 2017, Vol. 38
2017, Vol. 38



2. 南京信息工程大学大气物理学院, 南京 210044;
3. 南京信息工程大学江苏省大气环境与装备技术协同创新中心, 南京 210044;
4. 北京市通州区气象局, 北京 101100
2. School of Atmospheric Physics, Nanjing University of Information Science & Technology, Nanjing 210044, China;
3. Jiangsu Collaborative Innovation Center of Atmospheric Environment and Equipment Technology(CICAEET), Nanjing University of Information Science & Technology, Nanjing 210044, China;
4. Tongzhou District Meteorological Bureau of Beijing, Beijing 101100, China
近地层臭氧主要是由氮氧化物(NOx)、一氧化碳(CO)和挥发性有机物(VOCs)等前体物经一系列光化学反应所形成的一种二次污染物.近年来, 由于人类活动的加剧, 导致臭氧浓度从工业化前期的10 nL·L-1上升到20~45 nL·L-1[1].在下一个世纪, 臭氧浓度可能将会继续增加. Meehl等[2]的研究表明, 2015~2050年间全球平均臭氧浓度将增加20%~25%, 到2100年将增加40%~60%.大量研究已经表明, 目前近地层较高的臭氧浓度足以对水稻、大豆、小麦、马铃薯和玉米等作物的产量带来威胁[3, 4].预计到2030年, 小麦产量将下降10.6%~15.6%, 玉米产量下降4.5%~6.3%, 大豆产量下降12.1%~16.4%, 每年的经济损失将达到120~350亿美元[5], 严重影响到全球的粮食安全.
近地层臭氧不仅会降低植物叶面积、加速叶片老化, 还会抑制其光合作用, 导致植物生物量和产量降低[6~9].地表臭氧对植物影响的实验研究仅仅是针对特定的区域, 采用的实验方法大多是控制或者半控制性实验, 人力和物力消耗较大, 而且获得的数据有限.而模型研究则可以克服以上缺陷, 可以用来模拟其影响过程, 较好地应用于地表臭氧污染的区域风险评估.因此, 通过模型预测臭氧浓度变化对植物干物质和产量损失的影响已经成为大气环境领域研究的热点之一.例如, Zhang等[10]的研究表明, 在当前臭氧浓度水平下, 中国东北地区近地层臭氧可能会使大豆产量减少23.4%~30.2%. Zhu等[11]估算了臭氧对鲁西北平原冬小麦产量的损失约为12.9%. Jin等[12]的研究发现, 水稻产量损失与AOT40存在着较好的线性关系, 当AOT40达到12.93 μL·(L·h)-1时, 水稻将减产约10%.郑启伟等认为[13], 当AOT40分别为4.48 μL·(L·h)-1和14.99 μL·(L·h)-1时, 小麦和水稻的单株产量将减产10%. Emberson等[14]基于臭氧通量模型估算了亚洲主要植物的产量损失, 研究发现, 在植物生长季期间平均AOT40为35~75 nL·(L·h)-1时, 小麦、水稻、大豆分别减产5%~48%、3%~47%和10%~65%.
臭氧对植物影响的评估指标和方法, 由最初的臭氧浓度响应模型(如Wellbull方程、线性方程、二次方程和指数方程)和臭氧剂量响应模型(如SUM06、AOT40和W126)[15~17]逐渐发展到臭氧吸收通量响应模型, 该模型既充分考虑了温度、水汽压差、光照等气候因子, 又考虑了植物对环境因子的响应, 有着很好的适用性[9, 18], 用其来评估臭氧对植物干物质和产量的影响更贴切.因此, 开展气孔臭氧吸收通量的研究, 是定量估算臭氧对植物影响的前提.本文将根据长三角地区冬小麦主要生长季期间观测的3 a(2014~2016年)臭氧浓度数据, 分析臭氧浓度、AOT40的长期变化特征, 引进气孔导度模型和通量模型, 计算冬小麦叶片气孔臭氧吸收通量, 并利用前期建立的相关模型评估了臭氧对冬小麦干物质和产量的潜在影响, 以期为我国种植区大气污染物的环境影响评价提供参考, 也为制定防御臭氧对植物伤害的对策提供科学依据.
1 材料与方法 1.1 观测地点臭氧浓度等项目的观测是在南京信息工程大学农业气象试验站(32°14′N, 118°42′E)的冬小麦麦田试验地(海拔约22 m), 该地土壤类型为黄棕壤, 土质细腻而又均匀, 肥力中等, pH值为7.35, 土壤有机质含量为10.35g·kg-1, 全氮含量为0.55 g·kg-1, 全磷含量为0.47 g·kg-1, 全钾含量为0.21g·kg-1, 速效磷含量为4.46 mg·kg-1, 速效钾含量为59.38 mg·kg-1.观测地周围地势平坦, 下垫面植被均为冬小麦扬麦13, 且植被覆盖均匀.冬小麦主要生长季为3~5月.
1.2 仪器和观测臭氧浓度的观测所采用的仪器为EC9810-O3监测仪(澳大利亚Ecotech公司), 该仪器的精度(体积分数)为1×10-9, 每5 min获取一次数据.仪器的检测原理为紫外吸收法, 测量时使用单个玻璃测量池体, 分析仪的微处理器根据朗伯比尔定律精确地计算出臭氧的浓度.
常规气象数据是通过安装在观测点的WatchDog2000系列自动气象站所获取, 包括温度、湿度、太阳辐射、风速、风向等, 每小时获取一个数据.
同时, 选择晴朗无云的天气, 每个生育期利用美国Decagon公司生产的SC-1稳态气孔计对冬小麦叶片气孔导度进行测定.每次所观测的时间为08:00~17:00, 根据实际天气状况对测量时间和周期进行调整.测量时每隔2 h选择5~6株标记过的冬小麦进行测定, 并分别计算每2 h的平均值.拔节期和孕穗期选择冬小麦的倒二叶, 扬花期和灌浆期选择冬小麦的旗叶进行测定.所观测的时间为2014~2016年每年的3~5月.
1.3 评估指标和模型介绍 1.3.1 AOT40的计算AOT40可表示高浓度臭氧在一定的时间内对植物造成损害的累积效应, 它是一种剂量指标, 与浓度指标相比, 更能反映出臭氧对植物长时间胁迫效应, 其与植物的相对产量之间有较好的相关性[19].计算公式如下:

|
式中, AOT40为大气中臭氧浓度大于40 nL·L-1时的小时累积效应指数.
1.3.2 气孔导度模型气孔导度模型采用Pleijel等2007年修正的Jarvis型气孔导度阶乘式模型[9], 其具体形式如下:

|
式中, GST为气孔导度值[以O3/PLA计, mmol·(m2·s)-1], gmax为冬小麦最大气孔导度, 取值为260.1 mmol·(m2·s)-1. fphen、fO3、fPAR、ftemp和fVPD分别为物候期、O3、光强、温度和水汽压差对气孔导度的限制函数, 该值介于0~1之间.其中, fphen和fO3是最主要的限制因子, 分别用来表示植物正常发育而衰老和由于受到臭氧胁迫效应而导致植物提前衰老的影响.如果fphen < fO3, 则min(fphen, fO3)=fphen, 表示生育期是最主要的限制因子, 反之, fO3成为最主要的限制因子, 通常情况下, 这主要发生在冬小麦生育后期. fmin是在土壤湿度不是限制条件下白天的最小气孔导度, 取值为0.01[20, 21].而fphen函数主要基于热时间的积累而得到, 表示温度对植物发育速率的影响.另外, 有O3胁迫的实测GST从CO2和H2O转化为O3的计算主要根据Graham定律, 分子扩散转化因子为0.61[22].
(1) 光合有效辐射限制函数fPAR

|
式中, L为GST光响应函数的饱和率, 是针对冬小麦的一个常数, PAR为光合有效辐射.
(2) 温度限制函数ftemp

|
式中, t表示小时平均温度(℃), tmax和tmin分别为限制气孔导度使得达到最低值fmin的最高和最低温度. topt为无任何气孔导度限制时的最适温度.

|
(3) 水汽压差限制函数fVPD
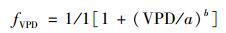
|
式中, VPD为小时水汽压差(kPa), a、b是根据函数拟合后所得到的参数.
(4) 物候限制函数fphen

|
式中, TT表示基于生物学零度的有效积温(℃·d).参数fa和fb分别为开始和结束观测时的气孔导度GST与最大气孔导度gmax的比值.
(5) O3胁迫系数fO3

|
式中, AFsto0根据1.3.3节的相关公式来计算, 当Y值为0时观测期间的叶片累积O3吸收通量.
1.3.3 O3吸收通量模型冬小麦叶片气孔O3吸收通量的计算根据阻力相似原则, 计算公式如下:

|
式中, Fleaf为冬小麦叶片气孔O3通量, rb为叶片边界层阻力, 为叶片边界层导度(gb)的倒数(以O3/PLA计), 取值为1.138 mol·(m2·s)-1[23], rs为气孔阻力, 是气孔导度GST的倒数.
累积气孔O3吸收通量计算公式如下:

|
式中, Fleafi为小时O3吸收通量(nmol·m-2), n为观测期间的小时数(h).
2 结果与分析 2.1 气象因子的变化特征温度(t)、相对湿度(RH)、光合有效辐射(PAR)和水汽压差(VPD)等气象因子是影响冬小麦气孔臭氧吸收通量的主要影响因素, 图 1为2014~2016年冬小麦生长季期间各气象因子的小时变化特征.从中可以看出, 2014~2016年温度的小时变化范围分别为1.4~36.0、0.6~32.2和2.4~31.3℃.相对湿度的变化范围分别为10.9%~90.0%、13.9%~94.9%和17.7%~95.4%.光合有效辐射的变化范围分别为0.0~1 668.3、0.0~1 975.6和0.0~1 753.4 μmol·(m2·s)-1.水汽压差的变化范围分别为0.11~4.65、0.07~3.42和0.07~3.32 kPa.其中2014年的3月1日、3月19日、3月24~25日、3月28日、4月11日、4月18~19日、4月21日、4月25~26日、5月9~10日、5月13日、5月16日、5月24日, 2015年的3月5日、3月14日、3月17~19日、3月30日、4月1~7日、4月12~13日、4月18~19日、4月28日、5月1~2日、5月8日、5月10~11日、5月14日、5月17日、5月26~27日、5月29日, 2016年的3月3~4日、3月7~8日、3月12~13日、3月17日、3月30日、4月2~3日、4月5~6日、4月16日、4月19~20日、4月23~27日、5月1~2日、5月5日、5月9日、5月14~15日、5月20~21日、5月25~28日、5月31日为雨天, 此时温度、光合有效辐射和水汽压差下降, 而相对湿度随着降雨的发生而迅速上升.

|
图 1 冬小麦生长季期间气象因子的小时变化 Fig. 1 Hourly variation of meteorological factorsduring the growing season for winter wheat |
图 2为2014~2016年冬小麦主要生长季期间(即每年的3月1日~5月31日)臭氧浓度的小时变化特征.从中可以看出, 臭氧浓度在整个观测期间从前期到后期总体上呈现逐渐增加的趋势, 5月最大, 3月最小.其中, 2015年平均臭氧浓度最大, 为37.7 nL·L-1. 2014年与2015年相比变化不大, 为36.2 nL·L-1. 2016年最小, 为33.6 nL·L-1.

|
图 2 冬小麦生长季期间臭氧浓度的小时变化 Fig. 2 Hourly variation of ozone concentrationduring the growing season for winter wheat |
为了深入地分析臭氧浓度的时间变化规律, 图 3给出了2014~2016年的每年3~5月臭氧浓度的平均日变化情况, 黑色的实线为平均值, 灰色的阴影部分为标准偏差.从中可知, 一天之中臭氧浓度呈单峰型变化特征, 最低值出现在07:00左右, 之后随着太阳辐射的增强以及温度的上升, NOx和VOCs等臭氧前体物光解速度增加, 导致臭氧浓度开始持续上升, 并在15:00~16:00达到一天中的最大值, 而后随着太阳辐射强度的减弱, 臭氧浓度呈现快速下降的过程.

|
图 3 冬小麦生长期间臭氧浓度的平均日变化特征 Fig. 3 Diurnal variation characteristics of mean ozone concentrationduring the growing season for winter wheat |
图 4是2014~2016年的每年3~5月AOT40的逐日累积值(AOT40day)和连续累积值(AOT40)的变化情况.可以看出, AOT40day在冬小麦整个生长季期间波动比较大, 在冬小麦生长前期, 臭氧浓度较低, AOT40day对AOT40的贡献较少, 而在生长季后期, 臭氧浓度较高, 导致AOT40day对AOT40的贡献较大, 甚至超过500nL·(L·h)-1以上.从2014~2016年, 日小时臭氧浓度大于40 nL·L-1的天数, 即AOT40day>0的天数分别为82、83和79 d, 占总共92 d中的89.13%、90.22%和85.87%.在整个观测期间, 计算得到2014~2016年3~5月AOT40的数值分别为17.08、17.90和11.84 μL·(L·h)-1.

|
图 4 冬小麦生长季期间AOT40day和AOT40的变化 Fig. 4 Changes in AOT40day and AOT40during the growing season for winter wheat |
根据Pleijel等[9]2007年修正的Jarvis型气孔导度阶乘式模型, 在此基础上, 课题组前期对该模型已进行修正和调整, 得到了适用于本地区模型中各参数的数值, 见表 1.并将其代入到1.3.2节的各限制函数中, 计算出了2014~2016年整个观测期间限制函数fPAR、ftemp和fVPD的变化特征, 如图 5所示, 其中黑线表示的是环境因子在冬小麦生长期间日均值的变化, 红线表示的是5日滑动平均值.结合2.1节中气象因子的变化特征可以看出, fPAR的值在雨天条件下较低, 而在晴天条件下, fPAR波动不太明显.而温度限制函数ftemp在整个观测期间总体上呈现逐渐增加的状态, 水汽压差限制函数fVPD则呈现逐渐减小的状态.说明在冬小麦生育前期, 温度成为限制冬小麦气孔导度的重要因素, 而在冬小麦生育中后期, 水汽压差成为限制冬小麦气孔导度的主要因素.
|
|
表 1 气孔导度阶乘式模型中各限制函数的参数数值 Table 1 Limiting functions of the multiplicative stomatal conductance model and values of the function parameters |

|
图 5 环境变量影响因子的变化 Fig. 5 Variations in the impact factors of the environmental variables |
根据1.3.3节中的公式, 分别计算了2014~2016年冬小麦在每年的3~5月每天的小时累积臭氧吸收通量(Fst)和整个观测期间臭氧累积吸收通量(AFsto), 见图 6.从中可以看出, Fst整体上呈现中间高两边低的变化状态, 中间出现的高值区间对应冬小麦扬花期与灌浆期, 此阶段为冬小麦生理活动的最强时期, 气候因子更加有利于植物叶片气孔的张开, 之后随着作物的衰老, 其生理活动减弱, 气孔导度降低, 导致气孔吸收通量变小.因此, 在2014~2016年3~5月的整个观测期间, 冬小麦气孔臭氧累积吸收通量(AFsto)分别为9.36、9.32和8.65 mmol·m-2.

