 2018, Vol. 39
2018, Vol. 39



2. 中国地质科学院岩溶地质研究所, 国土资源部/广西岩溶动力学重点实验室, 桂林 541004;
3. 联合国教科文组织国际岩溶研究中心, 桂林 541004
2. Key Laboratory of Karst Dynamics, Ministry of Land and Resources & Guangxi, Institute of Karst Geology, Chinese Academy of Geological Sciences, Guilin 541004, China;
3. International Research Centre on Karst under the Auspices of UNESCO, Guilin 541004, China
过去通常认为, 水库又称“人工湖泊”或“人工湿地”是大气二氧化碳和甲烷等温室气体的主要排放源[1~4], 其排放强度与水环境、淹没的土壤及植被类型等因素有关[5~7].此外, 水库产生的温室气体是水体中自身或陆源的有机质通过水体微生物分解产生, 并通过气泡排放、植物传输和扩散、水轮机、溢洪道和大坝下游等途径由水体逃逸至大气环境[8].耿雪萌[9]利用Wetland-DNDC模型得出密云水库消落带CH4总排放量为15.1 g·m-2, 而Bevelhimer等[10]在调查美国东南部的水库时发现:水库CO2释放速率为994~2 760 mg·(m2·d)-1, CH4释放速率为6~187 mg·(m2·d)-1.此外, Deemer等[11]通过计算发现全球通过水库年释放的温室气体高达0.8 Pg (0.5~1.2 Pg).然而最新研究结果表明:河流、湖泊和水库具有高度的环境异质性和微生物群落多样性, 是碳氮转化的主要场所, 固定的有机碳相当于海洋固碳总量的近40%[12].此外, 总面积约为1.6×106 km2的非碳酸盐型淡水水域对溶解无机碳的滞留率仅为5 g·(m2·a)-1, 而总面积约为0.18×106 km2的碳酸盐型淡水水域对溶解无机碳的平均滞留率为100 g·(m2·a)-1[13].尽管宋昂等[14]、Li等[15]以及卢晓漩等[16]认为微型光合生物(光合细菌等)对岩溶水体有机碳的固定具有重要的贡献, 然而关于微型光合生物对岩溶水体二氧化碳和甲烷释放速率的影响至今鲜见报道.
为了解岩溶区水库微型光合生物对岩溶水体CO2和CH4排放速率的影响, 本文选择桂林五里峡水库作为研究对象, 设置消落带、中游、交汇处、坝前、坝尾等5个采样点, 运用静态箱法, 结合微型光合生物初级生产力, 探讨岩溶区水库微型光合生物与岩溶水体CO2和CH4排放速率的关系, 以期为进一步评估微型生物对岩溶水体碳循环的贡献提供了理论依据.
1 材料与方法 1.1 研究区概况五里峡水库位于广西桂林市兴安县漠川河上游, 库区位于25°30′06″~25°35′31″N, 110°44′41″~110°47′12″E之间.五里峡水库建于1972年, 主要由湘江支流漠川河补给, 库容约1.08×109 m3, 是一座以灌溉为主, 兼有发电、防洪、漓江补水等综合功能的大型水库.五里峡水库受亚热带季风气候影响, 多年平均气温19.0℃, 12月平均气温8.79℃, 多年平均降水量1 887.1 mm[17], 12月占全年3.3%.水库中、上游库区分布于岩溶区, 出露的地层主要为中泥盆统东岗岭阶(D2d)碳酸盐岩; 下游及大坝地区出露的地层主要为中泥盆统信都组(D2x、D2d)砂岩夹页岩地层, 岩溶区面积占水库库区总面积的45%, 主要岩溶地貌为峰丛谷地, 为地表水补给型半岩溶水库.
