 2017, Vol. 38
2017, Vol. 38



2. 华东师范大学上海市城市化生态过程与生态恢复重点实验室, 上海 200241
2. Shanghai Key Laboratory for Urban Ecological Processes and Eco-Restoration, East China Normal University, Shanghai 200241, China
汞是环境中重要的金属污染物, 可经食物链对人体产生严重的毒害作用.因在常温下具有较高的蒸气压, 汞主要以气态存在于大气中.有研究表明大气中的气态单质汞可以随着大气循环进行长距离传输, 再经过干湿沉降至地表, 污染生态环境[1, 2].湿地是一种独特的生态系统, 其水体含有较为丰富的可溶性碳及腐殖酸, 它们可以和汞形成稳定的络合物.所以湿地能够将大气沉降和地表径流等输入的汞固定下来, 成为了汞的活性库[3, 4].湿地水体中Hg2+经光照、化学、微生物还原等形成溶解性气态元素汞(Hg0)[5, 6]及沉积物在厌氧条件下经微生物和其他非生物因素作用还原产生的Hg0, 这些可通过扩散进入水层并向大气释放[7, 8].
湿地植物根、茎中通常具有发达的通气组织, 以利于缺氧条件下由地上向根部运输氧气满足根细胞的呼吸.这些通气组织是否也可成为还原性沉积物中产生的气态汞由地下向地上运输的通道?有研究发现, 湿地植物(香蒲、克拉莎草、芦苇)存在的条件下, 大大提高了湿地生态系统的汞释放通量[9].在佛罗里达的野外实验也表明生长香蒲的湿地, 其汞的释放通量远高于无植物的水面, 且释放量与蒸腾作用相关[10].通过这些为数不多的野外研究, 可以推测根系可能在根际气体的运输中扮演着重要的角色, 但尚未有直接的证据, 其中的过程和机制有待进一步研究.
因此, 本研究以水稻这一典型的湿地植物为例, 通过室内模拟实验, 设计分室密闭装置将水稻根系与水稻地上部分隔开, 对水稻根系进行气态单质汞暴露后, 收集由水稻叶片的汞释放量, 以探究水稻根系是否能吸收气态单质汞, 并将其转移到地上部释放到空气中, 以及其中的影响因素.本研究结果将进一步丰富植被在汞的生物地球化学循环中的作用.
1 材料与方法 1.1 实验材料通过预实验, 选定以下3种具有良好重金属耐性的水稻品种(表 1):其中X1为籼稻, 长势快, 茎秆粗壮, 抗倒伏. J1为粳稻, 长势较快, 茎秆较为粗壮.而J2也为粳稻, 但长势一般, 茎秆较细.预实验表明这3种水稻根、叶的生理生态指标具有明显的差异.
|
|
表 1 水稻品种信息 Table 1 Information on rice species |
1.2 研究方法 1.2.1 水稻幼苗培养方法
将挑选好的水稻种子放在30%的H2O2溶液中浸泡40 min, 用蒸馏水冲洗5遍, 洗净后置于垫有滤纸的培养皿中, 将培养皿放入28℃恒温培养箱中培养3 d, 再将幼苗转移到沙土培养基中培养5 d.水培:挑选生长较好、长势一致的幼苗转移到添加Kimura B营养液(pH=5.5)[11]的5LPVC盆中培养20d.所有水稻在人工气候箱中进行培养, 培养条件:湿度60%, 28℃/14 h光照和22℃/10 h黑暗, 光照强度为260 μmol·(m2·s)-1.
1.2.2 分室密闭实验装置为了避免气态汞的交叉污染, 本研究需要设计能将地上和地下组织严格区分的密闭实验装置, 各分室严格的气密性是实验成功的关键.为此, 利用耐高温的硼化硅材料定做了分室密闭反应室(图 1).该装置分为两个部分, 上部气室(即叶室)高30 cm, 直径12.5 cm, 下部气室(即根室)高10 cm, 直径12.5 cm.上下部气室中间开直径4 cm的圆孔, 圆孔中放置一个开有直径为5 mm孔洞的硅胶塞, 植物根、茎结合处放置于硅胶塞上的孔洞中.为保证上下两个气室严格密封, 植物与硅胶塞的之间封以黏土和凡士林, 硅胶塞与气室封以凡士林.叶室上方通过两个串联的气体吸附剂1和2(Milestone Sorbent Traps), 以吸收由叶片挥发的Hg0; 而为保证叶室的气压稳定, 在叶室下方通过充气泵充入空气(连接50 mL 200 mmol·L-1 KMnO4溶液除去空气中的Hg0).

