 2018, Vol. 39
2018, Vol. 39

2. 江苏省厌氧生物技术重点实验室, 无锡 214122;
3. 无锡国联环保科技股份有限公司, 无锡 214131
 , SHI Wan-sheng1,2 , ZHAO Ming-xing1,2 , XU Zhi-yang1,2 , RUAN Wen-quan1,2
, SHI Wan-sheng1,2 , ZHAO Ming-xing1,2 , XU Zhi-yang1,2 , RUAN Wen-quan1,2 , SONG Lian3 , ZHU Ge3
, SONG Lian3 , ZHU Ge3
2. Jiangsu Key Laboratory of Anaerobic Biotechnology, Wuxi 214122, China;
3. Wuxi Guolian Environmental Science and Technology Co., Ltd., Wuxi 214131, China
目前, 我国每年产生湿污泥(以TS为20%计)约3000万t[1], 污泥的处理与处置成为污水处理厂急需解决的问题.近年来, 高含固污泥厌氧消化技术被认为是一种环境友好且相对可持续的处理方法, 可将污泥中大量的有机物转化为沼气, 实现污泥减量化和资源化, 也便于污泥的运输和集中处理, 同时克服了传统厌氧消化处理过程中反应器占地面积大[2], 建设、运行成本高等缺陷而更具有经济性[3].
但研究发现, 在高含固条件下, 污泥的厌氧消化效率往往较低. Fujishima等[4]的研究结果表明, 当TS从3%升高至11%时, 挥发性固体(volatile solid, VS)的去除率由45.6%降至33.8%.其中一个原因就是污泥蛋白质的分解率较低.据Yang等报道[5], 在污泥TS为28 767 mg·L-1下, 蛋白质的分解率只有32.2%.同时, 蛋白质的水解产生大量的氨氮并累积在系统中, 当氨氮浓度达到一定值时就会对整个厌氧消化系统内的微生物生长产生抑制作用[6].其中, 以游离氨(free ammonia nitrogen, FAN)的抑制尤为明显[7]. FAN浓度过高时会影响污泥厌氧消化系统的稳定性[8], 甚至导致系统的崩溃.因此, 研究高含固条件下蛋白质的转化规律对污泥厌氧消化系统的稳定性和高效性至关重要.
基于此, 本研究采用剩余污泥在TS为12%的条件下进行厌氧消化处理.通过对污泥蛋白质的转化产物进行分析, 阐明厌氧消化过程中污泥蛋白质的转化规律, 揭示高含固污泥厌氧消化过程中蛋白质分解率较低的原因, 以期为提高高含固污泥厌氧消化效率奠定基础.
1 材料与方法 1.1 实验内容实验所采用的剩余污泥取自无锡市某污水处理厂, 接种污泥为经中温厌氧消化后的消化污泥.污泥特性见表 1.实验装置采用Bioprocess全自动甲烷潜力测试系统; 控制反应温度为37℃, 甲烷体积由AMPTS v5.0软件统计.实验采用序批式发酵法, 在1 L培养瓶中加入剩余污泥297.5 g, 接种污泥52.5 g和蒸馏水179 g, 使最终TS为12%.向培养瓶中通入5 min氮气以保证厌氧状态后进行厌氧消化, 直到甲烷产量不再变化时停止实验.实验设置3组平行.
|
|
表 1 污泥特性 Table 1 Characteristics of sludge used in this study |
1.2 指标分析方法
pH采用pH计(FE28K, METTLER, Switzerland)测定. TAN测定采用纳氏试剂光度法[9].污泥胞外聚合物(extracellular polymeric substances, EPS)分为外层溶解性胞外聚合物(soluble EPS, SEPS)和内层固着性胞外聚合物(bound EPS, BEPS), 采用加热法提取[10], 分别获得溶解性蛋白质(SEPS蛋白质)和结合态蛋白质(BEPS蛋白质), 采用改进的Lowry法分析[11].污泥总蛋白质质量分数通过测定污泥凯氏氮减去TAN后乘以6.25计算得到[12]. FAN质量浓度通过TAN、pH和温度之间的关系计算得到[13], 计算公式如下:
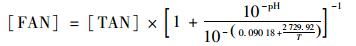
|
式中, [FAN]和[TAN]分别为FAN和TAN的质量浓度(mg·L-1); T为热力学温度(K).