1.2 采样与分析2016年在五里峡水库消落带(WLX01)、中游(WLX03)、交汇处(WLX04)、坝前(WLX05)、坝尾(WLX02)共设置5个采样点(见图 1), 使用浮游静态箱(暗箱)采集CO2、CH4气体, 采集的方法是静态箱在下水前, 接箱内通风扇使箱内气体混合均匀, 静态箱下水时先采集一次箱内气体作为背景浓度, 再分别隔5、5、10、15 min采集一次气体, 保存于1 L气样袋中, 密封避光保存, 每个点共5个气袋, 气样袋取回实验室后48 h内用Agilent 7890B气相色谱仪进行CO2、CH4浓度分析.利用YSI EXO水质监测和测量平台(美国YSI公司)现场检测溶解氧(dissolved oxygen, DO)、水温(T)、pH, DIC带回实验室通过TOC-Ⅴ分析仪(日本岛津公司)测定.

|
图 1 五里峡水库采点示意 Fig. 1 Sampling sites in Wulixia reservoir |
为了解水体微型光合生物初级生产力, 按照王桂云等[18]的方法:采集50 mL原水样装入可密封的玻璃瓶中, 共采集2瓶, 其中一瓶水样加入10滴饱和氯化汞, 然后2瓶水样都加100 μL NaH14CO3溶液密封现场悬挂培养48 h, 培养液用0.22 μm微孔滤膜过滤, 最后将过滤膜用1220C超低本底液闪仪测量14C放射性活度.
1.3 温室气体排放速率及初级生产力计算方法水-气界面的CO2和CH4排放速率计算公式[19]见式(1):
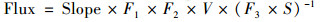
|
(1) |
式中, Slope为时间-浓度关系图中的斜率(×10-6 min-1), F1为分子摩尔质量(CO2为44 g·mol-1; CH4为6 g·mol-1), F2为min和h的转换系数(60), V为浮游箱内套入的空气体积(m3), F3为μg和mg的转换系数(1 000), S为水上浮游箱的表面积(m2), Flux为温室体排放速率[mg·(m2·h)-1].
初级生产力计算公式见式(2):
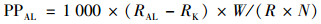
|
(2) |
式中, PPAL为光照下水体固碳速率μg·(L·h)-1, RAL为不过3 μm膜水样14C放射性活度(Bq), RK为加氯化汞水样有机14C放射性活度(Bq), W为DIC浓度(mg·L-1), R为原液有机14C的放射性活度(Bq), N为时间(h).
1.4 数据处理用Origin 8.5对水-气界面的CO2和CH4排放速率进行作图分析; 利用SPSS 19.0进行相关分析(双尾).
2 结果与分析 2.1 五里峡水库二氧化碳和甲烷排放速率五里峡水库温室气体排放速率结果如图 2所示.五里峡水库二氧化碳排放速率变化明显, 而甲烷排放速率变化不大. 2016年7月库区甲烷的波动范围最为明显, 其波动区间为0.08~1.03 mg·(m2·h)-1, 库区二氧化碳(除WLX05点)排放速率均为负值, 说明库区向大气吸收二氧化碳. 2016年10月库区二氧化碳的波动范围为20.65~115.06 mg·(m2·h)-1, 其中WLX05点二氧化碳排放速率达到最大值为115.06 mg·(m2·h)-1; 2016年12月, 五里峡水库二氧化碳的波动范围为-13.00~47.47 mg·(m2·h)-1, 甲烷的波动范围为-0.03~0.07 mg·(m2·h)-1.对比5个采样点二氧化碳平均排放速率, WLX01的平均排放速率最低为14.92 mg·(m2·h)-1, 而WLX05的平均排放速率最高为72.26 mg·(m2·h)-1, 因此五里峡水库温室气体排放速率在空间上总体呈现在库区较高, 而消落带(WLX01)、坝尾(WLX02)较低的格局.

