 2017, Vol. 38
2017, Vol. 38



随着我国社会经济的快速发展, 大量富含氮的工业、农业及城市生活废水进入水环境, 导致水体富营养化, 严重破坏地表水环境[1, 2].贡湖作为太湖的重要组成部分, 由于周边城市化进程加快, 污染排放加剧, 正面临着水生生态系统严重退化、生物多样性减少及污染净化能力下降等问题[3].亲水河作为流入贡湖的主要河道, 水体氮素含量的升高, 会对贡湖水生生态系统的生物多样性及水质稳定性造成严重影响.因此, 采取合适的方法降低入贡湖河道水体氮素营养盐浓度, 具有重要意义.
近年来, 国内外学者均对河道水体富营养化的控制及氮素营养盐脱除机制等方面做了大量研究.李宝珍等[4]对植物吸收利用铵态氮和硝态氮分子调控及对氮素吸收优化配置进行了研究.刘丹丹等[5]研究了伊乐藻和氮循环菌联用技术对于太湖氮素营养盐的吸收和反硝化影响.韩华杨等[6]探讨了底泥反硝化及厌氧氨氧化细菌对河道底泥氮素营养盐去除贡献. Peipoch等[7]进行了河岸水生植物对硝态氮和铵态氮的吸收能力对比研究.
然而, 以往的许多研究主要集中在水体氮素的去除效果, 并未对各氮素脱除途径进行定量地分析, 在伊乐藻-脱氮微生物联用系统中, 有关底泥存储、水生植物吸收与微生物过程对于氮素去除的定量贡献研究尚不多见[8, 9].
因此, 本研究主要针对亲水河氮素营养盐污染负荷高、水体流速趋于静止、含氧量低等特点, 运用固定化高效脱氮微生物(immobilized nitrogen cycling bacteria, INCB)与沉水植物伊乐藻(Elodea nuttallii)相结合的生态修复技术, 在室内进行原位模拟实验, 通过提高污染水体的氮素转化能力进而达到修复河道的目的.与此同时, 本研究还探讨了在脱氮微生物与伊乐藻联用系统下, 底泥存储、水生植物吸收及微生物过程3种途径对污染水体氮素脱除机制的影响.
1 材料与方法 1.1 采样点与样品采集2015年4月于入贡湖河道亲水河(32°17′52″N, 120°20′20″E, 图 1)采集水样、伊乐藻及底泥样品.为确保模拟实验准确性, 样品采集前根据亲水河污染及人为干扰情况, 选取未受干扰段河道进行样品采集工作.采用特制柱状采泥器(有机玻璃柱长度100 cm, 内径9 cm)采集24根完整泥柱样品, 底泥的取样深度约40 cm.所有采集的底泥、水样及伊乐藻样品于4 h内送至实验室. 2015年4月~2015年10月的水质长期跟踪监测资料显示, 亲水河上覆水中总氮浓度(TN)2.5~3.5 mg·L-1, 总磷浓度(TP)0.1~0.2 mg·L-1, 铵态氮浓度(NH4+-N)0.5~1.5mg·L-1, 硝态氮浓度(NO3--N)0.5~1.0mg·L-1, 底泥样品采集区域表层底泥(表层5cm)相关理化参数初始值见表 1.

|
图 1 太湖贡湖湾入湖河道-亲水河采样点地理位置示意 Fig. 1 Geographical location of sampling points in Qinshui River, Gonghu Bay, Taihu Lake |
|
|
表 1 实验区域表层底泥的物理化学参数(n=3) Table 1 Physiochemical characteristics of surface layer of the sampled sediment core |
1.2 固定高效脱氮微生物的制备与应用
实验采用甲基丙烯酸-β-羟乙酯(HEMA)、丙烯酸羟乙酯(HEA)按特定体积比与蒸馏水均匀混合, 在-78℃温度, 60Co-γ射线(吸收剂量1×104 Gy)条件下辐照制备具有生物相容性的固定化载体[10], 载体内具有的多孔结构, 可为高效脱氮微生物生长并进行繁殖提供微环境.
本研究采集亲水河底泥样品、水样及植物根区微生物样品, 经过多次富集、筛选分离得出土著脱氮微生物.通过连续培养, 将脱氮微生物固定于载体之上, 制备成固定化高效脱氮微生物.