|
图 1 分室密闭反应室系统 Fig. 1 Chamber closed reaction chamber system |
将培养好的水稻用蒸馏水洗净根部, 放入分室密闭反应室中, 将水稻地上部和根分别暴露在上下气室中, 并在下部气室放入3 g单质汞(根据27℃下汞的饱和蒸气压计算得出下部气室汞蒸气浓度约为0.021 5 g·m-3), 充气泵通过浮子流量计提供流速为200 mL·min-1的恒定空气气流, 气流通过洗气瓶去除空气中含有的汞, 随后气流进入上部气室将水稻挥发出的气体通过出气口载流到气体吸附剂1和2中进行吸收, 供气反应时间为150 min.进行3组重复.
1.3.2 去叶水稻汞挥发实验为了探究水稻叶片在汞挥发过程中的作用及其所挥发出的汞的来源, 实验还收集了叶片去除后的水稻挥发出的汞.选取长势一致的水稻X1作为样本, 将其叶片沿茎切除, 地上部只保留5 cm的茎, 按1.3.1节的方法收集地上部分挥发的汞.
1.3.2 水稻生理指标测定① 用甲基蓝染液将水稻根系染色, 用扫描仪(EPSON EU-22)扫描完整根系图片, 并用根系分析软件winrhizo分析根长、根表面积及根体积. ②水稻的根孔隙度利用改进的比重瓶法进行测量[12]. ③水稻叶片用扫描仪(EPSON EU-22)扫描获得完整叶片图片, 用软件ImageJ处理获得水稻叶面积[13]. ④利用改进的快速称重法测定水稻蒸腾作用强度[14, 15].
1.3.3 汞含量测定植物组织(冻干)、吸附剂中的汞均由直接测汞仪(Milestone DMA-80)测定, 即样品经过高温分解后被持续的高纯氧气流载入到催化剂管, 后所有的汞被带到金汞齐管, 通过瞬间加热, 汞蒸气被释放到单波长253.7 nm原子吸收分光光度计中进行测量.
1.4 计算方法及统计分析通过测定, 对数据进行以下处理.
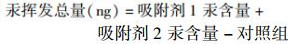
|
(1) |
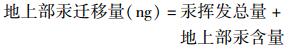
|
(2) |
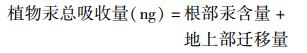
|
(3) |
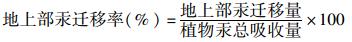
|
(4) |
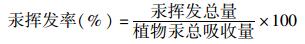
|
(5) |
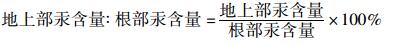
|
(6) |
利用SPSS软件对上述获得数据进行方差分析、显著性检验及相关性分析并作图.
2 结果与分析 2.1 3种基因型水稻的生理指标水稻生理相关指标如表 2, 水稻X1与J2和J1在叶面积、根表面积、根干重和蒸腾作用强度有显著性差异(P < 0.05), 其中X1叶面积和根表面积最高, 蒸腾作用强度最低.根孔隙度、根体积这3种水稻差异性显著(P < 0.05), 其中X1根体积最高, J1根孔隙度最高.比根面积、根冠比、根长这3种水稻无显著性差异(P>0.05).
|
|
表 2 水稻生理生态指标 Table 2 Physio-ecological indexes for rice |
2.2 3种基因型水稻测汞数据
通过收集测定, 水稻叶片汞挥发量及水稻样品中汞含量结果见表 3.研究结果表明, 水稻根部暴露在汞蒸气时, 根系可以吸收气态汞单质并向地上部少量转运, 再通过叶片向空气中释放.其中叶片汞挥发总量水稻X1最高, 并与J2和J1有显著性差异(P < 0.05).地上部汞含量与地上部汞迁移量三者都具显著性差异(P < 0.05), 其中水稻X1最高、J1最低.而单位叶面积汞挥发量三者无显著性差异(P < 0.05).水稻J1根部汞含量及植物汞总吸收量较低且与J1和X1相比有显著性差异(P < 0.05).这些差异产生的原因有可能是和根的生物量有关.