1.3 三维荧光光谱分析采用F-7000型荧光光度计(HITACH, Japan)对厌氧消化前后污泥组分的特性进行分析.三维荧光光谱的激发波长和发射波长分别为200~400 nm和250~500 nm, 激发和发射狭缝均为5.0 nm.
1.4 蛋白质的二维电泳及质谱分析采用土壤蛋白提取试剂盒(MOBIO, United States)提取污泥蛋白, 并通过非干扰蛋白测定试剂盒(生工, 中国)测量蛋白质质量浓度.取500 μg蛋白质提取物进行2-DE.第一向等电聚焦使用IPGphor进行固相pH梯度胶条(17 cm, pH梯度为3.0~10.0)的水化和上样.每个胶条在10 mL的平衡缓冲液[0.05 mol·L-1三(羟甲基)氨基甲烷, pH 8.8, 6 mol·L-1尿素, 30%(质量分数)甘油和2%(质量分数)十二烷基硫酸盐(sodium dodecyl sulfate, SDS)]中进行两次胶条平衡(第一次添加0.002%溴酚蓝和0.125 mol·L-1二硫苏糖醇, 第二次添加0.002%溴酚蓝和0.125 mol·L-1碘乙酰胺).然后在Multiphor Ⅱ系统中进行第二向SDS电泳[14]. 2-DE结束, 用考马斯亮蓝R250对凝胶染色后进行下一步分析.实验中通过手动目测分析差异蛋白点.
从聚丙烯酰胺凝胶上切下待分析的蛋白点, 进行酶解[15].将酶解得到的干粉重新溶解于5 μL含0.1%三氟乙酸(trifluoroacetic acid, TFA)的溶液中, 然后按照1:1的比例与含50%乙腈和1%TFA的α-氰基-4-羟基肉桂酸饱和溶液混合后取1 μL样品进行质谱点靶鉴定, 仪器为Applied Biosystems公司ABI5800串联飞行时间质谱仪.采用正离子模式和自动获取数据的模式进行数据采集; 肽指纹图谱法的质谱扫描范围为800~3500;选择强度最大的10个峰进行二级质谱.将一级和二级质谱数据整合并使用Applied Biosystems公司GPS 3.6和Matrix Science的Mascot 2.3对质谱数据进行分析和蛋白鉴定.
2 结果与讨论 2.1 厌氧消化过程中污泥蛋白质的生物转化效率图 1为厌氧消化过程中污泥总蛋白质质量分数随消化时间的变化曲线.初始时, 污泥总蛋白质质量分数为23.71%;消化前期, 随着易降解有机物的快速分解, 污泥中蛋白质分解为小分子有机物和氨氮, 蛋白质质量分数大幅降低.到33 d后, 蛋白质的含量趋近于稳定; 到第45 d消化结束时, 污泥中总蛋白质的质量分数降低到19.24%;考虑到污泥的TS由12.0%降低到9.72%, 此时, 污泥总蛋白质质量分数降低了34.26%.

|
图 1 总蛋白质质量分数的变化曲线 Fig. 1 Variation of total protein in the sludge during the operation |
污泥中蛋白质在厌氧过程中首先进行水解成小分子和氨氮, 在厌氧消化前15 d, TAN质量浓度从初始的55 mg·L-1迅速上升至752 mg·L-1[图 2(a)]; 然后, TAN质量浓度上升速率有所放缓, 直到第45 d消化结束时, 污泥中TAN质量浓度达到最大值.由于氮素是厌氧微生物生长所必须的营养元素[16], 一般认为, TAN质量浓度低于200 mg·L-1时, 对厌氧消化过程是有利的[17].何仕均等的研究发现[18], 当TAN质量浓度大于800 mg·L-1时开始表现抑制产甲烷作用, 抑制程度为7%, 且抑制程度与TAN质量浓度呈正相关.根据实验结果, 可知TAN质量浓度已超过对微生物无影响的阈值, 因此这将可能导致对厌氧消化产生抑制.由于TAN中的FAN具有细胞膜渗透性[19], 可通过被动扩散进入细胞内, 破坏质子平衡, 导致K+流失, 从而对产甲烷微生物体系产生毒性[20], 本实验同时对厌氧消化过程中的pH和FAN进行了监测, 其变化如图 2(b)和图 2(c)所示.从图 2中可知, 在反应的最初9 d内, pH和FAN均上升较快, 这与微生物分解含氮化合物产生大量氨氮导致pH和FAN升高所对应.第9~12 d时, pH和FAN均有一定幅度的下降, 而在第18 d后, TAN质量浓度达到821 mg·L-1, 可能对微生物活性产生抑制而减缓了TAN的产生.第12~39 d, pH一直在7.7~7.9之间波动; FAN则在第12~30 d内于80 mg·L-1以下波动.之后, pH与FAN均呈现出上升趋势, FAN的质量浓度逐渐对产甲烷微生物体系产生抑制作用.

