 2017, Vol. 38
2017, Vol. 38



随着能源危机与环境污染的加剧, 微藻生物柴油越来越受到各国研究者的关注[1~3].就整体工艺而言, 微藻生物柴油的生产包括藻种选育、微藻培养、微藻收获、油脂提取与转脂.微藻培养是其中一个重要环节, 在微藻培养过程中, 利用淡水培养微藻需消耗大量淡水资源, 有悖于当前淡水资源紧缺的背景.同时, 营养物质的投加进一步提高了微藻的培养成本[4, 5].利用城市污水培养微藻, 藻类通过光合作用释放出氧气供给好氧异养型微生物进行代谢活动, 而好氧异养型微生物对有机污染物进行氧化分解, 代谢产物二氧化碳和无机氮、磷化合物又供给藻类作为光合作用所需的碳源和营养, 如此循环, 形成藻菌之间共生的关系[6].菌藻混合培养既可以收获藻类, 又可以净化污水, 受到国内外极大关注[7].
陈小晨等[8]发现低浓度的光合细菌液对铜绿微囊藻和小球藻具有明显的促进作用, 且小球藻受到的影响比铜绿微囊藻要显著. Zhou等[9]以曲霉为生物质载体实现小球藻的固定化培养, 利用菌藻共生体高效收获藻体.细菌除了对微藻的生长有影响, 在污水处理方面也有广泛的应用. Hargreaves[10]将光合细菌应用于水产养殖, 增加了水体的溶氧量, 有效降低水体污染程度.王冰等[11]研究了小球藻和光合细菌共培养对污水的处理效果, 结果表明菌藻共生体对氨氮的去除率在4 h内可达到80%.目前, 此领域的大多数研究集中在利用微藻改善污水水质或者利用人工污水培养基及灭菌污水培养微藻方面, 而将实际城市污水处理与微藻培养制备生物柴油相耦合的研究报道较少, 也极少关注微藻培养后菌群结构的变化情况[12, 13].
鉴于城市污水含有大量的原生菌类, 且微藻与细菌之间存在着互生、拮抗等复杂的相互关系[14].本研究将基于城市污水系统中菌藻共生体系, 筛选出能够有效促进小球藻生长和油脂积累的细菌, 通过分析体系中菌群多样性并确定主导菌群, 初步探讨菌类促进藻类生长的机制, 使菌藻共生体系更好地指导微藻培养与污水处理耦合技术的进一步推广应用.
1 材料与方法 1.1 藻种与培养基实验所用微藻为课题组前期经离子注入法诱变筛选的蛋白核小球藻诱变株(CH-M)[15], 藻种采用BG-11培养基[16]进行传代培养保藏.选取的细菌共有3个来源:水产养殖领域常用的对水质具有净化作用的光合细菌(广东省菌种保藏中心)、EM菌和枯草芽孢杆菌(河南中广集团天义生物谷); 课题组前期从污水中筛选的高效氨化菌、聚磷菌、阴离子洗涤剂降解菌(LAS菌)、二甲基甲酰胺降解菌(DMF菌)[17]; 从小球藻污水培养系统中筛选出的可在培养基中快速生长的6种细菌W1、W2、W3、W4、W5、W6[18].
实验所用城市污水取自深圳大学城市政污水井, 经简单预处理后的污水水质见表 1.
|
|
表 1 大学城城市污水水质/mg·L-1 Table 1 Municipal wastewater quality at the Shenzhen University/mg·L-1 |
1.2 实验方法 1.2.1 微藻培养
以BG11培养基培养处于对数生长期的微藻为种子液, 接种培养体积为600 mL, 于气泡柱式光生物反应器(直径为5 cm、长度为50 cm、容积大小约为1 L)中培养, 以相应的不加微藻的城市污水为空白对照, 使扣除空白后藻液初始吸光度D680=0.1, 于培养温度(25±1)℃, 光照强度100 μmol·(m2·s)-1, 光暗比(L:D)为12 h:12 h条件下, 连续通气培养(空气流速为120 mL·min-1, 由底部通入).每天取样测定扣除空白后藻液的D680以监测藻细胞生长情况.取培养达到稳定期的藻液测定扣除空白后藻细胞的干重、油脂产量.每组实验各设3个平行.
1.2.2 污水预处理方法城市污水采用沉淀+过滤的预处理方式, 原污水先沉淀2 h处理后, 上清液用定性滤纸过滤后使用.