|
|
表 3 水稻汞相关数据 Table 3 Data for mercury in rice |
2.3 去除叶片水稻与有叶水稻的比较
将同生长期的水稻X1去除叶片进行汞挥发实验, 与正常水稻(未去除叶片)进行对比, 结果如表 4.去叶水稻的汞挥发总量是正常有叶水稻汞挥发总量的3倍, 差异显著(P < 0.05).而正常水稻地上部汞总量是去叶水稻的3倍, 并呈显著性差异(P < 0.05).地上部汞迁移总量、根部汞总量、植物汞总吸收量两者相似, 无显著性差异(P>0.05).
|
|
表 4 去除叶片水稻的汞含量数据 Table 4 Data for mercury in leafless rice |
2.4 水稻汞含量相关性分析
水稻根孔隙度与水稻根部汞含量呈显著负相关, 并呈二次拟合(R=0.830 9, P < 0.01, 如图 2), 说明随着水稻的孔隙度的增加, 根部的汞含量是先增加后减少的.

|
图 2 水稻根汞含量与根孔隙度相关性 Fig. 2 Relationship between root mercury content and root porosity in rice |
水稻的地上部汞迁移总量占植物汞总吸收总量比例范围在1.3%~3%之间(如表 5), 其中J1地上部迁移率最高, J2水稻地上部迁移率最低, 且J1和J2具显著性差异(P < 0.05).水稻的汞挥发量占植物吸收汞总量比例较低, 小于1.3%, 且挥发率X1和J2与J1呈显著性差异(P < 0.01).说明水稻根吸收的气态单质汞只有很少的一部分从根部转移到地上部, 并挥发出去.
|
|
表 5 水稻汞挥发迁移比例/% Table 5 Percentage of mercury volatilization in rice/% |
水稻的地上部汞迁移总量与根表面积呈线性显著正相关(R=0.896, P < 0.01), 且与根体积呈线性显著正相关(R=0.871, P < 0.01, 如图 3、4).表明水稻根系吸收的汞转移到地上部的量与根系结构具有密切相关性, 根系越发达转移到地上部的汞量越多.

|
图 3 水稻根体积和地上部汞迁移总量相关性 Fig. 3 Relationship between the root volume of rice and total amount of mercury transported above ground |

|
图 4 水稻根表面积与地上部汞迁移总量相关性 Fig. 4 Relationship between total root surface area and above ground mercury transport of rice |
通过实验表明, 水稻叶片可以挥发气态单质汞, 并且汞挥发总量与水稻叶面积呈线性显著正相关(R=0.897 07, P < 0.01, 如图 5), 说明水稻汞的释放可能是通过水稻叶片进行的, 且释放量随着水稻叶面积的增加而增大.而水稻单位叶面积汞挥发量与水稻蒸腾作用强度呈线性显著正相关(R=0.73, P < 0.01, 如图 6).

|
图 5 水稻叶面积与汞挥发总量相关性 Fig. 5 Relationship between rice leaf area and total amount of mercury volatilization |

|
图 6 水稻蒸腾作用强度与单位叶面积汞挥发量相关性 Fig. 6 Relationship between rice transpiration intensity and unit leaf area of mercury volatilization |
通过设计分室密闭装置, 对水稻根系进行气态单质汞暴露, 并收集叶片挥发的气体.经过测定, 发现水稻根系能吸收气态单质汞, 并将其转运到地上部, 再挥发到空气中.水稻的汞挥发总量籼稻和粳稻具显著性差异(P < 0.05), 且汞挥发总量与水稻的叶面积呈显著正相关(R=0.897, P < 0.01). 3种基因型水稻的单位叶面积汞挥发量虽略有差异, 但并不显著(P>0.05).
