小麦在标记时吸收的标记13C在小麦-土壤整个系统完全分配[6, 13, 18], 过早取样, 会高估进入到土壤有机碳中的碳量, 低估根际呼吸消耗的碳; 取样过晚, 又徒增收集土壤呼吸的劳动量, 给试验的顺利进行增加难度, 增加费用.结合其他研究者的结果, 冬小麦光合固定的14C在小麦-土壤整个系统分配至少需要19 d[18], 示踪期宜长不宜短, 因此, 本试验选择了28 d的示踪期.
1.3 取样标记结束后, 每2~5 d更换1次NaOH溶液.每天定时用空气泵在土壤与隔板间注入一定量无CO2的空气, 用于提供氧气.标记结束后第28 d, 对小麦进行破坏性取样.从基部剪断小麦植株, 将全部土壤反复过2 mm筛, 挑出根系, 并把土壤平铺于农用地膜上.把挑出的根连同附着于根上的土壤用水浸泡, 轻轻振荡, 20 min后, 过1 mm筛, 并把筛上的根系冲洗干净.然后把浸泡和冲洗小麦根的用水、沉淀的泥浆均匀地洒在平铺于地膜上的土壤中.将土壤用多次划网格取样的方式缩分, 最后取约50 g土壤.将小麦地上部和根分别置于105℃下杀青30 min, 60℃下烘干至恒重.
1.4 分析和测定 1.4.1 土壤呼吸13C丰度的样品制备把28 d内9次(第2、4、7、11、14、17、20、26和28 d)用于固定呼吸的NaOH溶液, 等体积混合(每次吸取10 mL), 将过量SrCl2加入到28 d内混合NaOH溶液和最后2 d的NaOH溶液中(26~28 d), 形成SrCO3沉淀.以酚酞作指示剂, 用稀盐酸滴定未与CO2反应的NaOH, 根据稀盐酸和NaOH的体积、浓度计算吸收的CO2量.将沉淀的SrCO3搅拌混匀, 60℃下烘干至恒重.
1.4.2 土壤有机碳13C丰度的样品制备从50 g土壤样品中取约20 g土壤置于白色平板上, 挑去残留细根.然后, 将其浸泡在100 mL去离子水中振荡30 min, 使土壤中水溶性有机碳充分溶解于水.土壤溶液静置一段时间后, 将上清液收集于烧杯内, 防止在以后冲洗HCl时, 损失水溶性有机碳.再往泥浆中加入50 mL 3 mol·L-1的HCl溶液, 用于去除泥浆中的碳酸盐.摇匀后放入振荡器以200 r·min-1振荡30 min后, 静置12 h; 缓缓将上清液倒掉, 用去离子水洗去土样中的酸, 加入去离子水, 充分搅拌均匀并静置一段时间后, 放入离心机中以3 000 r·min-1的转速离心3 min, 将上清液倒掉, 重复此过程, 用pH试纸检测上清液的pH值, 洗到中性为止, 并把酸化前的上清液倒回烧杯中, 在60℃条件下烘干[6, 19].
1.4.3 冬小麦植株、土壤和土壤呼吸13C丰度的测定测定之前利用球磨仪(Restol MM2000, Retsch, Haan, Germany)研磨(< 500 μm).上述各类样品13C丰度, 分别在DELTAplus XP型质谱仪(DELTAplus XP, ThermoFinnigan, Bremen, Germany)中测定.利用Thermo Elemental Analyzer 1112元素分析仪测定植物碳含量和土壤有机碳含量.碳同位素采用PDB(Peedee Belemnite)标准.
1.5 计算方法冬小麦光合固定的13C最终进入到小麦地上部、根部、土壤有机碳和土壤呼吸这4个组分中.进入各个组分的13C量(mg·pot-1)[6, 15]:
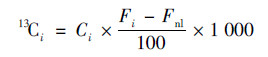
|
(1) |
式中, 13Ci为各组分的13C量, Ci为各组分的碳量, Fi和Fnl为各组分标记和不标记的13C丰度.
冬小麦净吸收的13C量计算公式为[6]:

|
(2) |
13C在各组分的分配比例(%):

|
(3) |
式中, 13Ci%为该组分的13C量占净吸收13C量的百分比, %.