1.3 室内实验设计将在亲水河采集的24根底泥柱被分成平行的6排, 每排分成4个不同的微环境处理系统, 分别为:处理组A, 未经处理的裸泥组; 处理组B, 仅添加INCB, 添加量为60 g±1 g; 处理组C, 仅添加伊乐藻, 每支截取长度为10 cm; 处理组D, 添加沉水植物伊乐藻和固定化脱氮微生物(E-INCB), 具体添加量如前所述.在室温约为20℃±1℃进行培养, 为期约6个月的室内培养结束后, 进行相关水质及理化数据的测定.
1.4 实验测试指标及方法 1.4.1 水质指标及测定方法铵态氮采用纳氏试剂分光光度法测定; 硝态氮采用紫外分光光度法测定; 总氮采用过硫酸钾氧化紫外分光光度法(日本岛津UV-2450) 测定; pH采用便携式pH计(YSI pH100) 进行测定; DO采用便携式溶氧仪(YSI 550A)进行测定.
1.4.2 N2排放通量测定经过为期6个月的室内培养, 实验柱内系统趋于稳定, 随后进行N2排放通量测定, 向3组平行的实验柱中加入同位素标记的15NH4+(15N at.%=99.11), 使得每个处理组中的15NH4+浓度约为100 μmol·L-1, 同位素添加之后, 将实验柱用橡胶塞密封, 随后在室温下(20℃±1℃)进行24 h无顶空密闭静态培养, 培养结束后, 用注射器采集水样, 溢流收集到密封良好的采集管中, 同时加入0.5 mL ZnCl2溶液, 用于抑制管中微生物活性, 恒温保存样品, 通过膜接口质谱仪(MIMS, Bay Instruments, Easton, MD, USA)对同位素其他28N2、29N2、30N2进行测定.
1.4.3 15N平衡模型实验为确保自动化C/N同位素比质谱仪测定结果的准确性, 需将系统初始15N丰度控制在10%以内.因此分别向3个平行组中加入同位素标记的15NH4+(15N at.%=99.11)500 μg, 每柱上部留有10 cm顶空用以进行气体采集, 用橡胶塞密封后, 在室温条件下(20℃±1℃)进行24 h培养, 培养时间结束后, 采集实验柱中气体、水体、植物及底泥样品, 用以测定15N丰度及相关理化性质.
1.4.4 样品采集与15N丰度测定水体15N丰度采用自动化C/N同位素比质谱仪(Europa Scientific Integra, Crewe, UK)测定.使用氧化镁戴氏合金法蒸馏分离NO3-和NH4+, 分别对15N丰度进行测定.在水样中加入氧化镁以蒸馏提取NH4+, 随后加入戴氏合金蒸馏分离出NO3-, 蒸馏气体用硼酸指示剂吸收, 使用0.01 mol·L-1硫酸溶液进行酸化, 将蒸馏出的NH3转化为含有(NH4)2SO4溶液, 于80℃烘箱烘干后进入同位素比质谱仪进行15N丰度的测定.
利用环形刀采集实验柱中底泥样品, 去除底泥中石块与植物根茎, 同时采集柱中伊乐藻样品, 用去离子水洗净表面泥土后, 将伊乐藻和底泥样品于80℃烘箱中烘至恒重, 使用研钵将干燥后的底泥样品研磨成均匀粉末, 将粉末样品过100目筛子.于万分之一天平称取定量底泥和植物样品, 随后使用同位素比质谱仪和在线元素分析仪(Europa Scientific Integra, Crewe, UK)测定样品15N丰度及氮素百分比.
1.4.5 N2O排放通量测定N2O排放通量采用静态箱-气相色谱法进行测定, 实验柱上部预设有高度10 cm的气体顶空收集室. 24 h室内培养结束后, 采用连有三通接头的注射器采集顶空气体, 注入真空收集瓶中, 使用气相色谱仪(Agilent 4890) 测定N2O浓度. N2O排放通量计算公式如下所示[11]:
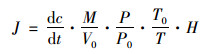
|
式中, dc/dt是N2O气体浓度随时间变化曲线斜率; M是气体摩尔质量; P是采样处大气压; T是采样时温度; V0、P0、T0分别是标准状况下气体摩尔体积, 大气压强和空气温度; H是水面上实验柱高度.
1.4.6 微生物群落多样性研究通过高通量测序对4个处理组底泥中微生物群落多样性进行对比分析, 包括DNA提取、PCR扩增及纯化、建库及MiSeq测序、数据分析这4个主要步骤.