为了探究水稻叶片在汞挥发过程中的作用及其所挥发出的汞的来源, 将水稻叶片沿茎切除, 地上部只保留5 cm的茎秆, 并对其进行汞挥发实验.通过与有叶的水稻对比, 发现水稻去除叶片并不影响根系吸收气态单质汞及根系转移到地上部汞的量, 但去除叶片导致水稻汞挥发总量显著增高.水稻根、茎叶中具有丰富的通气组织, 它对水稻与大气中气体交换和传输起着非常关键的作用[16~18]. Butterbach-Bahl等[19]在之前对水稻吸收排放CH4的研究中发现, 水稻向大气中排放CH4主要是通过通气组织进行的, 并且从根系转移到地上部分的主要运输阻力是根-茎结合部, 不同基因型水稻气体运输的差异是通气组织形态差异的结果, 特别是根-茎结合部结构差异.在本实验中, 并未破坏水稻的根-茎结合部, 所以地上部汞迁移量并未发生显著变化.综上推测水稻根系吸收气态单质汞并通过根-茎结合部转运到茎, 当地上部只保留5 cm茎的时候, 单质汞就通过茎的空腔直接释放到空气中.而地上部完整的时候, 转运到地上部的汞就只能通过水稻叶片的气孔释放到空气中.
水稻孔隙度的大小是表征水稻根系通气组织的重要指标, 水稻根系通气组织是植物薄壁组织内的细胞衰老坏死形成的空腔[20, 21], 水稻孔隙度越大, 其根部活细胞越少.在研究中发现水稻根孔隙度与水稻根部汞含量呈显著负相关, 并呈二次拟合(R=0.830 9, P < 0.01), 说明水稻根部吸收气态单质汞可能与水稻根部的活细胞的新陈代谢有关.
在实验中通过计算水稻汞的迁移率, 发现水稻根吸收的汞只有很少的一部分被转运到了地上部, 迁移率小于3%, 挥发的比例更低, 小于1.3%. Greger等的研究发现[22], 当对6种陆生植物(甜菜、小麦等)根部暴露200 μg·L-1的Hg2+时, 汞从根部转移到地上部的量也较少(0.17%~2.5%).而Du等的研究表明[23], 水稻汞累积主要保留在水稻根部, 数据表明水稻根部积累的汞浓度大约是水稻地上部汞浓度的20倍.地上部汞迁移率不同水稻略有差异, 其中J2迁移率最低、J1最高.而水稻的地上部汞迁移总量与根表面积呈线性显著正相关(R=0.896, P < 0.01), 且与根体积呈线性显著正相关(R=0.871, P < 0.01).所以水稻根系对气态单质汞向地上部的转运量与水稻根系密切相关.
本实验还发现水稻单位叶面积汞挥发量与水稻蒸腾作用强度呈线性显著正相关(R=0.73, P < 0.01).而蒸腾作用主要是以气孔蒸腾进行的, 气孔是气体交换和水分散失的通道, 植物的蒸腾作用一般可表征气孔活动情况.当进行蒸腾作用的时候, 植物叶片气孔处于开放的状态[24]. Borah等[25]发现水稻的CH4排放量与水稻的叶片蒸腾作用强度和气孔导度呈显著正相关. Philippot等[26]的研究发现在水稻淹水培养下, 产生的CH4中90%通过通气组织释放到大气中. Battke等[27]通过室内培养实验发现, 植物可吸收Hg2+, 在体内转化为汞单质并通过植物叶片释放到空气中. Leonard等[28]通过气体交换系统发现植物能作为汞从土壤到大气中的运输管道.所以水稻根部吸收气态单质汞后, 转运到地上部, 再通过叶片的气孔排放到大气中去.