|
图 2 五里峡水库CO2和CH4排放速率 Fig. 2 Exchange flux rate of CO2 and CH4 in Wulixia reservoir |
为了解环境因子对五里峡水库温室气体排放速率的影响, 还检测了水库表层微型光合生物初级生产力、水温、DIC和pH等指标(表 1).结果表明:五里峡水库表层水体温度变化范围为13.66~30.98℃, 并在12月表层水温最低, 7月表层水温最高.研究区水体受碳酸盐岩影响, 水体pH值介于7.19~9.52之间, 平均值为8.21, 结果与文献[14, 16]一致.同时, DIC浓度从7~12月逐渐递增; 而WLX05点的DIC浓度高于其它点.库区微型光合生物的初级生产力在7月最高, 平均值达到2.78 μg·(L·h)-1; 在10月最低, 平均值只有0.03 μg·(L·h)-1.受气体因素、水环境因素和陆地环境因素的共同影响[20], 库区在10月初级生产力最小, 而12月CO2和CH4通量最低.
|
|
表 1 五里峡水库水体理化性质 Table 1 Physical and chemical characteristics of water samples in Wulixia reservoir |
3 讨论 3.1 温室气体排放速率的时空变化
水库水体以溶解和颗粒形态存在的有机碳和无机碳是水气界面二氧化碳排放的物质基础, 其中溶解无机碳以CO2、H2CO3、HCO3-和CO32-等形式为主并影响温室气体排放速率.五里峡水库二氧化碳和甲烷排放速率在时间尺度上存在显著差异.五里峡水库库区7月二氧化碳(除WLX05点)排放速率均为负值, 说明夏季较高的水温(表 1)有利于微型光合生物的生长, 从而将水体中的溶解无机碳转变成有机碳, 进而减少水气界面二氧化碳排放速率[21, 22].五里峡水库二氧化碳排放速率9月最高, 溶解氧平均值为6.514 mg·L-1, 远低于其它4个月的均值, 这是因为水体微型光合生物经过7月旺盛生长的对数期和稳定的平台期后进入衰老期[23].因此, 在9月五里峡水库部分微型光合生物的死亡不但导致水体溶解氧急剧降低, 而且还在细菌作用下将水体中的有机碳转变成溶解无机碳[24]并释放到大气中.此外, 夏季适宜的水温不但能使微型光合生物进入旺盛生长的对数期和稳定的平台期, 而且还能使产甲烷菌进入旺盛生长的对数期和稳定的平台期, 进而导致五里峡水库库区甲烷的排放速率在7月达到最大值, 随后产甲烷菌进入衰老期和停滞期并在12月达到最小值[25].从而使五里峡水库在夏季具有一定的CO2净汇效应, 同时促进CH4的排放, 并在冬季是CO2的排放源和减缓CH4的排放.
此外, 五里峡水库库区断面温室气体排放速率在空间上存在显著的差异. WLX01点在库区的消落带, 二氧化碳排放速率变化范围是-9.19~33.96 mg·(m2·h), -1甲烷排放速率变化范围是0.01~0.29 mg·(m2·h)-1; WLX02点位于坝尾, 该断面二氧化碳排放速率变化范围-3.47~82.72 mg·(m2·h)-1, 甲烷排放速率变化范围是0.01~0.38 mg·(m2·h)-1, 并且较其它断面变化速率较小.这是因为周期性蓄水和排水不但产生了较大面积的消落带影响流域内部陆地碳输入时空变化格局和水汽碳交换的速度和方式[26, 27], 而且还影响坝尾溢洪道流量, 进而扰动CO2和CH4排放速率.作为水体碳积累作用较为典型的坝前断面(WLX05), 其二氧化碳排放速率并没有表现为显著的碳汇, 一定程度上表现为碳源.
