 2017, Vol. 38
2017, Vol. 38



厌氧氨氧化是指在厌氧条件下以NH4+-N作为电子供体, NO2--N作为电子受体, 将NH4+-N和NO2--N转化为N2的生物氧化过程[1].自从Thamdrup等[2]第一次在海洋沉积物中发现厌氧氨氧化反应后, 海洋厌氧氨氧化菌开始引起学者们的关注, 由于其不仅可以在高盐条件下有着高氨去除的潜力, 而且能够直接说明海洋环境内的氮循环, 因此研究者们相继在黑海、太平洋等海洋环境下发现厌氧氨氧化反应[3~5].然而, 海洋厌氧氨氧化菌的种属较少以及生长速率较慢成为研究海洋厌氧氨氧化工艺的主要障碍[6].目前为止, 有关海洋厌氧氨氧化工艺处理含盐废水的情况鲜见报道.
为了能够维持较高的脱氮效能, 基质NH4+-N和NO2--N的浓度应在适当的范围内, 基质浓度还应符合合理的化学计量比, 一旦NH4+-N和NO2--N的浓度较高, 两者将作为抑制剂影响厌氧氨氧化菌的生长和脱氮能力.针对淡水厌氧氨氧化菌的研究中, 很多学者得到了不同的实验结果.其中NH4+-N的抑制作用主要是游离氨(FA)浓度, Fernández等[7]的研究表明, 淡水厌氧氨氧化菌长期暴露于FA浓度在35~40 mg·L-1的条件下时, 反应性能极度不稳定, 去除效率接近零; Waki等认为[8], 当FA浓度在13~90 mg·L-1时, 便成为影响厌氧氨氧化反应器的抑制因素; Tang等[9]的研究结果则显示, FA浓度在57~187 mg·L-1时影响厌氧氨氧化菌的脱氮效能.而NO2--N的毒性作用要强于NH4+-N[10, 11], Fux等[12]发现长期暴露在NO2--N浓度为40 mg·L-1的条件下, 会导致厌氧氨氧化菌不可逆的失活[13]; Isaka等[10]则认为NO2--N浓度超过280 mg·L-1时开始抑制厌氧氨氧化反应器; Dapena-Mora等[11]的研究结果表明比厌氧氨氧化活性(SAA)在NO2--N浓度为350 mg·L-1时降低50%.
以上研究结果是在淡水厌氧氨氧化反应器的条件下得到的, 而海洋厌氧氨氧化菌在高盐条件下, 这两种反应基质对其抑制影响未有报道.鉴于此, 本研究通过固定一种反应基质, 逐渐增加另一种反应基质浓度的方式, 分别探讨了NH4+-N和NO2--N对海洋厌氧氨氧化菌的影响, 并利用基质抑制动力学分析了反应器稳定运行的策略, 以期为海洋厌氧氨氧化菌处理含海水污水的脱氮性能提供理论依据.
1 材料与方法 1.1 实验装置本研究采用ASBR反应器(如图 1), 其有效容积为7 L, 系有机玻璃制成.反应器内的温度通过温控箱控制在25℃左右, 整个反应器用锡纸包裹, 防止光对海洋厌氧氨氧化菌活性的影响.反应区内悬挂无纺滤布作为海洋厌氧氨氧化菌的生物膜载体, 且置有电动搅拌器, 反应器内污泥主要以红色颗粒为主.实验采用人工配制的模拟废水, 经高纯氮气吹脱15 min, 消除溶解氧的影响, 再从反应器下部进水口通过流动泵进水.运行一个周期包括进水4 min, 反应6 h, 静置25 min, 出水4 min.