|
图 2 TAN质量浓度、pH与FAN质量浓度的变化曲线 Fig. 2 Variation of TAN, pH, and FAN in the sludge during the operation |
消化实验结束时, 最终TAN质量浓度、pH和FAN质量浓度分别上升至1 201 mg·L-1、8.12和154 mg·L-1.因此, 在12%含固率条件下, 厌氧消化过程中产生的TAN和FAN已达到较高质量浓度, 对蛋白质的生物分解产生了一定的抑制作用.此外, 高含固率导致的传质困难也可能影响到蛋白质的分解.而这种较低的蛋白质转化效率可能是导致污泥厌氧消化中有机物转化效率不高的一个重要原因.
2.2 厌氧消化过程中污泥EPS蛋白质形态的变化图 3为污泥不同结合状态的EPS蛋白质在厌氧消化过程中的变化情况.厌氧消化开始时, 溶解态蛋白质(SEPS蛋白质)质量浓度较低, 为774 mg·L-1, 结合态蛋白质(BEPS蛋白质)质量分数较高, 占污泥干重的2.99%.随着厌氧消化的进行, SEPS蛋白质质量浓度随时间不断波动, 呈先降低后升高的趋势, 在第18 d达到最低, 说明污泥在厌氧消化过程中部分微生物细胞破壁, 胞内物质流出[21], 同时又在不停地被分解.随后, SEPS蛋白质质量浓度大幅上升, 在第45 d达到1 223 mg·L-1, 并留在污泥液相中难以分解.

|
图 3 SEPS蛋白质质量浓度与BEPS蛋白质质量分数的变化曲线 Fig. 3 Variation of SEPS protein and BEPS protein in the sludge during the operation |
污泥BEPS蛋白质质量分数在消化前9 d大幅下降, 在实验第9 d降至最低.随后, BEPS蛋白质质量分数开始在一定范围内波动, 说明胞外聚合物中结合态蛋白质的形态转化与分解作用主要发生在前9 d, 之后蛋白质从结合态向溶解态的转化速度减缓, 到第15 d, 结合态蛋白质已基本被分解, 污泥中剩余某些难消化或不能被消化的蛋白质使得BEPS蛋白质质量分数趋于稳定, 反应后期稳定在0.94~1.68%.因此, 污泥EPS中蛋白质在厌氧消化过程中不断由结合态向溶解态转化, 并进行微生物转化, 部分难以被微生物分解的蛋白质残留在污泥中.
2.3 厌氧消化过程中污泥组分的3D-EEM分析为了明确厌氧消化中污泥蛋白质的转化过程和转化产物, 采用3D-EEM对厌氧消化前后污泥组分的变化进行分析.如图 4所示, 峰a为酪氨酸蛋白类物质, 峰b为色氨酸蛋白类物质, 峰c为溶解性微生物代谢产物, 峰d为腐殖酸类物质, 峰e为富里酸类物质[22]. 图 4(a)、图 4(b)分别为污泥厌氧消化前后SEPS中有机物的变化情况, 从中可以看出, 厌氧消化反应前后污泥液相中有机物的变化较大; 除峰a的峰强减弱外, 峰b、峰c的峰强均大幅增强, 同时形成了峰d、峰e, 说明除酪氨酸蛋白类物质浓度有所下降外, 色氨酸蛋白类物质、溶解性微生物代谢产物、腐殖酸类及富里酸类物质浓度均有上升.原因可能是由于厌氧消化的水解作用, 污泥中EPS在消化过程中被分解, 使得污泥絮体解体[23], 污泥细胞溶胞释放出胞内有机物, 大分子有机物溶出或分解为小分子有机物有利于后面微生物利用[24].其中溶解性微生物代谢产物占了有机物的主要部分, 同时形成了较多的色氨酸蛋白类、腐殖酸类和富里酸类物质.