3.2 影响温室气体排放速率的因素初级生产力是指绿色植物利用太阳能进行光合作用, 把无机碳固定并转变成有机碳的能力[28].因此, 水体微型光合生物通过光合作用将溶解无机碳转变成有机碳, 进而影响温室气体排放速率.其初级生产力水平与光强、水温、透明度等环境因子有关[29, 30].本研究结果显示(表 2):五里峡水库水体CO2排放速率与初级生产力和pH呈显著负相关(r为-0.477和-0.520), 并与DIC呈弱负相关(r=-0.051).尽管水温不是影响水体微生物生长的关键因素[31], 但在微型光合生物生长所需要的最佳温度范围内, 水温的上升不但能够促使微型光合生物快速生长, 而且还能使其细胞中的生物化学反应速率加快[32], 进而将水体中更多的溶解无机碳固定下来转变成有机碳[33].此外, 五里峡水库位于碳酸盐岩区, 水体中的主要阴离子为HCO3-.岩溶水体的pH值主要取决于水中碳酸体系的平衡移动, 水温的升高将使原碳酸体系的平衡遭到破坏(r=-0.522), 使水的pH值迅速增高(r=0.494)[34].由于五里峡水库水体微型光合生物的快速生长和水中足够多的光合底物(溶解无机碳), 因此五里峡水库水体的微型光合生物通过形成初级生产力将溶解无机碳固定下来转变成有机碳.从而说明微型光合生物的初级生产力是影响五里峡水库温室气体排放速率的重要因素(r=-0.477).该结果与宋昂等[14]认为好氧不产氧光合细菌是五里峡水库有机碳的重要贡献者, 以及五里峡水库水体DOM整体呈现弱腐殖质化和自生源特征的结论[15]一致.从而说明水生微型生物的光合作用在岩溶碳固定过程中扮演着重要的角色.
此外, 五里峡水库水体CH4排放速率与微型光合生物的初级生产力呈显著正相关(r=0.771).微型光合生物形成的有机碳除一部分被暗呼吸利用外, 大部分输送到水库底部[35].由于水库底部的有机碳在厌氧环境下能够被产甲烷菌转化成甲烷和二氧化碳[36], 因此五里峡水库微型光合生物的初级生产力与CH4排放速率显著正相关.该结果与Wik等[37]关于夏季湖泊和水库升温能增加甲烷释放量结论相一致, 因为水温升高能增加微型光合生物的丰度, 提高水体的初级生产力.五里峡水库水体微型光合生物通过形成初级生产力在减缓水气界面二氧化碳排放速率的同时, 还使水库底部埋藏的有机碳含量增加.鉴于部分有机碳重新氧化成甲烷, 因此在五里峡水库水体微型光合生物的作用下水气界面的二氧化碳排放速率与甲烷排放速率显著负相关(r=-0.466).
综上所述, 五里峡水库水体微型光合生物通过形成初级生产力改变了水气界面的二氧化碳排放速率与甲烷排放速率.因此在不造成水库污染的前提下, 通过增加微型光合生物的初级生产力, 促进有机质的储存和掩埋, 进而减少返回大气的CO2通量.
|
|
表 2 温室气体排放速率与环境因子的相关性分析 Table 2 Correlation between greenhouse gas efflux and environmental factors |
4 结论
(1) 五里峡水库夏季为大气CO2的汇, 变化范围在-30.14~-3.47 mg·(m2·h)-1, 秋、冬季均为大气CO2的源, 变化范围在15.57~115.06 mg·(m2·h)-1, CO2的“源/汇”关系相当复杂, 而CH4的“源/汇”关系单一, 甲烷排放速率在夏季变化幅度明显, 变化范围在0.08~1.03 mg·(m2·h)-1, 而在秋、冬季变化稳定, 变化范围在-0.07~0.43 mg·(m2·h)-1, 水库为大气CH4的源.同时, 周期性蓄水和排水是影响五里峡水库不同位置CO2和CH4排放速率的重要因素, CO2和CH4排放速率在空间上表现为水库消落带和坝尾较低而库区较高的分布格局.
(2) 受气体因素、水环境因素和陆地环境因素的影响, 五里峡水库微型光合生物初级生产力与CO2排放速率呈显著负相关, 与CH4排放速率呈极显著正相关.因此, CO2和CH4通量与初级生产力的显著相关性说明水生微型生物的光合作用在岩溶碳固定过程中扮演着重要的角色.