|
图 1 海洋厌氧氨氧化反应器示意 Fig. 1 Schematic diagram of the marine ANAMMOX reactor |
实验所用海水取自胶州湾(黄海北部), 平均盐度为32‰, 经人工配制成模拟废水, 其主要成分为:29 mg·L-1 KH2PO4, 136 mg·L-1 CaCl2, 1 200 mg·L-1 KHCO3, 300 mg·L-1 MgSO4·7H2O.同时投加微生物所需的微量元素, 微量元素Ⅰ:EDTA 5 000 mg·L-1, FeSO4·7H2O 5 000 mg·L-1, 微量元素Ⅱ:EDTA 15 000 mg·L-1, H3BO3 11 mg·L-1, MnCl2·4H2O 990 mg·L-1, CuSO4·5H2O 250 mg·L-1, ZnSO4·7H2O 430 mg·L-1, NiCl2·6H2O 190 mg·L-1, Na2MoO4·2H2O 220 mg·L-1, CoCl2·6H2O 240 mg·L-1, NaSeO4·10H2O 210 mg·L-1. NH4+-N和NO2--N分别用NH4Cl和NaNO2提供.
1.3 分析方法NH4+-N:纳氏试剂比色法[14]; NO2--N:N-(1-萘基)-乙二胺光度法[14]; NO3--N:麝香草酚分光光度法[14]; pH值:WTW pH/Oxi 340i;温度:水银温度计; 紫外/可见光分光光度计:UV-5200.总氮容积负荷(TNLR)根据进水总氮浓度与HRT关系得到, 总氮去除负荷(TNRR)根据进、出水总氮浓度与HRT关系得到, 总氮去除率(TNRE)根据进、出水总氮浓度的关系得到, 游离氨(FA)和游离亚硝酸(FNA)根据He等[15]的研究, 即pH值与进、出水氨氮、亚硝态氮浓度的关系得到.

|
(1) |
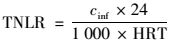
|
(2) |

|
(3) |

|
(4) |

|
(5) |
式中, cinf为进水总氮浓度(mg·L-1), ceff为出水总氮浓度(mg·L-1), HRT为水力停留时间(HRT=6 h, 采样间隔t=0.5 h), FA为游离氨浓度(mg·L-1), TAN为总氨浓度(NH4+-N+NH3),
FNA为游离亚硝酸(mg·L-1), TNN为总亚硝态氮(NO2--N+HNO2--N).
1.4 动力学模型Haldane模型和Aiba模型都是用来描述基质抑制动力学的.目前被广泛应用于描述厌氧氨氧化反应器的基质抑制动力学特性[16, 17].
Haldane模型:

|
(6) |
Aiba模型:

|
(7) |
式中, TNRR为总氮基质去除负荷[kg·(m3·d)-1], TNRRmax为最大基质去除负荷[kg·(m3·d)-1], KS为半饱和常数(mg·L-1), S为基质浓度(mg·L-1), KiA为Haldane抑制动力学常数(mg·L-1), KiB为Aiba抑制动力学常数(mg·L-1).
2 结果与讨论 2.1 基质NH4+-N对海洋厌氧氨氧化菌脱氮性能的影响该阶段NO2--N浓度固定为105.6 mg·L-1, 逐渐提高进水NH4+-N浓度, 实验结果如图 2所示.由图 2(a)~2(c)可知, NH4+-N浓度在780mg·L-1以下时, NO2--N的平均去除率基本稳定在97%以上, 虽然图 2(c)中NH4+-N去除率在该浓度范围内逐渐下降, 是由于剩余的NH4+-N浓度导致.同时由图 2(d)所示, ΔNO2--N:ΔNH4+-N=1.25±0.25, ΔNO3--N:ΔNH4+-N=0.19±0.07, 说明此时反应器内海洋厌氧氨氧化菌的脱氮能力几乎没有受到影响.