|
图 4 厌氧消化前后EPS的3D-EEM图谱 Fig. 4 3D-EEM spectra of EPS before and after anaerobic digestion |
图 4(c)、图 4(d)分别为污泥厌氧消化前后BEPS中有机物的变化情况, 从中可以看出, 污泥厌氧消化反应后, 峰a、b、c的强度均有不同程度的减弱, 峰d的强度稍有增强, 同时形成了峰e, 即酪氨酸、色氨酸蛋白类物质及溶解性微生物代谢产物浓度有一定幅度的下降, 而腐殖酸类及富里酸类物质浓度有所上升.说明污泥经过厌氧消化后大量有机物发生了分解或溶出到液相中, 导致酪氨酸蛋白类、色氨酸蛋白类物质及溶解性微生物代谢产物在BEPS中大大降低, 而腐殖酸类、富里酸类物质在SEPS与BEPS中均有大幅增加, 说明经过厌氧消化, 污泥中有大量腐殖酸类、富里酸类物质生成, 这些物质不仅更难分解, 而且对污泥的厌氧消化过程产生一定的抑制作用[25].
2.4 2-DE及MS分析厌氧消化前后污泥中蛋白质的变化为了深入了解厌氧消化前后污泥中蛋白质组分的变化情况, 本研究采用2-DE对污泥厌氧消化前后蛋白质组分进行分离, 然后对分离的蛋白质进行MS分析.污泥蛋白质的2-DE结果如图 5所示.从中可以发现污泥中的蛋白质经过消化后呈现显著差异, 部分蛋白质被分解.从pI梯度方向蛋白质分布变化可以发现, 厌氧消化后污泥中蛋白质的pI降低.从相对分子质量分布来看, 消化前污泥中的蛋白质主要分布在18 400~45 000, 消化后的污泥蛋白质主要集中在14 400~18 400和25 000~40 000.这意味着经过厌氧消化后, 蛋白质的相对分子质量下降, 趋向小分子化.

|
图 5 污泥厌氧消化前后的2-DE图 Fig. 5 2-DE of protein before and after anaerobic digestion |
为了进一步了解厌氧消化前后污泥蛋白质的性质, 将凝胶上的蛋白质点经过手动切割后进行MS分析, 结果如表 2所示.从中可以看出, 污泥中大部分的蛋白质来自于微生物.其中剩余污泥中的蛋白质主要来自于活性污泥微生物, 例如硝化细菌(Nitrospirae bacterium)、变形菌(Burkholderiales bacterium、Rhodocyclales bacterium、Gallionellales bacterium、Gilliamella apicola)、除磷细菌(Candidatus Accumulibacter aalborgensis)等体内的蛋白质, 还有部分来自人体的蛋白质, 例如血清蛋白(serum albumin)等.消化后污泥中的蛋白质主要来自于经过厌氧消化产生的大量厌氧微生物, 例如产甲烷菌(Methanosaeta concilii、Methanoculleus marisnigri)等, 而同时也发现如血清蛋白(serum albumin)等来自人体的蛋白质并未被分解, 残留了下来.实验结果表明, 污泥中存在的大部分蛋白质为微生物体内的酶等蛋白质, 而随着厌氧消化的进行, 底物被逐渐消耗, 微生物代谢及分泌酶的能力降低[26]; 同时, 有害中间产物氨氮和腐殖酸类物质的产生会进一步抑制蛋白酶的活性[27], 最终微生物无法继续利用该部分蛋白质或厌氧消化体系中不具备分解这些蛋白质的酶而导致污泥蛋白质分解的停滞, 使得这些蛋白质残留在污泥中而难以继续分解, 从而制约了污泥中蛋白质的分解效率.
|
|
表 2 污泥中蛋白质的鉴定 Table 2 List of identified proteins in the sludge |
3 结论
(1) 在TS为12%条件下的污泥厌氧消化过程中, 污泥蛋白质的存在形态发生了转变, 逐渐由结合态向溶解态转化, 表现为污泥絮体解体, 细胞破壁, 蛋白质流出后再被分解的过程.同时形成了大量氨氮, 导致的高氨氮浓度对厌氧消化效率, 尤其对蛋白质的分解表现出一定的抑制作用.
(2) 污泥中的部分蛋白质向腐殖质类、富里酸类物质转化而腐殖化, 变得更加难分解.