DNA提取:分别称取4个处理组底泥样品约500 mg, 使用DNA快速提取试剂盒(MP Biomedicals, LLC)提取各底泥样品的DNA, 随后使用仪器NanoDrop® ND-1000(NanoDrop Technologies, Willmington, DE, USA)对提取的DNA样品进行浓度和纯度检测, 保证DNA样品浓度高于20 ng·μL-1, A260/A280介于1.80~2.00之间.
PCR扩增及纯化:利用16S rRNA基因序列的V1-V3区域对样品进行PCR扩增, 正向引物为515F(5′-GTGCCAGCMGCCGCGG-3′), 反向引物为907R(5′-CCGTCAATTCMTTTRAGTTT-3′).使用仪器ABI GeneAmp® 9700(ABI, USA)进行PCR扩增, 单个样品做4个PCR平行.扩增条件为:95℃ 2 min; 25循环(95℃ 30 s, 55℃ 30 s, 72℃ 30 s); 72℃ 5 min.随后将各样品的4个平行PCR体系混合, 利用PCR纯化试剂盒(Axygen, USA)对PCR产物进行纯化, 最终洗脱体积为30 μL.
建库及MiSeq测序:准确移取100 ng PCR纯化产物混合均匀, 进行建库.建库完成后使用仪器2100 Bioanalyzer(Agilent Technologies, Santa Clara, USA)进行芯片电泳用以确认建库成功.随后使用Qubit 2.0 Fluorometer对文库进行精确定量.将建库成功的文库按照比例混合、变性并稀释至终浓度为8 pmol·L-1, 随后进行MiSeq上机测序.
数据分析:通过Sickle软件(https://github.com/najoshi/sickle)去除低质量的序列; 用Mothur软件(http://www.mothur.org/wiki/Main_Page)对测序数据按照样品筛分, 然后按照Mothur软件提供的标准流程进行降噪处理, 对降噪数据进行比对分析, 最终获得底泥样品中微生物群落结构信息.
1.4.7 数据分析方法使用SPSS 13.0数据处理包对数据进行分析; 采用单向或双向方差分析(ANOVA)来区别不同处理组样品之间的差异性; 采用皮尔逊相关分析(Pearson correlation)评价数据变化的相关性; 采用Origin 8.6进行图形绘制.不同处理组之间的显著性差异水平设置为P < 0.05.
2 结果与讨论 2.1 水体15N测定结果分析4个处理组存积于水体中的同位素15N量及上覆水总氮浓度如图 2所示, 不同处理组间水体15N存积量存在明显差异(单向方差分析, P < 0.05).处理组A(裸泥组)水体中15N的存积最高, 约为380.02 μg, 处理组B(INCB组)和处理组C(伊乐藻组)相对A组较低, 分别为328.12 μg和255.79 μg, 而处理组D(INCB+伊乐藻组)水体中15N存积量最低, 约为211.28 μg.同时, 上覆水总氮浓度与水体15N存积量有相同的趋势(r=0.991, P < 0.05), 处理组A上覆水总氮浓度最高, 处理组D的总氮浓度最低.本实验结果说明, 相较于未经生态修复的处理组, 固定化高效脱氮微生物与沉水植物伊乐藻的协同作用强化了系统中氮素脱除能力.

|
图 2 不同处理组上覆水中同位素15N存积量(n=5) 及总氮浓度(n=3) Fig. 2 Isotopic 15N storage and TN concentration of overlying water in the different treatment groups |
同位素15N-NH4+添加后, 种植伊乐藻的处理组C和组D中, 水生植物15N丰度均有增加.如图 3所示, 伊乐藻对15N-NH4+的吸收量分别为127.22 μg和98.93 μg, 占同位素15N-NH4+总添加量的25.44%和19.79%.处理组C中可检测出的15N丰度明显高于D组(单向方差分析, P < 0.05), 这可能是由于处理组D添加INCB后, 微生物丰度提高, 导致脱氮微生物与伊乐藻相互竞争氮素营养盐[12], 部分含15N氮素被反硝化及厌氧氨氧化微生物利用, 以气体形式释放[13].水生植物中大量存积15N, 说明铵态氮可以被植物大量吸收, 植物对氮素的吸收是水生生态系统脱氮的主要途径之一.