|
图 2 NH4+-N浓度对反应器内脱氮指标的影响 Fig. 2 Effects of NH4+-N concentration on the nitrogen removal index of the reactor |
随着进一步提高进水NH4+-N浓度, 出水NO2--N浓度出现轻微变化.当NH4+-N浓度为980 mg·L-1时, 游离氨(FA)的平均浓度为21.70 mg·L-1, 出水NO2--N的平均浓度为13.22 mg·L-1, 此时TNLR的平均值为4.38 kg·(m3·d)-1, TNRR的平均值为0.62kg·(m3·d)-1, 且由图 2(d)能够看出ΔNO2--N:ΔNH4+-N=1.30±0.15, ΔNO3--N:ΔNH4+-N=0.19±0.03.当进水NH4+-N浓度为1 100 mg·L-1时, TNLR的平均值上升至4.86 kg·(m3·d)-1, FA平均浓度为24.39mg·L-1, NO2--N的平均出水浓度为17.77 mg·L-1, TNRR的平均值为0.61kg·(m3·d)-1, ΔNO2--N:ΔNH4+-N=1.08±0.12, ΔNO3--N:ΔNH4+-N=0.16±0.02.而在进水NH4+-N浓度提高至1 200 mg·L-1时, TNLR的平均值提高至5.25kg·(m3·d)-1, FA平均浓度为26.57mg·L-1, 此时NO2--N的出水浓度又出现较小幅度的增加, 其平均值为19.75 mg·L-1, TNRR的平均值为0.59 kg·(m3·d)-1, ΔNO2--N:ΔNH4+-N=1.09±0.20, ΔNO3--N:ΔNH4+-N =0.14±0.03.由此看出, 较高的NH4+-N浓度对反应器的影响不太明显, 海洋厌氧氨氧化菌具有较强的耐氨能力.
李祥等[18]利用回流装置研究了NH4+-N浓度对其氮素转化特性的影响, 实验结果显示当进水NH4+-N浓度上升到900 mg·L-1时, 厌氧氨氧化反应器出现了NO2--N积累的现象, 而厌氧氨氧化细菌的活性未出现明显的抑制现象.与其不同的是, 本实验中未采用回流部分且进水NH4+-N浓度达到1 200 mg·L-1仍然保持较高的脱氮效率, 由此可知, 研究结果的不同与反应器装置设置和种群特性密切相关.
NH4+和NH3在水中存在一种化学平衡:NH4++OH-=NH3+H2O, 进水NH4+-N浓度逐渐增加, FA浓度也随之增加, 而FA进入微生物细胞内会改变细胞环境的pH值, 细胞内外的pH值不同, 从而影响跨膜电位, 甚至会导致细胞死亡[19].本实验中, 通过控制pH值, 同时逐渐提高进水NH4+-N浓度, 在FA浓度为17.32 mg·L-1时, 反应器开始出现轻微的影响, 而后NH4+-N浓度提高至1 200 mg·L-1, FA浓度提高26.57mg·L-1, 虽然出水NO2--N浓度略有提高, 而海洋厌氧氨氧化反应器仍表现出较好的活性.该结果与Fernández等[7]的研究相类似, 其认为在长期实验中, 当FA浓度在20~25 mg·L-1范围内时, 反应器开始不稳定, 而厌氧氨氧化污泥长期处于FA浓度为35~40 mg·L-1时, 其脱氮效率下降为零.本实验中, 当FA浓度上升至21.70 mg·L-1时, 出水NO2--N的浓度出现轻微的提高, 而FA最终增加至26.57mg·L-1时, 出水NO2--N浓度的变化较小, 因此, 海洋厌氧氨氧化反应器不仅有较强的耐氨能力, 而且也会逐渐适应FA浓度, 此外, 不同厌氧氨氧化种群的生理特征也可能会有助于其在反应器内适应FA的抑制作用.
2.2 基质NO2--N对海洋厌氧氨氧化菌脱氮性能的影响该阶段NH4+-N浓度固定为80 mg·L-1, 逐渐提高进水NO2--N浓度.实验结果如图 3(a)~3(c)所示.当进水NO2--N浓度为125.6 mg·L-1时, NH4+-N基本可以完全去除, 反应器几乎没有影响, 此时TNRR为(0.60±0.03)kg·(m3·d)-1.随着进水NO2--N浓度进一步提高至145.6 mg·L-1和185.6 mg·L-1时, 出水NH4+-N浓度出现轻微的变化, 但NH4+-N去除率基本维持在80%左右, 平均TNRR分别为(0.53±0.06)kg·(m3·d)-1、(0.57±0.04)kg·(m3·d)-1, 同时通过图 3(d)可知, ΔNO2--N:ΔNH4+-N=1.27±0.11, ΔNO3--N:ΔNH4+-N=0.26±0.02, 说明此时反应器比较稳定, 进水NO2--N浓度对反应器影响较小.