|
图 6 冬小麦生长季期间Fst和AFst的变化 Fig. 6 Changes of Fst and AFst during the growing season for winter wheat |
通过连续观测2014~2016年冬小麦叶片在每年的3~5月气孔导度的小时值, 并根据Pleijel等修订后的气孔导度模型, 结合表 1中模型的参数, 模拟了冬小麦3a的气孔导度值, 并与实测的气孔导度值进行线性回归分析, 以此来对比和验证模型的准确性, 如图 7所示.经过双样本t检验分析, 冬小麦叶片气孔导度的实测值与模拟值之间不存在极显著差异(P<0.01), 决定系数R2为0.81, 表明利用该模型模拟的气孔导度解释了实测值和模拟值中82%的差异性, 线性回归方程为y=1.05x+1.94.

|
图 7 气孔导度模型的验证 Fig. 7 Validation of stomatal conductance model |
利用本课题组前期大田实验中所获取的数据, 建立了冬小麦产量和臭氧剂量(AOT40)之间的函数方程, 发现冬小麦相对产量与AOT40之间有着很好的线性关系[24]:
y=-0.011 1x+0.992 9
式中, y为冬小麦相对产量, x为冬小麦生长季期间AOT40的值.将2014~2016年的每年3~5月的AOT40[17.08、17.90和11.84 μL·(L·h)-1]代入到上述方程, 得到在3 a的臭氧浓度水平下, 冬小麦的相对产量的平均值为81.97%, 即臭氧使冬小麦产量减少了18.03%.
此外, 基于前期大田实验观测数据, Wu等[25]建立了冬小麦干物质累积损失和臭氧累积吸收通量(AFst)的关系:
y=-0.021 4x+1.001 8
式中, y为冬小麦干物质累积相对值, x为冬小麦生长季期间臭氧累积吸收通量的值.将2014~2016年每年的3~5月的AFst(9.36、9.32和8.65 mmol·m-2)代入到上述公式, 得到在3 a的臭氧浓度水平下, 冬小麦的干物质累积相对平均值为80.69%, 即臭氧使冬小麦干物质累积减少了19.31%.
3 讨论近年来, 我国长三角地区臭氧污染日趋严重, 已对植物的生长和产量构成威胁.本文的研究结果表明, 2014~2015年冬小麦生长季期间平均臭氧浓度为35.8nL·L-1, 该值明显高于朱治林等在我国山东省观测的结果[26].研究臭氧对植物影响的最终目的是为了评估臭氧浓度增加对作物产量带来的损失.从20世纪80年代以来, 欧美国家率先在大田实验研究的基础上建立了相关模型来研究臭氧浓度变化对植物生长和产量的影响, 主要经历了浓度响应模型(如Wellbull方程、二次方程和指数方程)[27, 28]、剂量响应模型(如AOT40、SUM06等)和通量响应模型[29, 30].而我国学者在臭氧对植物生物量或产量损失的估算等方面做了一些工作[30~32], 但在相关评估方面尚未找到一个简单的综合指标体系.因此, 本文利用前期课题组通过多年大田实验得到的臭氧剂量-冬小麦产量模型和臭氧通量-冬小麦干物质模型, 估算了近3年臭氧对冬小麦的潜在影响.
目前, 基于AOT40的剂量指标和基于气孔导度模型的通量指标通常被用来评估臭氧对植物的潜在影响.实际上, 基于AOT40的剂量响应指标是一种浓度阈值方法, 它仅仅是利用冠层顶部的臭氧浓度与阈值(40 nL·L-1)之差的累积值而获得, 但是由于该指标未考虑到植物本身的解毒能力、防御能力和夜间的修复能力, 这在一定程度上会高估臭氧的胁迫效应, 使得该阈值并不具备明确的生物学意义[18, 33],
气孔是植物叶片与外界环境进行气体交换最主要的途径, 气孔导度表示的是气孔张开的程度, 影响着植物的蒸腾、光合和呼吸作用等.当臭氧通过叶片气孔后, 首先影响的是靠近气孔的细胞, 使得气孔减小或闭合, 而气孔的关闭或减小将会减少叶片对二氧化碳的吸收和水汽通量, 从而影响其光合速率.此外, 臭氧进入植物体后将会导致叶绿体结构发生改变, 从而使得植物的光合能力下降[34].气孔通量指的是叶片气孔吸收的臭氧量, 它由气孔导度和臭氧浓度共同决定.有研究表明, 基于气孔导度模型的通量响应指标, 考虑了温度、光合有效辐射、生育期、水汽压差和臭氧浓度等气候因子对植物叶片气孔通量的影响, 通过研究生物量和产量的减产率与气孔通量的关系比AOT40更加紧密, 具有很好的适用性, 能够更准确评估臭氧的环境影响[9, 15, 21].例如, Pleijel等[35]通过比较小麦和土豆产量与AOT40指标和通量指标之间的关系, 发现两种植物的相对产量与通量指标的相关性比AOT40指标更加显著. Karlsson等[36]认为臭氧吸收通量比AOT40能更好地反映森林树木叶片伤害和生物量减少的程度. Mills等[37]整合分析了欧洲国家近20年有关臭氧对植物影响的成果, 通过研究发现利用基于臭氧通量的评估指标比基于臭氧浓度的评估指标要好.在我国, Feng等[32]的研究表明, 气孔累积吸收通量与水稻产量的相关性高于AOT40等剂量指标.佟磊等[38]的研究发现, 水稻产量与累积气孔臭氧通量具有显著的线性递减关系, 其相关系数高于臭氧浓度指标.而他在冬小麦实验研究中发现[39], 基于AOT40的剂量指标比基于臭氧通量的指标在冬小麦产量损失评估中更占优势, 这可能是由于数据样本不足降低了模型参数的拟合精度, 与此对应的是Gonzalez-Fernandez等[40]的研究与之类似.
本研究得到的冬小麦产量和干物质减少的结果在一定程度上可能会高估植物产量和干物质累积的损失.研究表明, 植物对臭氧的胁迫具有一定的适应和解毒能力, 细胞壁上的抗坏血酸盐等抗氧化剂将分解小部分臭氧[41], 现有的研究通常设定一个阈值为y [nmol·(m2·s)-1], 即超过该阈值的臭氧吸收通量将会对植物产量等造成损失, 也即没有臭氧吸收或其值较低时将不抑制光合作用. Pleijel等[9]的研究表明, 该阈值y取值为6 nmol·(m2·s)-1时计算的累积臭氧吸收通量(即称为臭氧吸收有效剂量)的变化与植物产量损失相对值的相关性最高. Zhang等认为[10], 当y取值为9.6 nmol·(m2·s)-1时, 其计算的POD9.6的变化与大豆产量损失相对值的相关性最为显著.佟磊等[38]通过研究水稻臭氧通量-产量关系时发现, 当y为2 nmol·(m2·s)-1时, 气孔臭氧累积吸收通量与水稻产量的相关性最高. Wu等指出[25], 当阈值为4 nmol·(m2·s)-1时, POD04与冬小麦产量损失相对值的关系最为紧密, 这与佟磊等[39]的研究结果一致, 但却低于国外学者对欧洲小麦的研究结果[y为5~14 nmol·(m2·s)-1][9, 21].事实上, 由于不同作物、不同生育期以及不同气候因子的差异, 使得植物对臭氧的清除能力存在较大的波动, 张巍巍等[42]的研究则表明, 既使是同一作物, 由于品种间的不同, 对臭氧胁迫响应也存在差异[43]. Massman[44]认为臭氧伤害阈值是光合速率的函数, 随生育进程的光合能力变化而变化.目前, 由于研究的局限性, 还无法确定y的动态变化过程, 只能通过建立PODy与产量损失的关系, 从而找到其合适的阈值常数.本文在利用气孔导度模型结合通量模型模拟冬小麦臭氧通量时并未将y值考虑在内, 这在一定程度上高估了冬小麦产量的损失.因此, 结合臭氧吸收通量模型, 开展植物解毒效应的观测与模拟是当前和未来研究的热点问题.