(3) 总体来看, 经过厌氧消化后污泥中蛋白质的相对分子质量降低, 污泥蛋白质由大分子向小分子转化.由于污泥中残留的大部分蛋白质来源于微生物体内; 而随着厌氧消化过程的进行, 微生物代谢能力及酶活性随之降低, 或厌氧消化体系中不具备分解这些蛋白质的酶, 导致微生物最终无法继续利用该部分蛋白质, 从而限制了污泥中蛋白质的分解效率.
| [1] | Duan N N, Dong B, Wu B, et al. High-solid anaerobic digestion of sewage sludge under mesophilic conditions:feasibility study[J]. Bioresource Technology, 2012, 104: 150-156. DOI:10.1016/j.biortech.2011.10.090 |
| [2] | Hidaka T, Wang F, Togari T, et al. Comparative performance of mesophilic and thermophilic anaerobic digestion for high-solid sewage sludge[J]. Bioresource Technology, 2013, 149: 177-183. DOI:10.1016/j.biortech.2013.09.033 |
| [3] | Devlin D C, Esteves S R R, Dinsdale R M, et al. The effect of acid pretreatment on the anaerobic digestion and dewatering of waste activated sludge[J]. Bioresource Technology, 2011, 102(5): 4076-4082. DOI:10.1016/j.biortech.2010.12.043 |
| [4] | Fujishima S, Miyahara T, Noike T. Effect of moisture content on anaerobic digestion of dewatered sludge:ammonia inhibition to carbohydrate removal and methane production[J]. Water Science and Technology:A Journal of the International Association on Water Pollution Research, 2000, 41(3): 119-127. DOI:10.2166/wst.2000.0063 |
| [5] | Yang G, Zhang P Y, Zhang G M, et al. Degradation properties of protein and carbohydrate during sludge anaerobic digestion[J]. Bioresource Technology, 2015, 192: 126-130. DOI:10.1016/j.biortech.2015.05.076 |
| [6] | Rajagopal R, Massé D I, Singh G. A critical review on inhibition of anaerobic digestion process by excess ammonia[J]. Bioresource Technology, 2013, 143: 632-641. DOI:10.1016/j.biortech.2013.06.030 |
| [7] | Li N, Xue Y G, Chen S S, et al. Methanogenic population dynamics regulated by bacterial community responses to protein-rich organic wastes in a high solid anaerobic digester[J]. Chemical Engineering Journal, 2017, 317: 444-453. DOI:10.1016/j.cej.2017.02.098 |
| [8] |
陈思思, 戴晓虎, 薛勇刚, 等. 影响高含固厌氧消化性能的主要因素研究进展[J]. 化工进展, 2015, 34(3): 831-839, 856. Chen S S, Dai X H, Xue Y G, et al. Research progress of the main factors influencing properties of high-solid anaerobic digestion[J]. Chemical Industry and Engineering Progress, 2015, 34(3): 831-839, 856. |
| [9] | 国家环境保护总局. 水和废水监测分析方法[M]. 第四版. 北京: 中国环境科学出版社, 2002: 279-281. |
| [10] | Li X F, Zhang L N, Du G C. Influence of sludge discharge on sludge settleability and membrane flux in a membrane bioreactor[J]. Environmental Technology, 2010, 31(11): 1289-1294. DOI:10.1080/09593331003713669 |
| [11] | Lowry O H, Rosebrough N J, Farr A L, et al. Protein measurement with the Folin phenol reagent[J]. The Journal of Biological Chemistry, 1951, 193(1): 265-275. |
| [12] | Miron Y, Zeeman G, Van Lier J B, et al. The role of sludge retention time in the hydrolysis and acidification of lipids, carbohydrates and proteins during digestion of primary sludge in CSTR systems[J]. Water Research, 2000, 34(5): 1705-1713. DOI:10.1016/S0043-1354(99)00280-8 |
| [13] | Hansen K H, Angelidaki I, Ahring B K. Anaerobic digestion of swine manure:inhibition by Ammonia[J]. Water Research, 1998, 32(1): 5-12. DOI:10.1016/S0043-1354(97)00201-7 |
| [14] | Laemmli U K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4[J]. Nature, 1970, 227(5259): 680-685. DOI:10.1038/227680a0 |