1.3 分析方法 1.3.1 微藻干重的测定微藻的干重采用重量法测定, 10 mL藻液经预称重(W1)的微孔滤膜(Φ0.45 μm)过滤、洗涤, 于105℃下烘至恒重(W2), 不加微藻的空白对照组干重为W0, 微藻干重DW (g·L-1)的计算公式如下:
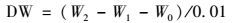
|
微藻油脂提取采用氯仿甲醇共溶剂提取法[19].待提取结束后, 收集氯仿相, 转移至预称重的锡纸盘中, 待有机溶剂挥发完全后, 于80℃烘箱中烘至恒重, 得到微藻油脂产量, 微藻细胞油脂含量为微藻油脂产量与微藻干重的质量比.
微藻脂肪酸分析, 首先对提取的油脂进行甲酯化[20], 将5 mL藻液中脂肪酸转化成相对应的脂肪酸甲酯.采用赛里安456-GC气相色谱仪进行分析, 色谱柱为BR-2560柱, 100 m×0.25 mm(内径)×0.20 μm(膜厚), FID检测器, 分流方式进样, 分流比为30:1, 进样量为1 μL.
1.3.3 菌群结构分析方法微生物菌群结构的研究主要是通过对细菌16S rRNA的V4~V5高可变区基因进行测序, 并基于测序结果进行数据筛选以及微生物菌群信息的分析.主要包括:细菌组DNA提取、PCR扩增、电泳验证、PCR产物纯化与浓度测定、高通量测序以及测序结果分析.细菌组DNA的提取采用MOBIO PowerSoil® DNA Isolation Kit试剂盒(MOBIO Solon, USA), 将提取出来的DNA采用引物对515F:5′-GTGCCAGCMGCCGCGGTAA-3′和907R:5′-CCGTCAATTCMTTTRAGTTT-3′对16S rRNA高可变的V4~V5区进行序列扩增. PCR产物纯度采用1%的琼脂糖凝胶电泳进行验证, DNA marker采用DL2000(SIGMA, USA).电泳条件为100 V电压, 电泳30 min.最后通过紫外凝胶成像系统观察目的DNA片段大小, 估测DNA浓度, PCR产物纯化采用E.Z.N.A.® Gel Extraction Kit(OMEGA, USA).最后将胶回收后的PCR产物建库后使用Hiseq2500 PE250(Illumina, USA)测序平台对构建的扩增子文库进行测序[21].
1.3.4 数据分析方法数据采用统计软件SPSS 21.0进行单因素方差分析(one-way ANOVA), 采用LSD法进行统计检验(P<0.05).
2 结果与讨论 2.1 细菌促进小球藻产脂的初步优化通过考察13种菌对小球藻的干重和油脂产量的促进作用, 初步筛选出促进作用明显的细菌.将小球藻按照初始吸光度D680=0.1接种到投加了不同种类细菌的城市污水中, 污水中细菌初始吸光度D600为0.02, 取稳定期的藻液测其干重, 结果如图 1所示.从中可以看出, 所投加的细菌中, 光合细菌、枯草芽孢杆菌、氨化菌、DMF菌、W4、W5、W6等7种菌对小球藻干重促进作用明显, 其中投加W4菌的情况下微藻干重增加最为显著.不同细菌投加情况下, 微藻的油脂含量和油脂产量如图 2、图 3所示, 不同字母表示组间差异显著(P<0.05).

|
图 1 小球藻在添加不同细菌城市污水中的生物质产量 Fig. 1 Biomass production of microalgea in municipal wastewater with different bacteria |

|
图 2 小球藻在添加不同细菌城市污水中的油脂含量 Fig. 2 Lipid content of microalgea in municipal wastewater with different bacteria |

|
图 3 小球藻在添加不同细菌城市污水中的油脂产量 Fig. 3 Lipid production of microalgea in municipal wastewater with different bacteria |
由图 2、图 3可以看出, 投加细菌后大多数细菌对微藻油脂含量影响不大, 小球藻油脂含量受不同细菌种类的影响不大.由于微藻的油脂产量由微藻干重和油脂含量共同决定, 投加光合细菌、枯草芽孢杆菌、DMF菌、W4和W5对小球藻的油脂产量提高有明显促进作用(P<0.05), 其中光合细菌对小球藻油脂产量的促进作用最为显著, 油脂产量相比于空白对照组提高了46.51%.因此, 选取对微藻生长和产脂具有显著促进作用的光合细菌、枯草芽孢杆菌、DMF菌、W4和W5作为后续进一步优化的对象.