各个时期输入到各个组分的碳量(g·pot-1)[6, 20, 21]:
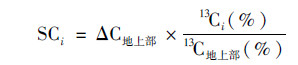
|
(4) |
式中, ΔC地上部为每一生育时期地上部碳量的变化量, 以地上部作为参照是考虑到地上部的生物量和含碳量可以准确定量.
本研究使用上述公式计算小麦根际呼吸、输入到土壤有机碳和根的光合碳量, 是基于下列理论:本试验中4次标记13C的分配规律, 分别代表冬小麦分蘖期、拔节期、开花期和灌浆期各自吸收光合碳的平均分配规律.该计算方法虽然是一个估算, 但这种估算对于研究植物-土壤系统的碳平衡很有意义[22].
1.6 数据分析方差分析用SPSS 17.0软件计算.同一组分不同生育期的干物质累积量、根冠比、地上部占植株总质量的比例、13C分配比例和输入到地下各组分碳量之间的显著性差异分析均设为P < 0.05水平, 且平均值比较采用最小显著差异法(least significant difference, LSD).制图采用Excel 2013软件.
2 结果与分析 2.1 不同生育期冬小麦的干物质累积量冬小麦的干物质累积量随着生育期的增长而增加[图 2(a)].冬小麦根冠比随生育期的增长而下降, 由分蘖期的0.63显著下降到花期的0.14, 然后在后3个生育时期保持稳定[图 2(b)].冬小麦地上部生物量占植株总重的比例随生育期的增长而降低, 由分蘖期的0.62增加到收获期的0.94.在后3个生育期, 随着生育期的增长, 籽粒占整个植株总重的比例而增加(从花期的0.30上升到收获期的0.56), 秸秆占植株总重的比例而下降, 从花期的0.60下降到收获期的0.38[图 2(c)].

|
T-T、E-A、A-G和G-H分别代表在分蘖期标记-分蘖期收样、拔节期标记-开花期收样、花期标记-灌浆期收样和灌浆期标记-收获期收样; 不同小写字母表示同一组分不同生育期间的显著性差异(P < 0.05) 图 2 冬小麦不同生育时期的干物质累积量、根冠比和地上部占植株总重的比值 Fig. 2 Dry weight accumulation, root/shoot, and shoot/total plant weight of winter wheat at different growth stages |
光合固定的13C大部分(质量分数64.3%~90.3%)保留在地上部(籽粒+秸秆), 随着生育期的增长而增加, 即从分蘖期的64.3%增加到灌浆期的90.3%(图 3).随着生育期的增长, 光合固定的13C在籽粒中的比例增加, 从拔节期的5.1%增加到灌浆期的60.9%, 在秸秆中的分配反而下降(从拔节期的67.9%下降到灌浆期的29.4%).在地下部的13C比例与在地上部的截然相反:分配到地下部(根+土壤有机碳+根际呼吸)的13C比例随生育期的增长而降低, 由分蘖期的35.7%下降到灌浆期的9.7%, 根、土壤有机碳和根际呼吸中13C的比例也是随生育期的增长而降低.除分蘖期外, 在其它3个生育期, 地下部各个组分的13C比例在根际呼吸最大, 土壤中最小, 在分蘖期, 根中的比例最大, 土壤中的最小.

|
T、E、A和G分别代表在分蘖期标记、拔节期标记、花期标记和灌浆期标记,下同 图 3 光合13C在土壤-冬小麦体系各组分中的分配(平均值±SE) Fig. 3 Proportion (mean±SE) of photosynthesized 13C in each component of the wheat-soil system |
在4个生育期, 冬小麦向地下部转移的13C分别占净吸收量的质量分数为35.7%、27.0%、15.1%和9.7%(图 3).在向地下部转移的13C中(图 4), 大部分13C都被根际呼吸所消耗(除分蘖期外), 在灌浆期时达到最大(65.3%), 在花期最小(56.7%), 并且拔节期(62.9%)和花期之间无显著差异.分配到根的13C与根际呼吸则相反, 随生育期的增长而下降, 在分蘖期最高(59.3%), 在灌浆期仅为24.3%.进入土壤有机碳的13C比例在分蘖期最高(17.8%), 显著高于后3个生育期的比例(10.4%~14.3%之间), 在拔节期和花期之间无显著差异, 显著高于灌浆期的比例.示踪期28 d最后2 d产生的13C只占整个示踪期根际呼吸产生13C量的0.7%~2.7%(图 5).