(3) 通过综合分析显示:五里峡水库CO2和CH4通量并没有直接受到水温的影响, 而是通过水温影响初级生产力改变了水气界面的二氧化碳排放速率与甲烷排放速率.五里峡水库CO2和CH4通量时空分布格局显著受到pH和初级生产力的影响, 并且水体微型光合生物通过形成初级生产力产生不同强度的温室气体排放.从而为人们进一步评估微型生物对岩溶水体碳循环的贡献提供了理论依据.
| [1] |
程炳红, 郝庆菊, 江长胜. 水库温室气体排放及其影响因素研究进展[J]. 湿地科学, 2012, 10(1): 121-128. Cheng B H, Hao Q J, Jiang C S. Research progress on the emission of greenhouse gases from reservoir and its influence factors[J]. Wetland Science, 2012, 10(1): 121-128. |
| [2] | Yang L, Lu F, Zhou X P, et al. Progress in the studies on the greenhouse gas emissions from reservoirs[J]. Acta Ecologica Sinica, 2014, 34(4): 204-212. DOI:10.1016/j.chnaes.2013.05.011 |
| [3] | Yang M, Grace J, Geng X M, et al. Carbon dioxide emissions from the littoral zone of a Chinese reservoir[J]. Water, 2017, 9(7): 539. DOI:10.3390/w9070539 |
| [4] | Miller B L, Arntzen E V, Goldman A E, et al. Methane ebullition in temperate hydropower reservoirs and implications for US policy on greenhouse gas emissions[J]. Environmental Management, 2017, 60(4): 615-629. DOI:10.1007/s00267-017-0909-1 |
| [5] | Kumar A, Sharma M P. Estimation of green house gas emissions from Koteshwar hydropower reservoir, India[J]. Environmental Monitoring and Assessment, 2017, 189(5): 240. DOI:10.1007/s10661-017-5958-7 |
| [6] | Li Z, Zhang Z Y, Lin C X, et al. Soil-air greenhouse gas fluxes influenced by farming practices in reservoir drawdown area:A case at the Three Gorges Reservoir in China[J]. Journal of Environmental Management, 2016, 181: 64-73. DOI:10.1016/j.jenvman.2016.05.080 |
| [7] |
赵炎, 曾源, 吴炳方, 等. 三峡水库香溪河支流水域温室气体排放通量观测[J]. 水科学进展, 2011, 22(4): 546-553. Zhao Y, Zeng Y, Wu B F, et al. Observation on greenhouse gas emissions from Xiangxi River in Three Gorges Region[J]. Advances in Water Science, 2011, 22(4): 546-553. |
| [8] |
杨平, 仝川. 淡水水生生态系统温室气体排放的主要途径及影响因素研究进展[J]. 生态学报, 2015, 35(20): 6868-6880. Yang P, Tong C. Emission paths and measurement methods for greenhouse gas fluxes from freshwater ecosystems:a review[J]. Acta Ecologica Sinica, 2015, 35(20): 6868-6880. |
| [9] |
耿雪萌, 杨萌, Grace J, 等. Wetland-DNDC模型模拟密云水库消落带甲烷排放[J]. 资源与生态学报, 2016, 7(4): 281-290. Geng X M, Yang M, Grace J, et al. Simulating methane emissions from the Littoral zone of a reservoir by Wetland DNDC model[J]. Journal of Resources and Ecology, 2016, 7(4): 281-290. |
| [10] | Bevelhimer M S, Stewart A J, Fortner A M, et al. CO2 is dominant greenhouse gas emitted from six hydropower reservoirs in southeastern United States during peak summer emissions[J]. Water, 2016, 8(1): 15. DOI:10.3390/w8010015 |
| [11] | Deemer B R, Harrison J A, Li S Y, et al. Greenhouse gas emissions from reservoir water surfaces:A new global synthesis[J]. Bioscience, 2016, 66(11): 949-964. DOI:10.1093/biosci/biw117 |
| [12] |
黄力, 冯雪莲, 杜全生, 等. 水圈微生物重大研究计划:聚焦水圈微生物组研究的核心科学问题[J]. 中国科学院院刊, 2017, 32(3): 266-272. Huang L, Feng X L, Du Q S, et al. Focusing on key scientific issues of microbiome research in hydrosphere:NSFC major research plan for microbes in hydrosphere[J]. Bulletin of Chinese Academy of Sciences, 2017, 32(3): 266-272. |