2.2 细菌促进小球藻产脂的进一步优化 2.2.1 最佳菌种及浓度的确定选取对小球藻油脂产量提高促进作用明显的5种菌:光合细菌、枯草芽孢杆菌、DMF菌、W4和W5, 对其投加量进行进一步优化, 5种细菌在污水中的初始吸光度D600均设置为0、0.002、0.01、0.02、0.03, 将小球藻按照初始吸光度D680=0.1接种到添加不同量细菌的城市污水中.取稳定期的藻液测其微藻干重, 油脂产量, 并计算其油脂含量, 所得结果如图 4~6所示.从中可以看出, 光合细菌的初始吸光度D600为0.01, W4菌的初始吸光度D600为0.02时, 对小球藻的干重和油脂产量促进作用最显著, 干重比空白对照组分别显著提高了32.09%和33.69%(P<0.05), 油脂产量比空白对照组分别显著提高了22.58%、21.50%(P<0.05).因此选取初始吸光度D600为0.01的光合细菌和初始吸光度D600为0.02的W4菌作为后续研究对象.

|
图 4 小球藻在添加不同量细菌城市污水中的生物质产量 Fig. 4 Biomass production of microalgea in municipal wastewater with different amount of bacteria |

|
图 5 小球藻在添加不同量细菌城市污水中的油脂含量 Fig. 5 Lipid content of microalgea in municipal wastewater with different amount of bacteria |

|
图 6 小球藻在添加不同量细菌城市污水中的油脂产量 Fig. 6 Lipid production of microalgea in municipal wastewater with different amount of bacteria |
对投加光合细菌, W4菌与空白对照组油脂进行甲酯化反应, 通过对生成的脂肪酸甲酯进行气相色谱分析, 考察小球藻脂肪酸组成的差异, 结果如表 2所示.从中可知, 4种微藻在脂肪酸组成上均以C16、C18为主, 为绿藻中常见脂肪酸[22, 23]. Song等[24]和Ma等[25]的研究表明, 适合制备生物柴油的原料应含有较多的C16~C18脂肪酸, 主要包括十六烷酸(C16:0)、十八烷酸(C18:0)、十八碳烯酸(C18:1)、十八碳二烯酸(C18:2)、十八碳三烯酸(C18:3).对照组与投加光合细菌、W4菌的实验组中该5种脂肪酸在总脂肪酸中的质量占比分别可达94.41%、93.91%、94.14%, 适宜于作为制备生物柴油的原料.同时, 脂肪酸甲酯的结构会对生物柴油的性能(十六烷值、低温流动性、氧化安定性等)产生影响[26].有研究表明, 生物柴油的十六烷值随着饱和脂肪酸甲酯含量的增加而增加, 多不饱和脂肪酸甲酯含量的增加而降低[27].生物柴油低温流动性随着饱和脂肪酸甲酯尤其是长碳链质量分数增高而变差.生物柴油的氧化安定性随着不饱和脂肪酸不饱和度增高而变差.高品位的生物柴油应尽量少地含有多不饱和脂肪酸甲酯和饱和脂肪酸甲酯, 尽量多地含有单不饱和脂肪酸, 以改善其十六烷值、低温流动性、氧化安定性、运动黏度等性能[28].从表 2中可以看出, 投加光合细菌、W4菌的实验组中单不饱和脂肪酸的质量占比分别为10.39%、10.60%, 比对照组5.35%有大幅度提高, 有利于提升所得生物柴油的品位.
|
|
表 2 添加细菌组与对照组脂肪酸分析/% Table 2 Fatty acid profiles of microalgae with bacteria/% |
2.3 细菌对菌群结构影响
取培养达到稳定期的空白对照组与投加细菌(光合细菌, W4菌)的实验组藻液, 通过DNA提取、16S V4~V5高变区PCR扩增, 高通量测序考察藻液中菌群的组成与多样性.
2.3.1 菌群多样性分析菌群多样性是指某个集合群落内, 不同类型微生物的变化以及它们之间的相对丰度.一般而言, 系统中微生物菌群多样性越高, 越有利于抵抗外界环境的变化, 较快的进行系统自我调整和更新.反之, 微生物菌群多样性较低, 存在优势度较高微生物群落的系统, 可能富集重要的功能种群, 有利于对目标污染物的去除, 但可能带来运行不稳定的风险.