|
图 3 不同处理组中伊乐藻对15N的吸收量(n=5) Fig. 3 Isotopic 15N taken up by Elodea nuttallii in the different treatment groups |
不同处理组中底泥对15N的存储量如图 4所示, 底泥中15N-NH4+的量介于24.3 μg(处理组D)到39.68 μg(处理组A).底泥对铵态氮的存储是氮素循环的重要过程之一, 存储强度受悬浮颗粒界面及特性的影响[14].处理组A中底泥对15N储存量略高于其他三组, 主要由于INCB的添加或伊乐藻种植可能增加了底泥中脱氮微生物的丰度及活性, 微生物的繁殖及伊乐藻生长会消耗底泥中的氮素营养盐[15].

|
图 4 不同处理组底泥中15N存储量(n=5) Fig. 4 Isotopic15N storage in sediment in the different treatment groups |
15NH4+添加后经过24 h的密闭培养, 不同处理组N2排放通量如图 5所示, 相较于其他3个处理组, 处理组B具有最高的氮气排放通量为164.32 μmol·(m2·h)-1, 最低的氮气排放通量出现在处理组C, 排放通量为49.65 μmol·(m2·h)-1(处理组B高于处理组D, 处理组C低于处理组A; 单向方差分析, P < 0.05).添加INCB的两个处理组(处理组B和处理组D)中的15N标记的N2排放通量高于处理组A和C, 这一结果说明INCB的添加促进了氮素向N2的转化.

|
图 5 不同处理组中N2排放通量(n=5) Fig. 5 N2 emission fluxes in the different treatment groups |
反硝化和厌氧氨氧化是两个主要微生物作用产生N2的过程[16], 图 6显示了4个不同处理组泥水界面的微生物组成种类, 从门级别分类结果来看, 在处理组A中主要是变形杆菌门微生物(Proteobacteria)和蓝细菌门微生物(Cyanobacteria), 占微生物群落组成的85.5%和11.73%;变形杆菌门微生物(Proteobacteria)、拟杆菌门微生物(Bacteroidetes)和酸杆菌门微生物(Acidobacteria)在处理组B中分别占65.61%、11.14%和4.76%;而在处理组C中, 变形杆菌门微生物(Proteobacteria)和硝化螺菌门(Nitrospira)的丰度较高, 占73.54%和8.64%;处理组D中微生物组成结构与其他三组不同, 变形杆菌门微生物(Proteobacteria)占46.65%, 其次是拟杆菌门微生物(Bacteroidetes)占18.72%, 酸杆菌门微生物(Acidobacteria)7.03%和硝化螺菌门(Nitrospira)5.88%. 4个不同处理组中变形门微生物所占比例显著高于其他门级别微生物(P < 0.05), 在变形门微生物中较常见的细菌有氨化细菌、硝化细菌和反硝化细菌, 最常见反硝化细菌都属于变形菌门[17].同时, 对于添加了INCB的处理组(处理组B和处理组D), 浮霉状菌门微生物(Planctomycetes)的丰度明显提升(P < 0.05), 在4个处理组占比分别为0.21%、2.68%、1.96%和3.82%.现有研究表明厌氧氨氧化细菌属于浮霉状菌, 厌氧氨氧化细菌广泛存在于自然和人工生态系统, 是生态系统氮素循环过程的重要环节[18, 19].根据N2产生量及微生物多样性测定结果, 拟杆菌门微生物(Bacteroidetes)、酸杆菌门微生物(Acidobacteria)和硝化螺菌门(Nitrospira)等在氮循环中具有重要作用的微生物丰度同样得到提升.

|
图 6 不同处理组的微生物群落门级别分类相对丰度(n=1) Fig. 6 Relative abundance of the phyla identified among the four different treatment groups |
同时, 通过对泥水界面底泥溶解氧侵蚀深度的研究, 4个不同处理组之间存在显著性差异(P < 0.05).如图 7所示, 种植伊乐藻的C、D两组溶解氧侵蚀深度约30 mm, 而A、B两组未种植伊乐藻的处理组中溶解氧侵蚀深度仅约14 mm.对比裸泥组及添加INCB的处理组之间, 其氧侵蚀深度并无明显区别, 说明INCB的添加对氧侵蚀深度无显著影响.在种植伊乐藻的处理组C和D中, 由于植物根部释放氧气, 氧侵蚀深度得到增加, 在根系周围底泥形成好氧-厌氧微环境的同时, 还会导致根区土壤氧化还原电位的变化[20, 21].因此, 添加INCB或沉水植物的处理组中反硝化细菌和厌氧氨氧化细菌丰度均有所上升, 微生物群落结构发生改变, 提升了氮素转化效率, 氮气产生速率得到提高.