目前, 臭氧通量的观测方法主要有箱式法、模型拟合法和微气象学法.其中基于涡度相关法的微气象学法是近几年发展起来的一项新的先进技术, 并在国外得到一定的运用[45, 46], 但是在中国该项技术还处于刚起步阶段, 我国科学家朱治林研究团队运用涡度相关通量法率先在鲁西北地区冬小麦麦田开展臭氧通量的观测, 弥补了我国在这一领域内的空缺, 同时也是当前我国研究臭氧对植物影响评估的一个升华[47].本研究的下一步工作将引进国外先进的涡度相关技术, 在我国长三角地区冬小麦下垫面开展臭氧通量的长期观测, 为更加科学合理地评估臭氧对植物的影响提供强有力的方法和数据支撑.
4 结论(1) 2014~2016年冬小麦主要生长季期间(每年的3月1日~5月31日), 平均臭氧浓度分别为36.2、37.7和33.6 nL·L-1.臭氧浓度从前期到后期呈现逐渐增加的状态, 并呈现明显的单峰型日变化特征.整个观测期间, 2014~2016年的每年3~5月麦田AOT40值分别为17.08、17.90和11.84 μL·(L·h)-1.
(2) 利用Javis气孔导度模型, 计算了冬小麦叶片气孔导度的变化, 经过与实测值的对比验证, 表明该模型可以用来模拟本地区冬小麦的气孔导度.同时结合臭氧通量模型, 模拟了2014~2016年冬小麦气孔臭氧吸收通量分别为9.36、9.32和8.65 mmol·m-2.
(3) 利用前期课题组建立的模型, 估算了在近3年臭氧浓度平均水平下, 近地层臭氧会使冬小麦产量减少18.03%, 干物质累积减少19.31%.
| [1] | Vingarzan R. A review of surface ozone background levels and trends[J]. Atmospheric Environment, 2004, 38(21): 3431-3442. DOI:10.1016/j.atmosenv.2004.03.030 |
| [2] | Meehl G A, Stocker T F, Collins W D, et al. Contribution of Working Group I to the Fourth Assessment Report of theintergovernmental panel on climate change[R].Cambridge:Cambridge University Press, 2007. http://www.mendeley.com/catalog/contribution-working-group-i-fourth-assessment-report-intergovernmental-panel-climate-change/ |
| [3] | Ashmore M, Toet S, Emberson L. Ozone-a significant threat to future world food production?[J]. New Phytologist, 2006, 170(2): 201-204. DOI:10.1111/nph.2006.170.issue-2 |
| [4] | Avnery S, Mauzerall D L, Liu J F, et al. Global crop yield reductions due to surface ozone exposure:1. Year 2000 crop production losses and economic damage[J]. Atmospheric Environment, 2011, 45(13): 2284-2296. DOI:10.1016/j.atmosenv.2010.11.045 |
| [5] | Avnery S, Mauzerall D L, Liu J F, et al. Global crop yield reductions due to surface ozone exposure:2. Year 2030 potential crop production losses and economic damage under two scenarios of O3 pollution[J]. Atmospheric Environment, 2011, 45(13): 2297-2309. DOI:10.1016/j.atmosenv.2011.01.002 |
| [6] | Feng Z W, Jin M H, Zhang F Z, et al. Effects ofground-level ozone (O3) pollution on the yields of rice andwinter wheat in the Yangtze River Delta[J]. JournalofEnvironmental Sciences, 2003, 15(3): 360-362. |
| [7] |
赵辉, 郑有飞, 曹嘉晨, 等. 大气臭氧污染对冬小麦气孔吸收通量的影响机制及其时空格局[J]. 环境科学, 2017, 38(1): 412-422. Zhao H, Zheng Y F, Cao J C, et al. Influencing mechanism and spatio-temporal pattern of stomatal ozone flux of winter wheat under ozone Pollution[J]. Environmental Science, 2017, 38(1): 412-422. |
| [8] |
郑有飞, 胡程达, 吴荣军, 等. 地表臭氧浓度增加对冬小麦光合作用的影响[J]. 生态学报, 2010, 30(4): 847-856. Zheng Y F, Hu C D, Wu R J, et al. Experiment with effects of increased surface ozone concentration upon winter wheat photosynthesis[J]. Acta Ecologica Sinica, 2010, 30(4): 847-856. |
| [9] | Pleijel H, Danielsson H, Emberson L, et al. Ozone risk assessment for agricultural crops in Europe:further development of stomatal flux and flux-response relationships for European wheat and potato[J]. Atmospheric Environment, 2007, 41(14): 3022-3040. DOI:10.1016/j.atmosenv.2006.12.002 |