生物多样性测定主要有3个空间尺度从小到大分为:α多样性、β多样性、γ多样性.其中α多样性主要关注局域均匀生境下的物种数目, 因此也被称为生境内的多样性.多样性指数是反映丰富度和均匀度的综合指标, 与以下两个因素有关:① 种类数目, 即丰富度; ② 种类中个体分配上的均匀性.群落丰富度(community richness)的指数, 包括Chao1指数和observed species指数, 数值越大, 说明样品中物种越丰富; 群落多样性的指数, 包括PD whole tree指数、Shannon指数和Simpson指数, 数值越大, 说明样品群落多样性越高.所得实验组与对照组样品的α多样性参数统计结果如表 3所示.从中可以看出, 投加细菌(光合细菌, W4菌)的实验组Chao1指数和observed species指数均小于对照组, 表明投加细菌会降低藻液中菌群的丰富度, 虽然PD whole tree指数有所降低, 但Shannon指数和Simpson指数并未发生明显变化, 表明投加细菌并未改变菌群多样性.
|
|
表 3 不同样品的生物多样性 Table 3 Microbial diversity of different samples |
2.3.2 主导菌群分析
对投加光合细菌、W4菌的实验组与空白对照组有效序列在不同物种分类水平上的丰度进行分析. 图 7为样品在门分类级别上的丰度.从中可以看出, 对照组与实验组在门级别上, 均以变形菌门(Proteobacteria)占主导.这与世界各地分布的污水处理厂中微生物菌群结构的相关报道一致[29, 30].空白对照组与投加光合细菌、W4菌组, Proteobacteria丰度分别高达84.13%、84.50%和77.99%.其他占主导的门为拟杆菌门(Bacteroidetes)丰度分别为11.33%、13.89%和20.16%.由于Proteobacteria门的丰度高达77.85%~84.50%, 因而分析了其下属的α-、β-、γ-、δ-、ε-Proteobacteria的丰度.结果表明, 空白对照组α-Proteobacteria和γ-Proteobacteria的丰度较多, 分别为32.52%和31.16%, 其它依次为β-Proteobacteria, 丰度为20.24%.而投加细菌(光合细菌、W4) 组γ-Proteobacteria的丰度最大, 分别为36.91%和32.62%, 其次为α-Proteobacteria和β-Proteobacteria, 丰度分别为30.91%、24.40%和16.65%、20.81%.即相比于空白对照组, 投加细菌的实验组3种Proteobacteria变化不大.进一步对比空白对照组与实验组的属分类级别上的丰度.实验组丛毛单胞菌属(Comamonas)和假单胞菌属(Pseudomonas)的丰度大于对照组.丛毛单胞菌属可以将氨氮分解为亚硝酸盐氮, 同时可以将氨氮转化为氮气, 既能够作为脱氮微生物将氨氮转化为亚硝酸盐氮, 又能够将氨氮转化为氮气, 实现短程硝化-反硝化.该菌株的益处是具有对含氮废水的硝化-反硝化作用, 对氨氮的降解速度快, 适用于含氮工业废水、含氮生活污水的处理.假单胞菌属有利于废水中有机物残留尤其毒性芳香烃类化合物的降解, 说明投加的细菌有利于污水水质的改善.

|
图 7 样品在不同层级(门纲属)上的丰度 Fig. 7 Relative abundance at different levels |
城市污水中光合细菌初始吸光度D600为0.01, W4菌初始吸光度D600为0.02时, 对小球藻的干重和油脂产量促进作用最显著, 油脂产量分别可达0.114 g·L-1、0.113 g·L-1, 油脂产量比空白对照组分别提高了22.58%、21.50%.通过对生成的脂肪酸甲酯进行气相色谱分析, 结果显示光合细菌和W4菌的添加并未改变小球藻脂肪酸成分, 但提升了单不饱和脂肪酸的含量, 有利于提升所得生物柴油的品位.培养结束后污水的菌群结构分析显示投加细菌会降低污水中菌群的丰富度和多样性, 初步判断是投加的菌在藻液中能够成为优势菌群, 且实验组中丛毛单胞菌属和假单胞菌属的丰度大于对照组.