|
图 4 输入到地下部分的光合13C在地下各组分中的分配 Fig. 4 Proportion of photosynthesized 13C input belowground in each component belowground |

|
图 5 示踪期最后2 d占到总呼吸释放13C的比例 Fig. 5 Proportion of respired 13C collected within the last 2 days in total respired 13C over the whole chase period |
冬小麦根际呼吸碳量在拔节期最高, 为3.3 g·pot-1, 其余3个生育时期在0.2~0.8 g·pot-1之间, 整个生育期内根际呼吸总碳量为4.6 g·pot-1(图 6).冬小麦根系通过根际沉积, 将一部分光合碳转化为土壤有机碳, 也是在拔节期最高, 为0.8 g·pot-1, 其余3个生育期则较低(0.1~0.2 g·pot-1), 且生育期之间无显著差异, 冬小麦在整个生育期内向土壤有机碳中输入的光合碳量为1.1 g·pot-1.在整个生育期内, 冬小麦用于根系组织建成的总碳量为2.2 g·pot-1, 因此, 冬小麦在整个生育期内输入到地下部的光合碳量为8.0 g·pot-1[输入地下部的碳量=根(2.2 g·pot-1)+土壤有机碳(1.1 g·pot-1)+根际呼吸(4.6 g·pot-1); 图 6], 占净吸收光合碳量的21.5%[净吸收光合碳量=地上部(29.2 g·pot-1)+地下部(8.0 g·pot-1)].冬小麦在整个生育期内输入到地上部、根部、土壤有机碳和根际呼吸中的光合碳量, 分别占净吸收光合碳量的比例为78.5%、6.0%、3.1%和12.4%(表 1).

|
T、E、A、G 和 W 分别代表在分蘖期、拔节期、花期、灌浆期和整个生育期输入到地下各组分的碳量 图 6 小麦在生育期内输入到地下各组分的碳量(平均值±SE) Fig. 6 Net photosynthesized C inputs (mean±SE) into each component of the wheat-soil system over the whole wheat growing season |
|
|
表 1 不同生育时期脉冲标记碳在冬小麦-土壤体系的分配(平均值±SE) /% Table 1 Pulse labeled carbon partitioning (mean±SE) in the winter wheat-soil system at different growth stages/% |
结合冬小麦整个生育期的分配比例(表 1), 以华北平原小麦地上部干重15 t·hm-2[23]、含碳量42%计算, 冬小麦在整个生育期内输入到地下部的总碳量为1.72 t·hm-2, 其中0.99 t·hm-2被根际呼吸消耗, 释放回大气中, 用于建成根系组织的碳量为0.48 t·hm-2, 0.25 t·hm-2则转化为土壤有机碳.
3 讨论 3.1 不同生育期冬小麦光合碳的分配规律冬小麦生长时期不同, 光合产物的分配规律各异[5, 24].本试验中光合固定的13C大部分保留在植株地上部(64.3%~90.3%), 随生育期的增长, 地上部的光合碳数量和比例不断增加, 而向地下部比例则不断减少, 如地上部光合碳从分蘖期的64.3%上升到到灌浆期的90.3%, 地下部光合碳由分蘖期的35.7%下降到灌浆期的9.7%(图 3).这和其他研究者小麦各个关键生育期的标记结果平均值(51.6%~90.8%)[9, 10, 18, 21, 25]比较接近(表 1), 和地上部向地下部转移的碳分配变化规律相同.这是因为小麦生育前期根系碳库活力高, 需转运更多的碳, 而到成熟期则向籽粒中分配较多[10, 18].本研究冬小麦生长前期向地下部分配的碳量多于成熟期, 小麦拔节期向地下部分配了30%~40%的净光合碳, 而成熟期仅有5%~20%[21].分配到地下部光合碳形成根系的比例也与生育时期有关:冬小麦生育前期有近50%光合碳用于根系构建, 但成熟期仅有10%用于根系组织构成, 其余碳通过根系呼吸及根系沉积物损失[2, 18].类似地, 光合碳在土壤中的比例也随生育期呈下降趋势, 但累积量则不断增加[18].