| [10] | Zhang W W, Feng Z Z, Wang X K, et al. Quantification of ozone exposure-and stomatal uptake-yield responserelationships for soybean in Northeast China[J]. Science of the Total Environment, 2017, 599-600: 710-720. DOI:10.1016/j.scitotenv.2017.04.231 |
| [11] | Zhu Z L, Sun X M, Zhao F H, et al. Ozone concentrations, flux and potential effect on yield during wheat growth in the Northwest-Shandong Plain of China[J]. JournalofEnvironmental Sciences, 2015, 34: 1-9. |
| [12] | Jin M H, Feng Z W, Zhang F Z. Impacts of ozone on the biomass and yield of rice in open-top chambers[J]. JournalofEnvironmental Sciences, 2001, 13(2): 233-236. |
| [13] |
郑启伟, 王效科, 谢居清, 等. 外源抗坏血酸对臭氧胁迫下水稻叶片膜保护系统的影响[J]. 生态学报, 2006, 26(4): 1131-1137. Zheng Q W, Wang X K, Xie J Q, et al. Effects of exogenous ascorbate acid on membrane protective system of in situ rice leaves under O3 stress[J]. Acta Ecologica Sinica, 2006, 26(4): 1131-1137. |
| [14] | Emberson L D, Büker P, Ashmore M R, et al. A comparison of North American and Asian exposure-response data for ozoneeffects on crop yields[J]. Atmospheric Environment, 2009, 43(12): 1945-1953. DOI:10.1016/j.atmosenv.2009.01.005 |
| [15] | Keller F, Bassin S, Ammann C, et al. High-resolution modelling of AOT40 and stomatal ozone uptake in wheat and grassland:a comparison between 2000 and the hot summer of 2003 in Switzerland[J]. Environmental Pollution, 2007, 146(3): 671-677. DOI:10.1016/j.envpol.2006.06.025 |
| [16] | Mills G, Buse A, Gimeno B, et al. A synthesis of AOT40-based response functions and critical levels of ozone for agricultural and horticultural crops[J]. Atmospheric Environment, 2007, 41(12): 2630-2643. DOI:10.1016/j.atmosenv.2006.11.016 |
| [17] | Ferretti M, Cristofolini F, Cristofori A, et al. A simple linear model for estimating ozone AOT40 at forest sites from raw passive sampling data[J]. Journal of Environmental Monitoring, 2012, 14(8): 2238-2244. DOI:10.1039/c2em30137g |
| [18] | Emberson L D, Ashmore M R, Cambridge H M, et al. Modelling stomatal ozone flux across Europe[J]. Environmental Pollution, 2000, 109(3): 403-413. DOI:10.1016/S0269-7491(00)00043-9 |
| [19] | Pleijel H, Ojanpera K, Danielsson H, et al. Effects of ozone on leaf senescence in spring wheat-possible consequences for grain yield[J]. PhytonAnnales Rei Botanicae, 1997, 37(3): 227-232. |
| [20] | Pleijel H, Danielsson H, Vandermeiren K, et al. Stomatal conductance and ozone exposure in relation to potato tuber yield-results from the European CHIP programme[J]. European Journal of Agronomy, 2002, 17(4): 303-317. DOI:10.1016/S1161-0301(02)00068-0 |
| [21] | Danielsson H, Karlsson G P, Karlsson P E, et al. Ozone uptake modelling and flux-response relationships-an assessment of ozone-induced yield loss in spring wheat[J]. Atmospheric Environment, 2003, 37(4): 475-485. DOI:10.1016/S1352-2310(02)00924-X |
| [22] | Campbell G S, Norman J M. An introduction to environmental biophysics (2nd ed.)[M]. New York, Berlin, Heidelberg: Springer, 1998: 286. |