| [1] | Chisti Y. Constraints to commercialization of algal fuels[J]. Journal of Biotechnology, 2013, 167(3): 201-214. DOI:10.1016/j.jbiotec.2013.07.020 |
| [2] | Han S F, Jin W B, Tu R J, et al. Biofuel production from microalgae as feedstock: current status and potential[J]. Critical Reviews in Biotechnology, 2015, 35(2): 255-268. DOI:10.3109/07388551.2013.835301 |
| [3] | Abomohra A E F, Jin W B, Tu R J, et al. Microalgal biomass production as a sustainable feedstock for biodiesel: current status and perspectives[J]. Renewable and Sustainable Energy Reviews, 2016, 64: 596-606. DOI:10.1016/j.rser.2016.06.056 |
| [4] | Yang J, Xu M, Zhang M X, et al. Life-cycle analysis on biodiesel production from microalgae: water footprint and nutrients balance[J]. Bioresource Technology, 2011, 102(1): 159-165. DOI:10.1016/j.biortech.2010.07.017 |
| [5] | Morales-Amaral M D M, Gómez-Serrano C, Acién F G, et al. Production of microalgae using centrate from anaerobic digestion as the nutrient source[J]. Algal Research, 2015, 9: 297-305. DOI:10.1016/j.algal.2015.03.018 |
| [6] |
巫小丹, 阮榕生, 王辉, 等. 菌藻共生系统处理废水研究现状及发展前景[J]. 环境工程, 2014, 32(3): 35-37, 69. Wu X D, Ruan R S, Wang H, et al. Current status and prospect of sewage purification with the algal-microbe symbiotic system[J]. Environmental Engineering, 2014, 32(3): 35-37, 69. |
| [7] | McKinlay J B, Harwood C S. Carbon dioxide fixation as a central redox cofactor recycling mechanism in bacteria[J]. Proceedings of the National Academy of Sciences of the United States of America, 2010, 107(26): 11669-11675. DOI:10.1073/pnas.1006175107 |
| [8] |
陈小晨, 薛凌展, 林泽, 等. 光合细菌对铜绿微囊藻和小球藻生长的影响研究[J]. 安徽农学通报, 2010, 16(23): 29-31. Chen X C, Xue L Z, Lin Z, et al. The effect of photosynthetic bacteria(PSB) on the growth of microcystis aeruginosa and chlorella vulgaris[J]. Anhui Agricultural Science Bulletin, 2010, 16(23): 29-31. DOI:10.3969/j.issn.1007-7731.2010.23.013 |
| [9] | Zhou W G, Cheng Y L, Li Y, et al. Novel fungal pelletization-assisted technology for algae harvesting and wastewater treatment[J]. Applied Biochemistry and Biotechnology, 2012, 167(2): 214-228. DOI:10.1007/s12010-012-9667-y |
| [10] | Hargreaves J A. Photosynthetic suspended-growth systems in aquaculture[J]. Aquacultural Engineering, 2006, 34(3): 344-363. DOI:10.1016/j.aquaeng.2005.08.009 |
| [11] |
王冰, 周集体, 杨宝灵, 等. 光合细菌-藻类共固定深度净化污水的研究[J]. 大连民族学院学报, 2014, 16(3): 249-252. Wang B, Zhou J T, Yang B L, et al. Deep treatment of wastewater by co-immobilized photosynthetic bacteria and algae[J]. Journal of Dalian Nationalities University, 2014, 16(3): 249-252. |
| [12] | Ramsundar P, Guldhe A, Singh P, et al. Assessment of municipal wastewaters at various stages of treatment process as potential growth media for Chlorella sorokiniana under different modes of cultivation[J]. Bioresource Technology, 2017, 227: 82-92. DOI:10.1016/j.biortech.2016.12.037 |
| [13] | Ryu B G, Kim E J, Kim H S, et al. Simultaneous treatment of municipal wastewater and biodiesel production by cultivation of Chlorella vulgaris with indigenous wastewater bacteria[J]. Biotechnology and Bioprocess Engineering, 2014, 19(2): 201-210. DOI:10.1007/s12257-013-0250-3 |
| [14] | Muñoz R, Guieysse B. Algal—bacterial processes for the treatment of hazardous contaminants: a review[J]. Water Research, 2006, 40(15): 2799-2815. DOI:10.1016/j.watres.2006.06.011 |