|
图 7 不同处理组底泥溶解氧侵蚀深度(n=1) Fig. 7 Dissolved oxygen profiles in the different treatment sediments |
不同处理组中N2O排放通量如图 8所示, N2O排放通量介于217.90 μg·(m2·h)-1和55.06 μg·(m2·h)-1之间.其中处理组A的排放通量最低, 处理组D的排放通量最高, 添加INCB的处理组(处理组B和处理组D)相较于未添加INCB的处理组(处理组A和处理组C)具有更高的N2O排放通量(P < 0.05).底泥微生物参与的硝化和反硝化过程是产生N2O的主要途径[22], N2O是硝化和反硝化过程的中间产物之一[23], 根据图 6所示结果, 在种植伊乐藻的处理组C和D, 硝化螺菌门(Nitrospira)微生物丰度得到明显提升, 更高的微生物丰度和底泥溶解氧浓度会导致更高N2O释放量[24]. N2O排放通量及微生物丰度测定结果说明, 固定化高效脱氮微生物与伊乐藻联用技术的应用改善了底泥中微生物生存环境, 提升微生物丰度及活性, 促进氮素循环. N2O是一种温室气体, 对于添加了INCB和伊乐藻的处理组D, N2O排放通量较高. 表 2显示在相关研究中N2O排放通量, 其仅占总氮素气体比例极少部分.本研究结果发现, 水生植物的种植和微生物的活性会对N2O的释放产生影响.

|
图 8 不同处理组中N2O排放通量(n=5) Fig. 8 N2O emission fluxes in the different treatment groups |
|
|
表 2 相关研究中N2O排放通量及占总氮素气体比例 Table 2 Reported values of N2O emission and contribution to total nitrogen released in form of gas |
2.5 15N平衡模型
4个不同处理组内氮素平衡模型如图 9所示, 在处理组C和D组中沉水植物伊乐藻对15NH4+吸收率分别为25.44%和19.79%.在4个不同处理组中, 底泥对于15NH4+存储率分别为7.94%、5.52%、6.47%和4.86%. 15NH4+的总去除率分别为24%、34.38%、48.84%和57.74%, 在加入伊乐藻和INCB的D组中, 虽然INCB降低了植物对氮营养盐的吸收速率, 但总体氮素去除率仍大于添加INCB的B组和添加伊乐藻的C组.实验结果表明, 在各生态处理系统中, 植物吸收和微生物脱氮作用都能去除水体中的氮素, 均是氮素脱除有效途径.伊乐藻与高效脱氮微生物联用技术(E-INCB)的应用, 可以提高水体氮素的脱除速率, 对于促进水体净化具有重要作用.

|
图 9 不同处理组不同途径对于15NH4+的转化情况 Fig. 9 Proportion of 15NH4+ transformed by different pathways in the different treatment groups |
(1) 伊乐藻与固定化高效脱氮微生物联用修复技术的应用, 改变了泥水界面微环境, 增加了氧侵蚀深度, 为脱氮微生物提供良好的好氧-缺氧生存环境, 促进微生物对氮素吸收转化过程.
(2) 通过种植沉水植物伊乐藻和添加固定化高效脱氮微生物, 增加了底泥微生物的生物活性和丰度, 且在添加伊乐藻与脱氮微生物处理组中氮素脱除效率最高.
(3) 在添加伊乐藻和固定化高效脱氮微生物的处理组中, 变形杆菌门微生物(Proteobacteria)、拟杆菌门微生物(Bacteroidetes)、硝化螺菌门微生物(Nitrospira)和浮霉状菌门微生物(Planctomycetes)等微生物丰度均得到提高, 说明伊乐藻和固定化高效脱氮微生物的加入, 增强了河道底泥中的脱氮微生物的多样性.