| [23] | Makowski D, Doré T, Monod H. A new method to analyse relationships between yield components with boundary lines[J]. Agronomy for Sustainable Development, 2007, 27(2): 119-128. DOI:10.1051/agro:2006029 |
| [24] | 郑有飞, 吴荣军. 地表臭氧变化特征及其作物响应[M]. 北京: 气象出版社, 2012. |
| [25] | Wu R J, Zheng Y F, Hu C D. Evaluation of the chronic effects of ozone on biomass loss of winter wheat based on ozone flux-response relationship with dynamical flux thresholds[J]. Atmospheric Environment, 2016, 142: 93-103. DOI:10.1016/j.atmosenv.2016.07.025 |
| [26] |
朱治林, 孙晓敏, 赵风华, 等. 鲁西北平原冬小麦田臭氧浓度变化特征及对产量的潜在影响和机理分析[J]. 植物生态学报, 2012, 36(4): 313-323. Zhu Z L, Sun X M, Zhao F H, et al. Variation of ozone concentration of winter wheat field and mechanistic analysis of its possibleeffect on wheat yield in Northwest-Shandong Plain of China[J]. Chinese Journal of Plant Ecology, 2012, 36(4): 313-323. |
| [27] | Heck W W, Adams R M, Cure W W, et al. A reassessment of crop loss from ozone[J]. Environmental Science and Technology, 1983, 17(12): 572A-581A. DOI:10.1021/es00118a716 |
| [28] | Larsen R M, Heck W W. An air quality data analysis system for interrelating effects, standards, and needed source reductions:part8. An effective mean O3 crop reduction mathematical model[J]. Journal of the Air Pollution Control Association, 1984, 34(10): 1023-1034. DOI:10.1080/00022470.1984.10465850 |
| [29] | Pleijel H, Ojanperä K, Mortensen L. Effects of tropospheric ozone on the yield and grain protein content of spring wheat (Triticum aestivum L.) in the Nordic countries[J]. Acta Agriculturae Scandinavica, Section B-Soil & Plant Science, 1997, 47(1): 20-25. |
| [30] | UNECE. Revised manual on methodologies and criteria for mapping critical levels/loads and geographical areas where they are exceeded[R]. Berlin:Federal Environment Agency, 1996. |
| [31] |
吴荣军, 郑有飞, 赵泽, 等. 基于气孔导度和臭氧吸收模型的冬小麦干物质累积损失评估[J]. 生态学报, 2010, 30(11): 2799-2808. Wu R J, Zheng Y F, Zhao Z, et al. Assessment of loss of accumulated dry matter in winter wheat based on stomatalconductance and ozone uptake model[J]. Acta Ecologica Sinica, 2010, 30(11): 2799-2808. |
| [32] | Feng Z Z, Tang H Y, Uddling J, et al. A stomatal ozone flux-response relationship to assess ozone-induced yield loss of winter wheat in subtropical China[J]. Environmental Pollution, 2012, 164: 16-23. DOI:10.1016/j.envpol.2012.01.014 |
| [33] | Cieslik S. Ozone fluxes over various plant ecosystems in Italy:a review[J]. Environmental Pollution, 2009, 157(5): 1487-1496. DOI:10.1016/j.envpol.2008.09.050 |
| [34] |
张巍巍, 赵天宏, 王美玉, 等. 臭氧浓度升高对银杏光合作用的影响[J]. 生态学杂志, 2007, 26(5): 645-649. Zhang W W, Zhao T H, Wang M Y, et al. Effects of elevated ozone concentration on Ginkgo biloba photosynthesis[J]. Chinese Journal of Ecology, 2007, 26(5): 645-649. |
| [35] | Pleijel H, Danielsson H, Ojanperä K, et al. Relationships between ozone exposure and yield loss in European wheat and potato-a comparison of concentration-and flux-based exposure indices[J]. Atmospheric Environment, 2004, 38(15): 2259-2269. DOI:10.1016/j.atmosenv.2003.09.076 |