| [15] | Tu R J, Jin W B, Wang M, et al. Improving of lipid productivity of the biodiesel promising green microalga Chlorella pyrenoidosa via low-energy ion implantation[J]. Journal of Applied Phycology, 2016, 28(4): 2159-2166. DOI:10.1007/s10811-015-0783-2 |
| [16] | Stanier R Y, Kunisawa R, Mandel M, et al. Purification and properties of unicellular blue-green algae (order Chroococcales)[J]. Bacteriological Reviews, 1971, 35(2): 171-205. |
| [17] |
闫韫. 布吉河生物修复过程中氮循环功能菌群分布研究[D]. 哈尔滨: 哈尔滨工业大学, 2008. Yan Y. Study on the distribution of nitrogen ceycling microbial community in bioremediation process of Buji River[D]. Harbin: Harbin Institute of Technology, 2008. http://cdmd.cnki.com.cn/Article/CDMD-10213-2009291866.htm |
| [18] | 郭仕达. 城市污水高效降解菌对产油微藻生长促进作用的研究[D]. 哈尔滨: 哈尔滨工业大学, 2017. |
| [19] | Folch J, Lees M, Sloane Stanley G H. A simple method for the isolation and purification of total lipides from animal tissues[J]. Journal of Biological Chemistry, 1957, 226(1): 497-509. |
| [20] | Kaczmarzyk D, Fulda M. Fatty acid activation in cyanobacteria mediated by acyl-acyl carrier protein synthetase enables fatty acid recycling[J]. Plant Physiology, 2010, 152(3): 1598-1610. DOI:10.1104/pp.109.148007 |
| [21] | Hua Z S, Han Y J, Chen L X, et al. Ecological roles of dominant and rare prokaryotes in acid mine drainage revealed by metagenomics and metatranscriptomics[J]. The ISME Journal, 2015, 9(6): 1280-1294. DOI:10.1038/ismej.2014.212 |
| [22] |
刘林林, 黄旭雄, 危立坤, 等. 15株微藻对猪场养殖污水中氮磷的净化及其细胞营养分析[J]. 环境科学学报, 2014, 34(8): 1986-1994. Liu L L, Huang X X, Wei L K, et al. Removal of nitrogen and phosphorus by 15 strains of microalgae and their nutritional values in piggery sewage[J]. Acta Scientiae Circumstantiae, 2014, 34(8): 1986-1994. |
| [23] | Vidyashankar S, VenuGopal K S, Swarnalatha G V, et al. Characterization of fatty acids and hydrocarbons of chlorophycean microalgae towards their use as biofuel source[J]. Biomass and Bioenergy, 2015, 77: 75-91. DOI:10.1016/j.biombioe.2015.03.001 |
| [24] | Song M M, Pei H Y, Hu W R, et al. Evaluation of the potential of 10 microalgal strains for biodiesel production[J]. Bioresource Technology, 2013, 141: 245-251. DOI:10.1016/j.biortech.2013.02.024 |
| [25] | Ma Y B, Wang Z Y, Yu C J, et al. Evaluation of the potential of 9 Nannochloropsis strains for biodiesel production[J]. Bioresource Technology, 2014, 167: 503-509. DOI:10.1016/j.biortech.2014.06.047 |
| [26] | Stansell G R, Gray V M, Sym S D. Microalgal fatty acid composition: implications for biodiesel quality[J]. Journal of Applied Phycology, 2012, 24(4): 791-801. DOI:10.1007/s10811-011-9696-x |
| [27] |
方利国, 林璟. 脂肪酸甲酯对生物柴油十六烷值影响的研究[J]. 化工新型材料, 2008, 36(11): 94-96. Fang L G, Lin J. Effects of fatty acid methylesters on the cetane number of biodiesel[J]. New Chemical Materials, 2008, 36(11): 94-96. DOI:10.3969/j.issn.1006-3536.2008.11.033 |
| [28] | Hoekman S K, Broch A, Robbins C, et al. Review of biodiesel composition, properties, and specifications[J]. Renewable and Sustainable Energy Reviews, 2012, 16(1): 143-169. DOI:10.1016/j.rser.2011.07.143 |
| [29] | Wang X H, Wen X H, Yan H J, et al. Bacterial community dynamics in a functionally stable pilot-scale wastewater treatment plant[J]. Bioresource Technology, 2011, 102(3): 2352-2357. DOI:10.1016/j.biortech.2010.10.095 |
| [30] | Zhang T, Shao M F, Ye L. 454 Pyrosequencing reveals bacterial diversity of activated sludge from 14 sewage treatment plants[J]. The ISME Journal, 2012, 6(6): 1137-1147. DOI:10.1038/ismej.2011.188 |