| [13] | Downing J P, Meybeck M, Orr J C, et al. Land and water interface zones[J]. Water, Air, & Soil Pollution, 1993, 70(1-4): 123-137. |
| [14] |
宋昂, 彭文杰, 何若雪, 等. 好氧不产氧光合细菌反馈作用下的五里峡水库坝前水体化学特征研究[J]. 岩矿测试, 2017, 36(2): 171-179. Song A, Peng W J, He R X, et al. Hydrochemistry characteristics in front of the Wulixia reservoir dam associated with feedback from aerobic anoxygenic phototrophic bacteria[J]. Rock and Mineral Analysis, 2017, 36(2): 171-179. |
| [15] | Li Q, Song A, Peng W, et al. Contribution of aerobic anoxygenic phototrophic bacteria to total organic carbon pool in aquatic system of subtropical karst catchments, Southwest China:evidence from hydrochemical and microbiological study[J]. FEMS Microbiology Ecology, 2017, 93(6): 65. |
| [16] |
卢晓漩, 彭文杰, 李强, 等. 岩溶区水库冬季溶解有机质组成特征及来源:以桂林五里峡水库为例[J]. 环境科学, 2017, 38(10): 4120-4129. Lu X X, Peng W J, Li Q, et al. Distinguishing the properties and sources of the dissolved organic matter in karst reservoir water during winter using three-dimensional fluorescence spectrum technology:A case study in Wulixia reservoir of Guangxi province[J]. Environmental Science, 2017, 38(10): 4120-4129. |
| [17] |
陈超, 周广胜. 1961-2010年桂林气温和地温的变化特征[J]. 生态学报, 2013, 33(7): 2043-2053. Chen C, Zhou G S. Analysis on variation characteristics of air temperature and ground temperature in Guilin from 1961 to 2010[J]. Acta Ecologica Sinica, 2013, 33(7): 2043-2053. |
| [18] |
王桂云, 毛兴华, 吕瑞华, 等. 14 C法测定海洋初级生产力[J]. 黄渤海海洋, 1984, 2(1): 72-76. Wang G Y, Mao X H, Lv R H, et al. The carbon-14 technique for determining maring primary productivity[J]. Journal of Oceanography of Huanghai & Bohai Seas, 1984, 2(1): 72-76. |
| [19] |
彭文杰, 宋昂, 李强, 等. 一种便携式水土两用集气装置[P]. 中国专利: CN201720042679. X, 2017-08-01. |
| [20] |
赵炎, 曾源, 吴炳方, 等. 水库水气界面温室气体通量监测方法综述[J]. 水科学进展, 2011, 22(1): 135-146. Zhao Y, Zeng Y, Wu B F, et al. Review of methods for measuring greenhouse gas flux from the air-water interface of reservoirs[J]. Advances in Water Science, 2011, 22(1): 135-146. |
| [21] | Liu Z H, Zhao M, Sun H L, et al. "Old" carbon entering the South China Sea from the carbonate-rich Pearl River Basin:Coupled action of carbonate weathering and aquatic photosynthesis[J]. Applied Geochemistry, 2017, 78: 96-104. DOI:10.1016/j.apgeochem.2016.12.014 |
| [22] | Yang M X, Liu Z H, Sun H L, et al. Organic carbon source tracing and DIC fertilization effect in the Pearl River:Insights from lipid biomarker and geochemical analysis[J]. Applied Geochemistry, 2016, 73. |
| [23] | Bidle K D, Falkowski P G. Cell death in planktonic, photosynthetic microorganisms[J]. Nature Reviews Microbiology, 2004, 2(8): 643-655. DOI:10.1038/nrmicro956 |
| [24] | Zhu L, Che X, Liu H, et al. Greenhouse gas emissions and comprehensive greenhouse effect potential of Megalobrama amblycephala culture pond ecosystems in a 3-month growing season[J]. Aquaculture International, 2016, 24(4): 893-902. DOI:10.1007/s10499-015-9959-7 |
| [25] | Brooker M R, Bohrer G, Mouser P J. Variations in potential CH4 flux and CO2 respiration from freshwater wetland sediments that differ by microsite location, depth and temperature[J]. Ecological Engineering, 2014, 72: 84-94. DOI:10.1016/j.ecoleng.2014.05.028 |