| [36] | Karlsson P E, Uddling J, Skärby L, et al. Impact of ozone on the growth of birch (Betula pendula) saplings[J]. Environmental Pollution, 2003, 124(3): 485-495. DOI:10.1016/S0269-7491(03)00010-1 |
| [37] | Mills G, Hayes F, Simpson D, et al. Evidence of widespread effects of ozone on crops and (semi-)natural vegetation in Europe (1990-2006) in relation to AOT40-and flux-based risk maps[J]. Global Change Biology, 2011, 17(1): 592-613. DOI:10.1111/gcb.2010.17.issue-1 |
| [38] |
佟磊, 王效科, 苏德·毕力格, 等. 水稻气孔臭氧通量拟合及通量与产量关系的比较分析[J]. 农业环境科学学报, 2011, 30(10): 1930-1938. Tong L, Wang X K, Sudebilige, et al. Stomatal ozone uptake modeling and comparative analysis of flux-response relationships of rice[J]. Journal of Agro-Environment Science, 2011, 30(10): 1930-1938. |
| [39] |
佟磊, 冯宗炜, 苏德·毕力格, 等. 冬小麦气孔臭氧通量拟合及通量产量关系的比较分析[J]. 生态学报, 2012, 32(9): 2890-2899. Tong L, Feng Z W, Sudebilige, et al. Stomatal ozone uptake modeling and comparative analysis of flux-response relationships of winter wheat[J]. Acta Ecologica Sinica, 2012, 32(9): 2890-2899. |
| [40] | Gonzalez-Fernandez I, Kaminska A, Dodmani M, et al. Establishing ozone flux-response relationships for winter wheat:analysis of uncertainties based on data for UK and Polish genotypes[J]. Atmospheric Environment, 2010, 44(5): 621-630. DOI:10.1016/j.atmosenv.2009.11.021 |
| [41] | Barnes J, Zheng Y B, Lyons T. Plant resistance to ozone:the role of ascorbate[A]. In:Osama K, Saji H, Youssefian S, et al (Eds.).Air Pollution and Plant Biotechnology[M]. Tokyo:Springer, 2002.235-252. |
| [42] |
张巍巍, 王光华, 王美玉, 等. 东北春大豆品种东生1号对臭氧胁迫的响应[J]. 环境科学, 2014, 35(4): 1473-1478. Zhang W W, Wang G H, Wang M Y, et al. Responses of soybean cultivar Dongsheng-1 to different O3 concentrations in Northeast China[J]. Environmental Science, 2014, 35(4): 1473-1478. |
| [43] | Zhang W W, Wang G H, Liu X B, et al. Effects of elevated O3 exposure on seed yield, N concentration andphotosynthesis of nine soybean cultivars(Glycine max (L.) Merr.) inNortheast China[J]. Plant Science, 2014, 226: 172-181. DOI:10.1016/j.plantsci.2014.04.020 |
| [44] | Massman W J. Toward an ozone standard to protect vegetation based on effective dose:a review of deposition resistances and a possible metric[J]. Atmospheric Environment, 2004, 38(15): 2323-2337. DOI:10.1016/j.atmosenv.2003.09.079 |
| [45] | Fares S, Matteucci G, Mugnozza G S, et al. Testing of models of stomatal ozone fluxes with field measurements in a mixed Mediterranean forest[J]. Atmospheric Environment, 2013, 67: 242-251. DOI:10.1016/j.atmosenv.2012.11.007 |
| [46] | Launiainen S, Katul G G, Grönholm T, et al. Partitioning ozone fluxes between canopy and forestfloor by measurements and a multi-layer model[J]. AgriculturalandForestMeteorology, 2013, 173: 85-99. |
| [47] |
朱治林, 孙晓敏, 董云社, 等. 鲁西北平原玉米地涡度相关臭氧通量日变化特征[J]. 中国科学:地球科学, 2014, 57(2): 503-511. Zhu Z L, Sun X M, Dong Y S, et al. Diurnal variation of ozone flux over corn field in Northwestern Shandong Plain of China[J]. Science China Earth Sciences, 2014, 57(3): 503-511. |