光合碳在地下部的分配因植物类型而异:在其他禾谷类作物, 如玉米[6]、大麦[9]和水稻上[26]随生长进程的延长逐渐向地上部分配, 但牧草上没有类似规律[2, 27].主要原因在于, 农作物经过长期的选择和进化, 地上部收获指数趋于最大化, 进而导致向地下部分配的光合碳比例降低[2, 27].牧草营养生长期长, 收获后地下部根系继续生长, 而禾谷类作物收获后根系逐渐死亡, 造成牧草地下部的光合碳量高于禾谷类作物.禾谷类作物和牧草向地下部转运的光合碳, 分别在20%~30%和30%~50%[2, 21]之间.此外, 根系主动吸收土壤矿质养分是一个能量消耗的过程, 集约化施肥可以显著降低农作物吸收土壤养分的能量消耗, 进而农作物对地下部光合碳投入也相应较低[2].
对于禾谷类作物, 向地下部转运的20%~30%光合碳中, 约1/2在根系, 1/3被根系和微生物呼吸所消耗, 其余1/6被转化为土壤微生物量和有机质[2, 21].这与本研究结果接近(21.5%; 表 1), 即根际呼吸比例最大, 进入土壤有机碳的比例最小.但本研究的示踪期较长(28 d), 更多的根际沉积物被分解和根系释放, 因而根际呼吸比例偏大(58%), 而残留在根系的偏小(28%; 图 4).
3.2 试验方法本研究中(图 5), 在示踪期最后2 d内, 只有很少部分的13C在冬小麦根际呼吸中出现(0.7%~2.7%), 因此, 26 d后根际呼吸产生的13C量微小, 可以忽略, 即使再延长示踪期, 13C在冬小麦-土壤系统中各组分的分配比例也不会产生显著变化, 可以认为经过4周13C的分配已基本完成.关于小麦示踪期的长短, 一般认为15~21 d的示踪期是合适的[18, 28], 其中脉冲标记法光合碳分配主要在48 h内进行[28].利用稳定性碳标记, 不仅可以定量根际沉积物对土壤碳的贡献, 还可以区分不同土壤呼吸, 定量根际释放的碳[22].洗根法低估了植物在生长过程中输入到土壤的碳量[2, 13], 如果利用传统洗根法, 本研究中根际沉积转化的土壤有机碳和根际呼吸就无法定量, 导致进入地下部碳被低估58%.因此, 利用稳定性碳标记, 定量化作物对地下部碳的贡献有着传统方法不具备的优势.
3.3 冬小麦输入到地下的碳量本研究中, 冬小麦输入到地下部(根+土壤有机碳+根际呼吸)总碳量为1.72 t·hm-2, 略高于Kuzyakov等[2]汇总的结果(1.52 t·hm-2).可能原因是, 本研究是基于冬小麦整个生育期, 超过200 d, 而Kuzyakov等[2]的研究有2个研究周期超过200 d, 大部分在100 d左右.幼苗期, 作物根系生物量较小, 输入到地下部的光合碳碳量较少, 远低于后期输入到地下部的数量[2, 17].因此, 在植物-土壤光合碳分配研究中, 基于全生育期的标记和采样是十分必要的[6, 9].
华北平原的农业集约化不仅增加了作物产量, 还提高了土壤有机碳水平[29], 其中秸秆还田是主要原因, 但作物(冬小麦+夏玉米)根际沉积作用的贡献并没有得到准确定量.结合本课题组针对夏玉米利用13CO2脉冲标记的结果[6]发现:在华北平原冬小麦-夏玉米周年生产体系下, 作物生长过程中通过根系沉积物的形式输入碳量达到0.58 t·(hm2·a)-1[冬小麦0.25 t·(hm2·a)-1+夏玉米0.33 t·(hm2·a)-1], 这些根际沉积碳最终成为土壤有机碳的一部分, 参与到土壤生物活动[24].该结果是对当前我国华北平原生产水平下作物根际-土壤碳周转的首次报道, 对于该地区农田土壤固碳减排、培肥地力等具有重要科学支撑和实践意义.
4 结论(1) 用13C脉冲标记冬小麦后, 经过4周的示踪期, 13C在冬小麦-土壤系统中的分配已基本稳定.
(2) 冬小麦光合固定的13C大部分保留在地上部(51.6%~90.8%), 随生育期的增长, 增加向地上部的分配, 降低向地下部(根系、土壤和根际呼吸)的分配.
(3) 在整个生育期内, 冬小麦净吸收光合碳分配到地上部、根、土壤有机碳和根际呼吸中的比例分别为:78.5%、6.0%、3.1%和12.4%.