|
图 3 NO2--N浓度对反应器内脱氮指标的影响 Fig. 3 Effects of NO2--N concentration on the nitrogen removal index of the reactor |
然而, 当进水NO2--N浓度提高至225.6 mg·L-1时, 游离亚硝酸(FNA)的平均浓度为0.04 mg·L-1, 海洋厌氧氨氧化菌的活性开始受到明显的抑制作用, 出水NH4+-N浓度增加, 其平均去除率为73.61%, TNRR下降至(0.48±0.11)kg·(m3·d)-1, ΔNO2--N:ΔNH4+-N=1.13±0.22, ΔNO3--N:ΔNH4+-N=0.22±0.04.这可以认为, 该进水NO2--N浓度是开始抑制反应器脱氮效能的临界条件.而后进水NO2--N浓度继续增加至265.6 mg·L-1, FNA的平均浓度也随之提高至0.05 mg·L-1, 出水NH4+-N的平均浓度增加至31.42mg·L-1, 其去除率下降为59.88%, TNRR为(0.42±0.07)kg·(m3·d)-1.当进水NO2--N浓度为305.6 mg·L-1时, FNA的平均浓度提高至0.06 mg·L-1, 海洋厌氧氨氧化反应器的脱氮效能进一步下降, 出水NH4+-N浓度增加到44.93 mg·L-1, 去除率仅为43.93%, TNRR下降至(0.32±0.16)kg·(m3·d)-1.
针对厌氧氨氧化反应器起到主要抑制作用的是FNA, 还是亚硝酸盐, 学者们还未统一观点.一直以来, 大多数的观点认为FNA起到主要的抑制作用[7, 20], 而Puyol等认为[21], 当pH低于7.1时, 起到主要抑制作用的是FNA, 而pH高于7.1时, 亚硝酸盐起到主要的抑制作用.本实验中, 反应器内的进水pH值始终维持在7.5左右, 因此, 可以认为在本实验中主要抑制反应器脱氮效能的是亚硝酸盐.
由于NO2--N的生物毒性作用, 其浓度超过一定阈值会明显影响厌氧氨氧化反应器的脱氮能力, 与NH4+-N的抑制阈值相比, NO2--N的抑制阈值要低很多, 因此, 反应器更容易受到NO2--N的抑制作用[11, 22]. Kimura等[23]的研究表明, 当进水NO2--N浓度从750 mg·L-1降低至274 mg·L-1时, 厌氧氨氧化菌活性可以充分恢复.由此可以看出, NO2--N对厌氧氨氧化反应器的抑制作用可逆[24], 而NH4+-N的存在对反应器起到一定的保护作用, 使其对NO2--N的抑制作用具有较高的耐受性[25].在很多研究中都高估了NO2--N的抑制作用, 较高的NO2--N浓度未必是影响反应器稳定运行的原因[26].抑制反应器稳定运行的主要因素是游离亚硝酸(FNA), 随着逐渐提高进水NO2--N浓度, FNA的浓度也随之变化, 其进入细胞质内改变了细胞环境内的pH值[27].因此, 为了反应器的稳定运行, 若单独提高NO2--N浓度时应控制在265.6 mg·L-1以下.
2.3 基质NH4+-N和NO2--N的抑制动力学特性利用Haldane模型和Aiba模型分析进水NH4+-N和NO2--N浓度对海洋厌氧氨氧化反应器的抑制特性.如图 4所示.在图 4(a)~4(b)中以出水NH4+-N浓度为横坐标, TNRR为纵坐标, 得到NH4+-N浓度抑制的动力学曲线.通过相关系数R2可以看出, 两个模型均可以比较好地拟合进水NH4+-N浓度对海洋厌氧氨氧化反应器的抑制动力学特性.从拟合得到的参数来看, Haldane模型得到的TNRRmax为0.697 kg·(m3·d)-1, KS为1.416 mg·L-1, KiA为3 893.625 mg·L-1, 而Aiba模型得到的TNRRmax为0.694 kg·(m3·d)-1, KS为1.309 mg·L-1, KiB为4 394.010 mg·L-1, 两者得到的TNRRmax和KS的值相似, 但是却与实际值存在一定的误差, 尤其是TNRRmax的相对误差大约为3.59%.两个模型拟合得到的Ki比较高, 远高于祖波等[28]报道的抑制常数1 123 mg·L-1.在本实验中, 反应器在高氨氮浓度下仍具保持较好地脱氮能力, 这可能与反应器内菌种特性有关, 海洋厌氧氨氧化菌可能对NH4+-N的耐受性较高, 并且对其具有良好的适应性.