(4) 根据室内模拟实验结果, 植物吸收和微生物生化作用去除水体中的氮素是氮素脱除的有效途径, 通过对伊乐藻与高效脱氮微生物联用技术的应用, 可以提高水体氮素的脱除速率, 对于促进水体净化具有重要作用.
| [1] | Yang X Y, Liu Q, Fu G T, et al. Spatiotemporal patterns and source attribution of nitrogen load in a river basin with complex pollution sources[J]. Water Research, 2016, 94: 187-199. DOI:10.1016/j.watres.2016.02.040 |
| [2] | Wu H M, Zhang J, Li C, et al. Mass balance study on phosphorus removal in constructed wetland microcosms treating Polluted River water[J]. CLEAN-Soil, Air, Water, 2013, 41(9): 844-850. DOI:10.1002/clen.v41.9 |
| [3] |
吴锋, 战金艳, 邓祥征, 等. 中国湖泊富营养化影响因素研究——基于中国22个湖泊实证分析[J]. 生态环境学报, 2012, 21(1): 94-100. Wu F, Zhan J Y, Deng X Z, et al. Influencing factors of lake eutrophication in China——a case study in 22 lakes in China[J]. Ecology and Environmental Sciences, 2012, 21(1): 94-100. |
| [4] |
李宝珍, 范晓荣, 徐国华. 植物吸收利用铵态氮和硝态氮的分子调控[J]. 植物生理学通讯, 2009, 45(1): 80-88. Li B Z, Fan X R, Xu G H. Molecular regulation for uptake and utilization of ammonium and nitrate in plant[J]. Plant Physiology and Molecular Biology, 2009, 45(1): 80-88. |
| [5] |
刘丹丹, 李正魁, 叶忠香, 等. 伊乐藻和氮循环菌技术对太湖氮素吸收和反硝化的影响[J]. 环境科学, 2014, 35(10): 3764-3768. Liu D D, Li Z K, Ye Z X, et al. Nitrogen uptake and denitrification study on the joint treatment of aquatic vegetation and immobilized nitrogen cycling bacteria in Taihu Lake[J]. Environmental Science, 2014, 35(10): 3764-3768. |
| [6] |
韩华杨, 李正魁, 王浩, 等. 伊乐藻-固定化脱氮微生物技术对入贡湖河道脱氮机制的影响[J]. 环境科学, 2016, 37(4): 1397-1403. Han H Y, Li Z K, Wang H, et al. Effect of Elodea nuttallii-immobilized nitrogen cycling bacteria on nitrogen removal mechanism in an Inflow river, Gonghu Bay[J]. Environmental Science, 2016, 37(4): 1397-1403. |
| [7] | Peipoch M, Gacia E, Blesa A, et al. Contrasts among macrophyte riparian species in their use of stream water nitrate and ammonium:insights from 15N natural abundance[J]. Aquatic Sciences, 2014, 76(2): 203-215. DOI:10.1007/s00027-013-0330-7 |
| [8] |
赵琳, 李正魁, 周涛, 等. 伊乐藻-氮循环菌联用对太湖梅梁湾水体脱氮的研究[J]. 环境科学, 2013, 34(8): 3057-3063. Zhao L, Li Z K, Zhou T, et al. Denitrification study of Elodea nuttallii-nitrogen cycling bacteria restoration in Meiliang Bay, Taihu Lake[J]. Environmental Science, 2013, 34(8): 3057-3063. |
| [9] | Meng P P, Hu W R, Pei H Y, et al. Effect of different plant species on nutrient removal and rhizospheric microorganisms distribution in horizontal-flow constructed wetlands[J]. Environmental Technology, 2014, 35(5-8): 808-816. |
| [10] | Li Z K, Pu P M, Hu W P, et al. Improvement of Taihu water quality by the technology of immobilized nitrogen cycle bacteria[J]. Nuclear Science and Techniques, 2002, 13(2): 115-118. |
| [11] | Song C C, Wang Y S, Wang Y Y, et al. Emission of CO2, CH4 and N2O from freshwater marsh during freeze-thaw period in northeast of China[J]. Atmospheric Environment, 2006, 40(35): 6879-6885. DOI:10.1016/j.atmosenv.2005.08.028 |
| [12] | Toet S, Huibers L H F A, Van Logtestijn R S P, et al. Denitrification in the periphyton associated with plant shoots and in the sediment of a wetland system supplied with sewage treatment plant effluent[J]. Hydrobiologia, 2003, 501(1-3): 29-44. |
| [13] | Li C, Ren H Q, Xu M, et al. Study on anaerobic ammonium oxidation process coupled with denitrification microbial fuel cells (MFCs) and its microbial community analysis[J]. Bioresource Technology, 2015, 175: 545-552. DOI:10.1016/j.biortech.2014.10.156 |