4 结论水稻根系能吸收气态单质汞, 并通过根系的通气组织向地上部少量转运, 转运量与水稻根系结构相关.转运到地上部的气态单质汞通过通气组织转移到叶片, 并通过叶片上的气孔释放到空气中.
| [1] | Lindqvist O, Johansson K, Bringmark L, et al. Mercury in the Swedish environment-recent research on causes, consequences and corrective methods[J]. Water, Air, and Soil Pollut, 1991, 55(1-2): xi-261. DOI:10.1007/BF00542429 |
| [2] | Lindberg S, Bullock R, Ebinghaus R, et al. A synthesis of progress and uncertainties in attributing the sources of mercury in deposition[J]. AMBIO:A Journal of the Human Environment, 2007, 36(1): 19-33. DOI:10.1579/0044-7447(2007)36[19:ASOPAU]2.0.CO;2 |
| [3] |
丁振华, 刘金铃, 李柳强, 等. 中国主要红树林湿地沉积物中汞的分布特征[J]. 环境科学, 2009, 30(8): 2210-2215. Ding Z H, Liu J L, Li L Q, et al. Distribution of mercury in surficial sediments from main mangrove wetlansofChina[J]. Environmental Science, 2009, 30(8): 2210-2215. |
| [4] |
何天容, 冯新斌, 郭艳娜, 等. 红枫湖沉积物中汞的环境地球化学循环[J]. 环境科学, 2008, 29(7): 1768-1774. He T R, Feng X B, Guo Y N, et al. Geochemical cycling of mercury in the sediment of Hongfengreservior[J]. Environmental Science, 2008, 29(7): 1768-1774. |
| [5] | Zhang H, Lindberg S E. Air/water exchange of mercury in the Everglades Ⅰ:the behavior of dissolved gaseous mercury in the Everglades nutrient removal project[J]. Science of the Total Environment, 2000, 259(1-3): 123-133. DOI:10.1016/S0048-9697(00)00577-5 |
| [6] | Fantozzi L, Ferrara R, Frontini F P, et al. Dissolved gaseous mercury production in the dark:evidence for the fundamental role of bacteria in different types of Mediterranean water bodies[J]. Science of the Total Environment, 2009, 407(2): 917-924. DOI:10.1016/j.scitotenv.2008.09.014 |
| [7] | Bouffard A, Amyot M. Importance of elemental mercury in lake sediments[J]. Chemosphere, 2009, 74(8): 1098-1103. DOI:10.1016/j.chemosphere.2008.10.045 |
| [8] | Obrist D, Faïn X, Berger C. Gaseous elemental mercury emissions and CO2 respiration rates in terrestrial soils under controlled aerobic and anaerobic laboratory conditions[J]. Scienceof the Total Environment, 2010, 408(7): 1691-1700. DOI:10.1016/j.scitotenv.2009.12.008 |
| [9] | Lindberg S E, Dong W J, Chanton J, et al. A mechanism for bimodal emission of gaseous mercury from aquatic macrophytes[J]. Atmospheric Environment, 2005, 39(7): 1289-1301. DOI:10.1016/j.atmosenv.2004.11.006 |
| [10] | Lindberg S E, Dong W J, Meyers T. Transpiration of gaseous elemental mercury through vegetation in a subtropical wetland in Florida[J]. Atmospheric Environment, 2002, 36(33): 5207-5219. DOI:10.1016/S1352-2310(02)00586-1 |
| [11] | Chen Z, Tang Y T, Zhou C, et al. Mechanisms of Fe biofortification and mitigation of Cd accumulation in rice (Oryza sativaL.) grown hydroponically with Fe chelate fertilization[J]. Chemosphere, 2017, 175: 275-285. DOI:10.1016/j.chemosphere.2017.02.053 |
| [12] | Colmer T D. Aerenchyma and an inducible barrier to radial oxygen loss facilitate root aeration in upland, paddy and deep-water rice (Oryza sativa L.)[J]. Annals of Botany, 2003, 91(2): 301-309. DOI:10.1093/aob/mcf114 |
| [13] |
戴志聪, 杜道林, 司春灿, 等. 用扫描仪及Image J软件精确测量叶片形态数量特征的方法[J]. 广西植物, 2009, 29(3): 342-347. Dai Z C, Du D L, Si C C, et al. A method to exactly measure the morphologicalquantity of leaf using scanner and Image J software[J]. Guihaia, 2009, 29(3): 342-347. |