(4) 冬小麦在整个生育期内输入到地下部的光合碳量是1.72 t·hm-2, 其中有0.99 t·hm-2被根际呼吸消耗, 根部固持碳量为0.48 t·hm-2, 另有0.25 t·hm-2以土壤沉积的形式进入土壤.
| [1] | Lu C H, Fan L. Winter wheat yield potentials and yield gaps in the North China Plain[J]. Field Crops Research, 2013, 143: 98-105. DOI:10.1016/j.fcr.2012.09.015 |
| [2] | Kuzyakov Y, Domanski G. Carbon input by plants into the soil.Review[J]. Journal of Plant Nutrition and Soil Science, 2000, 163(4): 421-431. DOI:10.1002/(ISSN)1522-2624 |
| [3] | Rasse D P, Rumpel C, Dignac M F. Is soil carbon mostly root carbon? Mechanisms for a specific stabilisation[J]. Plant and Soil, 2005, 269(1-2): 341-356. DOI:10.1007/s11104-004-0907-y |
| [4] | Butterly C R, Armstrong R, Chen D L, et al. Carbon and nitrogen partitioning of wheat and field pea grown with two nitrogen levels under elevated CO2[J]. Plant and Soil, 2015, 391(1-2): 367-382. DOI:10.1007/s11104-015-2441-5 |
| [5] | Chowdhury S, Farrell M, Bolan N. Photoassimilated carbon allocation in a wheat plant-soil system as affected by soil fertility and land-use history[J]. Plant and Soil, 2014, 383(1-2): 173-189. DOI:10.1007/s11104-014-2173-y |
| [6] | Meng F Q, Dungait J A J, Zhang X, et al. Investigation of photosynthate-C allocation 27 days after 13C-pulse labeling of Zea mays L.at different growth stages[J]. Plant and Soil, 2013, 373(1-2): 755-764. DOI:10.1007/s11104-013-1841-7 |
| [7] | Simard S W, Durall D M, Jones M D. Carbon allocation and carbon transfer between Betula papyrifera and Pseudotsuga menziesii seedlings using a 13C pulse-labeling method[J]. Plant and Soil, 1997, 191(1): 41-55. DOI:10.1023/A:1004205727882 |
| [8] | Roper M M, Fillery I R P, Jongepier R, et al. Allocation into soil organic matter fractions of 14 C captured via photosynthesis by two perennial grass pastures[J]. Soil Research, 2013, 51(8): 748-759. DOI:10.1071/SR12375 |
| [9] | Swinnen J, Van Veen J A, Merckx R. Carbon fluxes in the rhizosphere of winter wheat and spring barley with conventional vs integrated farming[J]. Soil Biology and Biochemistry, 1995, 27(6): 811-820. DOI:10.1016/0038-0717(94)00230-X |
| [10] | Keith H, Oades J M, Martin J K. Input of carbon to soil from wheat plants[J]. Soil Biology and Biochemistry, 1986, 18(4): 445-449. DOI:10.1016/0038-0717(86)90051-9 |
| [11] | Swinnen J, Van Veen J A, Merckx R. Rhizosphere carbon fluxes in field-grown spring wheat:model calculations based on 14 C partitioning after pulse-labelling[J]. Soil Biology and Biochemistry, 1994, 26(2): 171-182. DOI:10.1016/0038-0717(94)90160-0 |
| [12] | Warembourg F R, Estelrich H D. Towards a better understanding of carbon flow in the rhizosphere:a time-dependent approach using carbon-14[J]. Biology and Fertility of Soils, 2000, 30(5-6): 528-534. DOI:10.1007/s003740050032 |
| [13] | Kuzyakov Y, Kretzschmar A, Stahr K. Contribution of Lolium perenne rhizodeposition to carbon turnover of pasture soil[J]. Plant and Soil, 1999, 213(1-2): 127-136. |
| [14] |
屈波, 孟凡乔, 齐孟文, 等. 华北高产农田生态系统中冬小麦分蘖期生长与碳截留[J]. 生态学杂志, 2007, 26(1): 25-30. Qu B, Meng F Q, Qi M W, et al. Growth and carbon sequestration of winter wheat at its tillering stage in high yield farm ecosystem of North China[J]. Chinese Journal of Ecology, 2007, 26(1): 25-30. |
| [15] |
齐鑫, 王敬国. 应用13C脉冲标记方法研究不同施氮量对冬小麦净光合碳分配及其向地下输入的影响[J]. 农业环境科学学报, 2008, 27(6): 2524-2530. Qi X, Wang J G. Distribution and translocation of assimilated C pulse-labeled with 13C for winter wheat (Trticum aestivums L.), as affected by nitrogen supply[J]. Journal of Agro-Environment Science, 2008, 27(6): 2524-2530. |