| [26] |
李哲, 姚骁, 何萍, 等. 三峡水库澎溪河水-气界面CO2、CH4扩散通量昼夜动态初探[J]. 湖泊科学, 2014, 26(4): 576-584. Li Z, Yao X, He P, et al. Diel variations of air-water CO2 and CH4 diffusive fluxes in the Pengxi River, Three Gor-ges Reservoir[J]. Journal of Lake Sciences, 2014, 26(4): 576-584. DOI:10.18307/2014.0412 |
| [27] | Cai Y H, Guo L D, Wang X R, et al. Abundance, stable isotopic composition, and export fluxes of DOC, POC, and DIC from the lower Mississippi River during 2006-2008[J]. Journal of Geophysical Research:Biogeosciences, 2015, 120(11): 2273-2288. DOI:10.1002/2015JG003139 |
| [28] |
张琪, 袁轶君, 米武娟, 等. 三峡水库香溪河初级生产力及其影响因素分析[J]. 湖泊科学, 2015, 27(3): 436-444. Zhang Q, Yuan Y J, Mi W J, et al. Primary production and its influencing factors in Xiangxi River, Three-Gorges Reservoir[J]. Journal of Lake Sciences, 2015, 27(3): 436-444. DOI:10.18307/2015.0310 |
| [29] |
邵晓阳, 徐耀阳, 韩新芹, 等. 香溪河官庄坪库湾叶绿素a及初级生产力的分布特征[J]. 水生生物学报, 2006, 30(1): 95-100. Shao X Y, Xu Y Y, Han X Q, et al. The distribution of chlorophyll a content and primary productivity in Guanzhuangping bay of Xiangxi River[J]. Acta Hydrobiologica Sinica, 2006, 30(1): 95-100. |
| [30] | Xing Y P, Xie P, Yang H, et al. Methane and carbon dioxide fluxes from a shallow hypereutrophic subtropical Lake in China[J]. Atmospheric Environment, 2005, 39(30): 5532-5540. DOI:10.1016/j.atmosenv.2005.06.010 |
| [31] | Amado A M, Meirelles-Pereira F, Vidal L O, et al. Tropical freshwater ecosystems have lower bacterial growth efficiency than temperate ones[J]. Frontiers in Microbiology, 2013, 4: 167. |
| [32] | Aigner S, Holzinger A, Karsten U, et al. The freshwater red alga Batrachospermum turfosum (Florideophyceae) can acclimate to a wide range of light and temperature conditions[J]. European Journal of Phycology, 2017, 52(2): 238-249. DOI:10.1080/09670262.2016.1274430 |
| [33] | Lechleitner F A, Dittmar T, Baldini J U L, et al. Molecular signatures of dissolved organic matter in a tropical karst system[J]. Organic Geochemistry, 2017, 113: 141-149. DOI:10.1016/j.orggeochem.2017.07.015 |
| [34] |
刘再华. 岩溶水文地球化学研究中pH值野外测定的必要性[J]. 中国岩溶, 1990, 9(4): 310-317. Liu Z H. Necessities of measuring pH in situ in the study of karst hydrogeochemistry[J]. Carsologica Sinica, 1990, 9(4): 310-317. |
| [35] |
张风菊, 薛滨, 姚书春, 等. 全新世大暖期中国湖泊碳埋藏速率初步研究[J]. 第四纪研究, 2013, 33(2): 401-402. Zhang F J, Xue B, Yao S C, et al. The organic carbon burial rates in Chinese Lake sediments during holocene megathermal[J]. Quaternary Sciences, 2013, 33(2): 401-402. |
| [36] | Townsend-Small A, Disbennett D, Fernandez J M, et al. Quantifying emissions of methane derived from anaerobic organic matter respiration and natural gas extraction in Lake Erie[J]. Limnology and Oceanography, 2016, 61(S1). |
| [37] | Wik M, Varner R K, Anthony K W, et al. Climate-sensitive northern lakes and ponds are critical components of methane release[J]. Nature Geoscience, 2016, 9(2): 99-105. DOI:10.1038/ngeo2578 |