| [14] |
邓东周, 范志平, 王红, 等. 林木蒸腾作用测定和估算方法[J]. 生态学杂志, 2008, 27(6): 1051-1058. Deng D Z, Fan Z P, Wang H, et al. Determination and estimation methods of tree transpiration:a review[J]. Chinese Journal of Ecology, 2008, 27(6): 1051-1058. |
| [15] |
马玲, 赵平, 饶兴权, 等. 乔木蒸腾作用的主要测定方法[J]. 生态学杂志, 2005, 24(1): 88-96. Ma L, Zhao P, Rao X Q, et al. Main determination methods of tree transpiration[J]. Chinese Journal of Ecology, 2005, 24(1): 88-96. |
| [16] |
傅志强, 黄璜, 何保良, 等. 水稻植株通气系统与稻田CH4排放相关性研究[J]. 作物学报, 2007, 33(9): 1458-1467. Fu Z Q, Huang H, He B L, et al. Correlation between rice plant aerenchymasystem and methane emission from paddy field[J]. Acta AgronomicaSinica, 2007, 33(9): 1458-1467. |
| [17] | Abiko T, Obara M. Enhancement of porosity and aerenchyma formation in nitrogen-deficient rice roots[J]. Plant Science, 2014, 215-216: 76-83. DOI:10.1016/j.plantsci.2013.10.016 |
| [18] | Drew M C, He C J, Morgan P W. Programmed cell death and aerenchyma formation in roots[J]. Trends in Plant Science, 2000, 5(3): 123-127. DOI:10.1016/S1360-1385(00)01570-3 |
| [19] | Butterbach-Bahl K, Papen H, Rennenberg H. Impact of gas transport through rice cultivars on methane emission from rice paddy fields[J]. Plant, Cell & Environment, 1997, 20(9): 1175-1183. |
| [20] |
樊明寿, 张福锁. 植物通气组织的形成过程和生理生态学意义[J]. 植物生理学通讯, 2002, 38(6): 615-618. Fan M S, Zhang F S. Aerenchyma formation in plant and its physiological and ecological significance[J]. Plant Physiology Communications, 2002, 38(6): 615-618. |
| [21] |
孔妤, 王忠, 顾蕴洁, 等. 植物根内通气组织形成的研究进展[J]. 植物学通报, 2008, 25(2): 248-253. Kong Y, Wang Z, Gu Y J, et al. Research progress on aerenchyma formation in plant roots[J]. Chinese Bulletin of Botany, 2008, 25(2): 248-253. |
| [22] | Greger M, Wang Y D, Neuschütz C. Absence of Hg transpiration by shoot after Hg uptake by roots of six terrestrial plant species[J]. Environmental Pollution, 2005, 134(2): 201-208. DOI:10.1016/j.envpol.2004.08.007 |
| [23] | Du X, Zhu Y G, Liu W J, et al. Uptake of mercury (Hg) by seedlings of rice (Oryza sativaL.) grown in solution culture and interactions with arsenate uptake[J]. Environmental and Experimental Botany, 2005, 54(1): 1-7. DOI:10.1016/j.envexpbot.2004.05.001 |
| [24] | 江苏农学院. 植物生理学[M]. 北京: 农业出版社, 1986: 102-103. |
| [25] | Borah L, Baruah K K. Effects of foliar application of plant growth hormone on methane emission from tropical rice paddy[J]. Agriculture, Ecosystems & Environment, 2016, 233: 75-84. |
| [26] | Philippot L, Hallin S. Towards food, feed and energy crops mitigating climate change[J]. Trends in Plant Science, 2011, 16(9): 476-480. DOI:10.1016/j.tplants.2011.05.007 |
| [27] | Battke F, Ernst D, Fleischmann F, et al. Phytoreduction and volatilization of mercury by ascorbate in Arabidopsis thaliana, European beech and Norway spruce[J]. Applied Geochemistry, 2008, 23(3): 494-502. DOI:10.1016/j.apgeochem.2007.12.023 |
| [28] | Leonard T L, TaylorJr G E, Gustin M S, et al. Mercury and plants in contaminated soils:1. Uptake, partitioning, and emission to the atmosphere[J]. Environmental Toxicology and Chemistry, 1998, 17(10): 2063-2071. DOI:10.1002/etc.v17:10 |