| [15] | Katayama H, Nagasu T, Oda Y. Improvement of in-gel digestion protocol for peptide mass fingerprinting by matrix-assisted laser desorption/ionization time-of-flight mass spectrometry[J]. Rapid Communications in Mass Spectrometry, 2001, 15(16): 1416-1421. DOI:10.1002/(ISSN)1097-0231 |
| [16] | Wang J L, Yan D L, Dixon R, et al. Deciphering the principles of bacterial nitrogen dietary preferences:a strategy for nutrient containment[J]. mBio, 2016, 7(4). |
| [17] | Liu T, Sung S. Ammonia inhibition on thermophilic aceticlastic methanogens[J]. Water Science and Technology:A Journal of the International Association on Water Pollution Research, 2002, 45(10): 113-120. DOI:10.2166/wst.2002.0304 |
| [18] |
何仕均, 王建龙, 赵璇. 氨氮对厌氧颗粒污泥产甲烷活性的影响[J]. 清华大学学报(自然科学版), 2005, 45(9): 1294-1296. He S J, Wang J L, Zhao X. Effect of ammonium concentration on the methanogenic activity of anaerobic granular sludge[J]. Journal of Tsinghua University (Science & Technology), 2005, 45(9): 1294-1296. |
| [19] |
戴晓虎, 何进, 严寒, 等. 游离氨调控对污泥高含固厌氧消化反应器性能的影响[J]. 环境科学, 2017, 38(2): 679-687. Dai X H, He J, Yan H, et al. Effects of free ammonia regulation on the performance of high solid anaerobic digesters with dewatered sludge[J]. Environmental Science, 2017, 38(2): 679-687. |
| [20] | Elbeshbishy E, Nakhla G, Hafez H. Biochemical methane potential (BMP) of food waste and primary sludge:influence of inoculum pre-incubation and inoculum source[J]. Bioresource Technology, 2012, 110: 18-25. DOI:10.1016/j.biortech.2012.01.025 |
| [21] | Du H X, Li F S. Characteristics of dissolved organic matter formed in aerobic and anaerobic digestion of excess activated sludge[J]. Chemosphere, 2017, 168: 1022-1031. DOI:10.1016/j.chemosphere.2016.10.108 |
| [22] | Zhang Z S, Guo L, Wang Y, et al. Degradation and transformation of extracellular polymeric substances (EPS) and dissolved organic matters (DOM) during two-stage anaerobic digestion with waste sludge[J]. International Journal of Hydrogen Energy, 2017, 42(15): 9619-9629. DOI:10.1016/j.ijhydene.2017.02.201 |
| [23] |
崔伟莎. 污泥流变特性变化规律及其影响因素的研究[D]. 北京: 北京建筑大学, 2014. 16. Cui W S. Research on the variation regulation of rheological properties of sludge and its influencing factors[D]. Beijing: Beijing University of Civil Engineering and Architecture, 2014. 16. http://cdmd.cnki.com.cn/Article/CDMD-10016-1014332848.htm |
| [24] |
邢立群, 彭永臻, 金宝丹, 等. 盐度强化剩余污泥碱性发酵产酸[J]. 中国环境科学, 2015, 35(6): 1771-1779. Xing L Q, Peng Y Z, Jin B D, et al. Enhanced production of short-chain fatty acids from waste activated sludge alkaline fermentation:the effect of sa-linity[J]. China Environmental Science, 2015, 35(6): 1771-1779. |
| [25] | Fernandes T V, Van Lier J B, Zeeman G. Humic acid-like and fulvic acid-like inhibition on the hydrolysis of cellulose and tributyrin[J]. Bioenergy Research, 2015, 8(2): 821-831. DOI:10.1007/s12155-014-9564-z |
| [26] | Huang Y, Yang L L, Ruan W Q, et al. Evaluation of the hydrolysis performance in the kitchen waste and excess sludge anaerobic co-digestion process[J]. Environmental Technology Reviews, 2015, 4(1): 149-157. DOI:10.1080/09593330.2015.1111429 |
| [27] |
郝晓地, 唐兴, 曹亚莉. 腐殖质对污泥厌氧消化的影响及其屏蔽方法[J]. 环境科学学报, 2017, 37(2): 407-418. Hao X D, Tang X, Cao Y L. Effects of Humic substances on anaerobic digestion of excess sludge and their blocked approaches[J]. Acta Scientiae Circumstantiae, 2017, 37(2): 407-418. |