|
图 4 Haldane模型和Aiba模型拟合曲线 Fig. 4 Fitted curves for the Haldane model and Aiba model |
在图 4(c)~4(d)中以出水NO2--N浓度为横坐标, 以TNRR为纵坐标, 得到NO2--N浓度抑制的动力学曲线.通过相关系数R2可知, 两个模型可以较好地模拟进水NO2--N浓度对海洋厌氧氨氧化反应器的抑制动力学特性.同时通过拟合得到的参数来看, Haldane模型得到的TNRRmax为0.690 kg·(m3·d)-1, KS为0.223 mg·L-1, KiB为287.208 mg·L-1, 而Aiba模型得到的TNRRmax为0.667 kg·(m3·d)-1, KS为0.115 mg·L-1, KiB为398.143 mg·L-1, 两者得到的3个参数值略有不同, Aiba模型得到的TNRRmax值更接近于实际值, 相对误差为3.97%, 而Haldane模型给出的TNRRmax值的相对误差为7.17%, 如果使用Aiba模型, 拟合得到的抑制常数KiB(398.143 mg·L-1)高于李伟刚等[29]得到的抑制常数(140.60 mg·L-1), 却远远低于金仁村[30]得到的NO2--N抑制常数(924 mg·L-1).这些研究结果的不同一方面与反应器内优势种群有关, 另一方面有报道说明, 在高盐环境下, 厌氧氨氧化菌可以产生更多的胞外聚合物来抵御极端环境[31, 32].因此, 对于海洋厌氧氨氧化菌来说可能自身可以阻挡部分NO2--N的抑制作用.
选取Haldane模型和Aiba模型进行对比, 主要是寻找更能够精确描述基质抑制对海洋厌氧氨氧化菌脱氮特性.因此, 对于Haldane模型和Aiba模型分别求导, 令两者的一阶导数为0, 最后将S用KS、Ki表示分别代回式(6)、式(7), 得到如下公式:

|
(8) |

|
(9) |
通过拟合得到的3个参数值代入式(8)、(9)分别得到Haldane模型和Aiba模型下NH4+-N和NO2--N抑制作用的最大总氮去除负荷(TNRRmax). Haldane模型下对于NH4+-N浓度对反应器脱氮能力的影响得到TNRRmax为0.671 kg·(m3·d)-1, 而Aiba模型下得到的TNRRmax为0.699 kg·(m3·d)-1; Haldane模型下对于NO2--N浓度对反应器脱氮能力的影响得到TNRRmax为0.654 kg·(m3·d)-1, 而Aiba模型下得到的TNRRmax为0.645 kg·(m3·d)-1.同时, 实验测得的NH4+-N和NO2--N抑制作用的最大总氮去除负荷分别为0.672 kg·(m3·d)-1和0.641 kg·(m3·d)-1, 由此可以看出:对于NH4+-N的抑制作用, Haldane模型下得到的TNRRmax与实际测量值更为接近, 其更适合描述氨氮抑制海洋厌氧氨氧化反应器的动力学特性; 而对于NO2--N抑制作用, Aiba模型下得到的TNRRmax与实际测量值更贴近, 其更适合描述亚硝态氮抑制海洋厌氧氨氧化反应器的动力学特性, 两者预测得到的TNRRmax值与实际值的相对误差较小.因此, 该结果为反应器的稳定运行提供了较好的参考价值.
3 结论(1) 当进水NH4+-N浓度提高至1 200 mg·L-1时, 海洋厌氧氨氧化反应器仍然具有较高的脱氮能力, NH4+-N对其抑制作用并不明显, 说明与淡水属厌氧氨氧化菌相比, 海洋厌氧氨氧化菌具有更强的耐氨能力.
(2) 当进水NO2--N浓度提高至265.6 mg·L-1时, 反应器脱氮能力明显被抑制, NH4+-N的去除率下降为59.88%, 相比于NH4+-N而言, 海洋厌氧氨氧化菌对NO2--N的抑制作用较为敏感.
(3) 通过Haldane模型和Aiba模型的拟合可知, Haldane模型更适合描述NH4+-N对反应器的抑制动力学特性, 而Aiba模型更适合描述NO2--N对反应器的抑制动力学特性.
| [1] |
曹天昊, 王淑莹, 苗蕾, 等. 不同基质浓度下SBR进水方式对厌氧氨氧化的影响[J]. 中国环境科学, 2015, 35(8): 2334-2341. Cao T H, Wang S Y, Miao L, et al. Influence of feeding modes on anammox under different influent substrate concentration in SBR[J]. China Environmental Science, 2015, 35(8): 2334-2341. |
| [2] | Thamdrup B, Dalsgaard T. Production of N2 through anaerobic ammonium oxidation coupled to nitrate reduction in marine sediments[J]. Applied and Environmental Microbiology, 2002, 68(3): 1312-1318. DOI:10.1128/AEM.68.3.1312-1318.2002 |
| [3] | Kuypers M M M, Sliekers A O, Lavik G, et al. Anaerobic ammonium oxidation by anammox bacteria in the Black Sea[J]. Nature, 2003, 422(6932): 608-611. DOI:10.1038/nature01472 |
| [4] | Rysgaard S, Glud R N, Risgaard-Petersen N, et al. Denitrification and anammox activity in Arctic marine sediments[J]. Limnology Oceanography, 2004, 49(5): 1493-1502. DOI:10.4319/lo.2004.49.5.1493 |
| [5] | Shehzad A, Liu J W, Yu M, et al. Diversity, community composition and abundance of anammox bacteria in sediments of the north marginal seas of China[J]. Microbes and Environments, 2016, 31(2): 111-120. DOI:10.1264/jsme2.ME15140 |
| [6] | Strous M, Heijnen J J, Kuenen J G, et al. The sequencing batch reactor as a powerful tool for the study of slowly growing anaerobic ammonium-oxidizing microorganisms[J]. Applied Microbiology and Biotechnology, 1998, 50(6): 589-596. |
| [7] | Fernández I, Dosta J, Fajardo C, et al. Short-and long-term effects of ammonium and nitrite on the anammox process[J]. Journal of Environmental Management, 2012, 95(S1): S170-S174. |
| [8] | Waki M, Tokutomi T, Yokoyama H, et al. Nitrogen removal from animal waste treatment water by anammox enrichment[J]. Bioresource Technology, 2007, 98(14): 2775-2780. DOI:10.1016/j.biortech.2006.09.031 |
| [9] | Tang C J, Zheng P, Mahmood Q, et al. Effect of substrate concentration on stability of anammox biofilm reactors[J]. Journal of Central South Universal Technology, 2010, 17(1): 79-84. DOI:10.1007/s11771-010-0014-6 |
| [10] | Isaka K, Sumino T, Tsuneda S. High nitrogen removal performance at moderately low temperature utilizing anaerobic ammonium oxidation reactions[J]. Journal of Bioscience and Bioengineering, 2007, 103(5): 486-490. DOI:10.1263/jbb.103.486 |
| [11] | Dapena-Mora A, Fernández I, Campos J L, et al. Evaluation of activity and inhibition effects on anammox process by batch tests based on the nitrogen gas production[J]. Enzyme and Microbial Technology, 2007, 40(4): 859-865. DOI:10.1016/j.enzmictec.2006.06.018 |
| [12] | Fux C. Biological nitrogen elimination of ammonium-rich sludge digester liquids[D]. Switzerland:ETH-Zürich, 2003. |
| [13] | van Hulle S W H, Vandeweyer H J P, Meesschaert B D, et al. Engineering aspects and practical application of autotrophic nitrogen removal from nitrogen rich streams[J]. Chemical Engineering Journal, 2010, 162(1): 1-20. DOI:10.1016/j.cej.2010.05.037 |
| [14] | 国家环境保护总局. 水和废水监测分析方法[M]. 第四版. 北京: 中国环境科学出版社, 2002. |