| [14] | Bédard C, Knowles R. Physiology, biochemistry, and specific inhibitors of CH4, NH4+, and CO oxidation by Methanotrophs and Nitrifiers[J]. Microbiological Reviews, 1989, 53(1): 68-84. |
| [15] | Eriksson P G, Weisner S E B. An experimental study on effects of submersed macrophytes on nitrification and denitrification in ammonium-rich aquatic systems[J]. Limnology and Oceanography, 1999, 44(8): 1993-1999. DOI:10.4319/lo.1999.44.8.1993 |
| [16] | Risgaard-Petersen N, Nielsen L P, Rysgaard S, et al. Application of the isotope pairing technique in sediments where anammox and denitrification coexist[J]. Limnology and Oceanography-Methods, 2003, 1(1): 63-73. DOI:10.4319/lom.2003.1.63 |
| [17] |
杨浩, 张国珍, 杨晓妮, 等. 16S rRNA高通量测序研究集雨窖水中微生物群落结构及多样性[J]. 环境科学, 2017, 38(4): 1704-1716. Yang H, Zhang G Z, Yang X N, et al. Microbial community structure and diversity in cellar water by 16S rRNA high-throughput sequencing[J]. Environmental Science, 2017, 38(4): 1704-1716. |
| [18] | Schmid M C, Maas B, Dapena A, et al. Biomarkers for in situ detection of anaerobic ammonium-oxidizing (anammox) bacteria[J]. Applied and Environmental Microbiology, 2005, 71(4): 1677-1684. DOI:10.1128/AEM.71.4.1677-1684.2005 |
| [19] | Schmid M C, Risgaard-Petersen N, Van De Vossenberg J, et al. Anaerobic ammonium-oxidizing bacteria in marine environments:widespread occurrence but low diversity[J]. Environmental Microbiology, 2007, 9(6): 1476-1484. DOI:10.1111/emi.2007.9.issue-6 |
| [20] | Soana E, Bartoli M. Seasonal variation of radial oxygen loss in Vallisneria spiralis L.:an adaptive response to sediment redox?[J]. Aquatic Botany, 2013, 104: 228-232. DOI:10.1016/j.aquabot.2012.07.007 |
| [21] | Christensen P B, Revsbech N P, Sand-Jensen K. Microsensor analysis of oxygen in the rhizosphere of the aquatic macrophyte Littorella uniflora (L.) ascherson[J]. Plant Physiology, 1994, 105(3): 847-852. DOI:10.1104/pp.105.3.847 |
| [22] | Bouwman A F. Nitrogen oxides and tropical agriculture[J]. Nature, 1998, 392(6679): 866-867. DOI:10.1038/31809 |
| [23] | Kimochi Y, Inamori Y, Mizuochi M, et al. Nitrogen removal and N2O emission in a full-scale domestic wastewater treatment plant with intermittent aeration[J]. Journal of Fermentation and Bioengineering, 1998, 86(2): 202-206. DOI:10.1016/S0922-338X(98)80114-1 |
| [24] | Sun H Y, Zhang C B, Song C B, et al. The effects of plant diversity on nitrous oxide emissions in hydroponic microcosms[J]. Atmospheric Environment, 2013, 77: 544-547. DOI:10.1016/j.atmosenv.2013.05.058 |
| [25] | Davidsson T E, Stepanauskas R, Leonardson L. Vertical patterns of nitrogen transformations during infiltration in two wetland soils[J]. Applied and Environmental Microbiology, 1997, 63(9): 3648-3656. |
| [26] | Søvik A K, Augustin J, Heikkinen K, et al. Emission of the greenhouse gases nitrous oxide and methane from constructed wetlands in Europe[J]. Journal of Environmental Quality, 2006, 35(6): 2360-2373. DOI:10.2134/jeq2006.0038 |
| [27] | Maltais-Landry G, Maranger R, Brisson J. Effect of artificial aeration and macrophyte species on nitrogen cycling and gas flux in constructed wetlands[J]. Ecological Engineering, 2009, 35(2): 221-229. DOI:10.1016/j.ecoleng.2008.03.003 |
| [28] | Elgood Z, Robertson W D, Schiff S L, et al. Nitrate removal and greenhouse gas production in a stream-bed denitrifying bioreactor[J]. Ecological Engineering, 2010, 36(11): 1575-1580. DOI:10.1016/j.ecoleng.2010.03.011 |