| [16] | Shi Y F, Wu W L, Meng F Q, et al. Integrated management practices significantly affect N2O emissions and wheat-maize production at field scale in the North China Plain[J]. Nutrient Cycling in Agroecosystems, 2013, 95(2): 203-218. DOI:10.1007/s10705-013-9558-9 |
| [17] | Tian J, Pausch J, Fan M S, et al. Allocation and dynamics of assimilated carbon in rice-soil system depending on water management[J]. Plant and Soil, 2013, 363(1-2): 273-285. DOI:10.1007/s11104-012-1327-z |
| [18] | Swinnen J, Van Veen J A, Merckx R. 14 C pulse-labelling of field-grown spring wheat:an evaluation of its use in rhizosphere carbon budget estimations[J]. Soil Biology and Biochemistry, 1994, 26(2): 161-170. DOI:10.1016/0038-0717(94)90159-7 |
| [19] |
石小霞, 赵诣, 张琳, 等. 华北平原不同农田管理措施对于土壤碳库的影响[J]. 环境科学, 2017, 38(1): 301-308. Shi X X, Zhao Y, Zhang L, et al. Effects of different agricultural practices on soil carbon pool in North China Plain[J]. Environmental Science, 2017, 38(1): 301-308. |
| [20] | Werth M, Kuzyakov Y. Root-derived carbon in soil respiration and microbial biomass determined by 14 C and 13C[J]. Soil Biology and Biochemistry, 2008, 40(3): 625-637. DOI:10.1016/j.soilbio.2007.09.022 |
| [21] | Gregory P J, Atwell B J. The fate of carbon in pulse-labelled crops of barley and wheat[J]. Plant and Soil, 1991, 136(2): 205-213. DOI:10.1007/BF02150051 |
| [22] |
何敏毅, 孟凡乔, 史雅娟, 等. 用13C脉冲标记法研究玉米光合碳分配及其向地下的输入[J]. 环境科学, 2008, 29(2): 446-453. He M Y, Meng F Q, Shi Y J, et al. Estimating photosynthesized carbon distribution and inputs into belowground in a maize soil following 13C pulse-labeling[J]. Environmental Science, 2008, 29(2): 446-453. |
| [23] | Jia S L, Wang X B, Yang Y M, et al. Fate of labeled urea-15N as basal and topdressing applications in an irrigated wheat-maize rotation system in North China Plain:Ⅰ winter wheat[J]. Nutrient Cycling in Agroecosystems, 2011, 90(3): 331-346. DOI:10.1007/s10705-011-9433-5 |
| [24] | Jin J, Wang G H, Liu J D, et al. Seasonal allocation of photosynthetically fixed carbon to the soybean-grown Mollisols in Northeast China[J]. Crop and Pasture Science, 2011, 62(7): 563-570. DOI:10.1071/CP11006 |
| [25] | Lodhi A, Sajjad M H, Mahmood A, et al. Photosynthate partitioning in wheat (Triticum aestivum L.) as affected by root-zone salinity and form of N[J]. Pakistan Journal of Botany, 2009, 41(3): 1363-1372. |
| [26] | Lu Y H, Watanabe A, Kimura M. Input and distribution of photosynthesized carbon in a flooded rice soil[J]. Global Biogeochemical Cycles, 2002, 16(4): 1085. |
| [27] | Hafner S, Unteregelsbacher S, Seeber E, et al. Effect of grazing on carbon stocks and assimilate partitioning in a Tibetan montane pasture revealed by 13CO2 pulse labeling[J]. Global Change Biology, 2012, 18(2): 528-538. DOI:10.1111/j.1365-2486.2011.02557.x |
| [28] | Derrien D, Marol C, Balesdent J. The dynamics of neutral sugars in the rhizosphere of wheat.An approach by 13C pulse-labelling and GC/C/IRMS[J]. Plant and Soil, 2004, 267(1-2): 243-253. DOI:10.1007/s11104-005-5348-8 |
| [29] | Liao Y, Wu W L, Meng F Q, et al. Increase in soil organic carbon by agricultural intensification in northern China[J]. Biogeosciences, 2015, 12(5): 1403-1413. DOI:10.5194/bg-12-1403-2015 |