| [15] | He S L, Niu Q G, Ma H Y, et al. The treatment performance and the bacteria preservation of anammox:a review[J]. Water, Air, & Soil Pollution, 2015, 226: 163. |
| [16] | Chen T T, Zheng P, Shen L D, et al. Kinetic characteristics and microbial community of anammox-EGSB reactor[J]. Journal of Hazardous Materials, 2011, 190(1-3): 28-35. DOI:10.1016/j.jhazmat.2010.12.060 |
| [17] | Jin R C, Xing B S, Yu J J, et al. The importance of the substrate ratio in the operation of the anammox process in upflow biofilter[J]. Ecologyical Engineering, 2013, 53: 130-137. DOI:10.1016/j.ecoleng.2012.12.027 |
| [18] |
李祥, 崔剑虹, 袁砚, 等. 高氨氮对具有回流的PN-ANAMMOX串联工艺的脱氮影响[J]. 环境科学, 2015, 36(10): 3749-3755. Li X, Cui J H, Yuan Y, et al. Effect of high ammonium on nitrogen removal in an partial nitritation-ANAMMOX process with reflux system[J]. Environmental Science, 2015, 36(10): 3749-3755. |
| [19] | Martinelle K, Westlund A, Häggstr m L. Ammonium ion transport-a cause of cell death[J]. Cytotechnology, 1996, 22(1-3): 251-254. DOI:10.1007/BF00353945 |
| [20] | Egli K, Fanger U, Alvarez P J J, et al. Enrichment and characterization of an anammox bacterium from a rotating biological contactor treating ammonium-rich leachate[J]. Archives of Microbiology, 2001, 175(3): 198-207. DOI:10.1007/s002030100255 |
| [21] | Puyol D, Carvajal-Arroyo J, Sierra-Alvarez R, et al. Nitrite (not free nitrous acid) is the main inhibitor of the anammox process at common pH conditions[J]. Biotechnology Letters, 2014, 36(3): 547-551. DOI:10.1007/s10529-013-1397-x |
| [22] | Bettazzi E, Caffaz S, Vannini C, et al. Nitrite inhibition and intermediates effecte on Anammox bacteria:A batch-scale experimental study[J]. Process Biochemistry, 2010, 45(4): 573-580. DOI:10.1016/j.procbio.2009.12.003 |
| [23] | Kimura Y, Isaka K, Kazama F, et al. Effects of nitrite inhibition on anaerobic ammonium oxidation[J]. Applied Microbiology and Biotechnology, 2010, 86(1): 359-365. DOI:10.1007/s00253-009-2359-z |
| [24] | Lotti T, van der Star W R L, Kleerebezem R, et al. The effect of nitrite inhibition on the anammox process[J]. Water Research, 2012, 46(8): 2559-2569. DOI:10.1016/j.watres.2012.02.011 |
| [25] | Carvajal-Arroyo J M, Puyol D, Li G, et al. Pre-exposure to nitrite in the absence of ammonium strongly inhibits anammox[J]. Water Research, 2014, 48: 52-60. DOI:10.1016/j.watres.2013.09.015 |
| [26] | Jaroszynski L W, Cicek N, Sparling R, et al. Importance of the operating pH in maintaining the stability of anoxic ammonium oxidation (anammox) activity in moving bed biofilm reactors[J]. Bioresource Technology, 2011, 102(14): 7051-7056. DOI:10.1016/j.biortech.2011.04.069 |
| [27] | Sprott G D, Patel G B. Ammonia toxicity in pure cultures of methanogenic bacteria[J]. Systematic and Applied Microbiology, 1986, 7(2-3): 358-363. DOI:10.1016/S0723-2020(86)80034-0 |
| [28] |
祖波, 张代钧, 阎青. 颗粒污泥厌氧氨氧化动力学特性及微量NO2的影响[J]. 环境科学, 2008, 29(3): 683-687. Zu B, Zhang D J, Yan Q. Effect of trace NO2 and kinetic characteristics for anaerobic ammonium oxidation of granular sludge[J]. Environmental Science, 2008, 29(3): 683-687. |
| [29] |
李伟刚, 于德爽, 李津. ASBR反应器厌氧氨氧化脱氮Ⅱ:反应动力学[J]. 中国环境科学, 2013, 33(12): 2191-2200. Li W G, Yu D S, Li J. Nitrogen removal in the anammox sequencing batch reactor Ⅱ:kinetics characteristics[J]. China Environmental Science, 2013, 33(12): 2191-2200. |
| [30] | 金仁村. 自养型生物脱氮反应器性能的研究[D]. 杭州: 浙江大学, 2007. http://d.wanfangdata.com.cn/Thesis/Y1124640 |
| [31] | Jin R C, Ma C, Yu J J. Performance of an anammox UASB reactor at high load and low ambient temperature[J]. Chemical Engineering Journal, 2013, 232: 17-25. DOI:10.1016/j.cej.2013.07.059 |
| [32] | Ismail S B, De La Parra C J, Temmink H, et al. Extracellular polymeric substances (EPS) in upflow anaerobic sludge blanket (UASB) reactors operated under high salinity conditions[J]. Water Research, 2010, 44(6): 1909-1917. DOI:10.1016/j.watres.2009.11.039 |